Abstrak
LISNAWITA. Survei dan Sebaran Geografi Nematoda Sista Kentang
Indonesia. Dibimbing oleh MEITY SURADJI SINAGA, GUSTAF ADOLF
WATTIMENA, SUPRAMANA dan GEDE SUASTIKA
Nematoda sista kentang (NSK) merupakan patogen utama yang menginfeksi tanaman kentang dan mempunyai daerah sebaran yang sangat luas di seluruh dunia. Di Indonesia, NSK merupakan patogen baru, yang dapat menjadi kendala utama pada produksi kentang. Melalui metode survei dilakukan penelitian untuk mengetahui sebaran geografi NSK di beberapa sentra pertanaman kentang yaitu Jawa Timur, Jawa Tengah dan Jawa Barat. Survei dilakukan pada 30 lokasi tanaman kentang masing-masing 6 lokasi di Jawa Timur (survei pada Desember 2004), 4 lokasi di Jawa Tengah (survei pada Oktober 2005) dan 20 lokasi di Jawa Barat (survei pada Pebruari 2006 dan 2007). Hasil survei menunjukkan bahwa NSK telah tersebar luas di beberapa wilayah di Jawa Timur, Jawa Tengah dan Jawa Barat dengan kepadatan populasi bervariasi dari rendah hingga tinggi. Kepadatan populasi NSK pada saat survei dilakukan untuk 3 lokasi di Desa Tulung Rejo (Kabupaten Kota Batu, Jawa Timur) berturut-turut adalah 21, 44 dan 675 sista per 100 ml tanah. Untuk Jawa Tengah, kepadatan populasi tertinggi terdapat pada Desa Pawuhan (Kabupaten Banjarnegara), yaitu 400 sista, diikuti dengan Desa Karangtengah (Kabupaten Banjarnegara), yaitu 270 sista, Desa Kepakisan (Kabupaten Banjarnegara) dengan 21 sista dan Desa Petak Bateng (Kabupaten Wonosobo) dengan 2 sista per 100 ml tanah. Sedangkan kepadatan populasi untuk Jawa Barat untuk 3 lokasi di Desa Sukamanah (Kabupaten B&ung) adalah 28 sista, 19 sista dan 2 sista per 100 ml tanah, dan satu lokasi Desa Mekarsari (Kabupaten B&ung) adalah 17 sista per 100 ml tanah
II. SURVEY & GEOGRAPHICAL DISTRIBUTION OF INDONESIAN POTATO CYST NEMATODE
Abstract
LISNAWITA. Survey and Geographical Distribution of Indonesian Potato
Cyst Nematode. Supervised by MEITY SURADJI SINAGA, GUSTAF ADOLF
WATTIMENA, SUPRAMANA and GEDE SUASTIKA
Potato cyst nematode (PCN) is a serious pathogen of potato and is found in most potato producing areas of the world. In Indonesia, PCN was only relatively recently discovered and new recognized as major constraint to potato production. A survey to determine the geographical distribution of PCN in Indonesia in 30 included potato production areas, i.e. 6 locations in East Java (survied in December 2004), 4 locations in Central Java (survied in October 2005) and 20 locations in West Java (survied in February 2006 and 2007). Based on this survey, PCN is widely established in East Java, Central Java and West Java, with population densities varying from low to high. In East Java, the highest population of PCN was found in Desa Tulung (Kota Batu District) with 675 cysts per 100 ml soil. However, two and three locations had only 44 and 21 cysts per 100 ml soil. In Central Java, PCN was detected in Pawuhan (Banjarnegara District) with the high population density 400 cysts per 100 ml soil. PCN was also found in Desa Karangtengah (Banjarnegara District), Desa Kepakisan (Banjarnegara District), and Desa Petak Bateng (Wonosobo District) with 270, 21 and 2 cysts per 100 ml soil, respectively. Populations of PCN were found in four locations in West Java, i.e. 3 locations in Desa Sukamanah (Bandung District) and one in Desa Mekarsari (Bandung District). The population densities from those four locations ranged from 2 to 28 cysts per 100 ml soil.
Pendahuluan
Nematoda sista kentang (NSK), Globodera spp. merupakan patogen penting pada pertanaman kentang yang menyebabkan kerusakan dan kehilangan hasil yang cukup besar di dunia (Evans & Turner 1998). Di Indonesia, keberadaan NSK pertama kali ditemukan menyerang pertanaman kentang di Dusun Sumber Brantas Desa Tulung Rejo Kecamatan Bumi Aji Kota Batu Jawa Timur. Daerah ini merupakan salah satu sentra penghasil kentang di Indonesia, sehingga budidaya kentang dilakukan secara besar-besaran. Oleh karena itu diperlukan ketersediaan bibit dalam jumlah yang besar. Untuk memenuhi kebutuhan bibit tersebut, banyak dilakukan impor bibit karena ketersediaan bibit di Indonesia belum memadai, baik secara kuantitas maupun kualitas.
Konsekuensi dari masuknya bibit kentang dari luar negeri ke wilayah Indonesia adalah masuknya organisme pengganggu tumbuhan yang belum ada di Indonesia, terutama yang bersifat tular bibit seperti NSK. NSK merupakan organisme pengganggu tanaman karantina (OPTK) karena potensi merusaknya yang sangat besar pada tanaman kentang, dan tersebar secara pasif bersama tanah atau umbi atau bahan pembiakan vegetatif yang lain. Selain itu, NSK merupakan salah satu patogen tanaman yang sulit dikendalikan, karena kemampuannya bertahan hidup yang sangat tinggi. Patogen ini mampu bertahan hidup di dalam tanah pada stadia dormansi yaitu berbentuk sista hingga mencapai 10 tahun (Taylor 1953 dalam Hamzah 2003) atau bahkan dapat mencapai lebih dari 20 tahun (Jones et al. 1998). Sifat dormansi sista NSK yang sangat panjang, juga dapat berimplikasi pada resistensi NSK di lahan-lahan yang telah terinfestasi.
Sebelum tahun 2003, NSK merupakan OPTK kategori A1, yang berarti tidak boleh masuk ke wilayah Indonesia, tetapi sejak tanggal 27 Januari 2006 berdasarkan Surat Keputusan Menteri Pertanian Nomor 38/Kpts/HK.060/1/ 2006, NSK masuk kategori A2 yaitu OPTK yang sudah berada di dalam Wilayah Negara Republik Indonesia dan dicegah pemencarannya ke daerah/pulau lain yang masih bebas.
Pada rapat kelompok kerja (Pokja) NSK pada 2-4 Mei 2007 di B&ung, dilaporkan saat ini penyebaran NSK semakin luas di Jawa Timur dan Jawa
Tengah. Di Jawa Timur saat ini petani tidak berani menanam kentang karena tanaman dapat mati pada umur satu bulan karena tingginya populasi NSK, sehingga terjadi penurunan luas pertanaman kentang dari 938,4 ha pada tahun 2003 menjadi 369 ha pada tahun 2006 (Anonimus 2007a). Di Jawa Tengah dilaporkan penyebaran NSK meluas dari 23 ha pada tahun 2003 menjadi 121 ha pada tahun 2006 (Suwardiwijaya et al. 2007).
Walaupun penyebaran NSK semakin luas, tetapi informasi tentang lokasi-lokasi penyebaran dan kepadatan populasi NSK masih sangat terbatas. Oleh karena itu perlu dilakukan survei ke lokasi-lokasi pertanaman kentang di Jawa Timur, Jawa Tengah dan Jawa Barat dan sehingga didapatkan pemetaan sebaran geografi NSK di Indonesia.
Tujuan Penelitian
Penelitian dengan metode survei ke sentra-sentra pertanaman kentang di Jawa Timur, Jawa Tengah dan Jawa Barat bertujuan untuk menentukan daerah sebar geografi NSK di Indonesia.
Metode Penelitian Tempat dan Waktu
Survei dilaksanakan di sentra pertanaman kentang di di Jawa Timur (Kabupaten Pasuruan dan Kota Batu), Jawa Tengah (Kabupaten Banjarnegara dan Wonosobo) dan Jawa Barat (Kabupaten Bandung). Semua sampel tanah dan akar dibawa dan dianalisis di Laboratorium Nematologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, IPB. Penelitian dilaksanakan pada bulan Desember 2004 sampai Maret 2007.
Pengumpulan Sampel Tanaman Kentang yang Terinfeksi
Sampel akar dan tanah diambil dari tanaman yang diduga terinfeksi NSK di lapangan dengan gejala daun menguning, layu dan pertumbuhan kerdil dari beberapa daerah pertanaman kentang di Jawa Timur (Kabupaten Pasuruan dan Kota Batu), Jawa Tengah (Kabupaten Banjarnegara dan Wonosobo) dan Jawa
Barat (Kabupaten Bandung). Pengambilan sampel tanah dan akar tanaman dilakukan dengan mengambil 100 ml contoh tanah pada kedalaman 0-20 cm. Lokasi hamparan pengambilan contoh tanah ditandai dengan GPS (Geo
Positioning System) untuk mengukur posisi geografis dan elevasi tanah. Pada
setiap lokasi diambil 5 (lima) contoh tanah yang mewakili kondisi lahan pada hamparan tersebut. Pengambilan contoh tanah dilakukan pada 30 lokasi hamparan yang tersebar di 3 (tiga) Propinsi yaitu Jawa Timur (6 lokasi), Jawa Tengah (4 lokasi), dan Jawa Barat (20 lokasi) (Gambar 2.2). Sampel tanah dan akar tanaman dimasukkan ke dalam kantung plastik, diberi label lokasi dan dimasukkan dalam kotak pendingin, selanjutnya dibawa ke Laboratorium Nematologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian IPB. Sebagai informasi tambahan, dari setiap sampel dicatat kultivar kentang dan umur tanaman.
Setiap sampel tanah diekstraksi dengan metode penyaringan (Shepherd 1985), sedangkan contoh akar dicuci dengan hati-hati kemudian dikeringanginkan. Selanjutnya sista yang ada pada sampel tanah dan akar dikumpulkan dengan kuas kecil atau jarum preparat dan jumlah sista per 100 ml tanah dihitung.
Hasil
Survei dan Pengambilan Sampel di lapangan
Berdasarkan hasil survei yang dilakukan di sentra produksi kentang di Jawa Timur, Jawa Tengah dan Jawa Barat diketahui NSK telah tersebar luas di beberapa wilayah survei. Di Jawa Timur NSK ditemukan pada 3 lokasi dari 6 lokasi yang diambil (50%), di Jawa Tengah NSK ditemukan pada ke-4 lokasi (100%), dan di Jawa Barat NSK ditemukan pada 4 lokasi dari 20 lokasi yang disurvei (20%). Hasil survei juga menunjukkan bahwa NSK tersebar pada ketinggian tempat yang berbeda yaitu mulai dari ketinggian 1400 m dpl sampai dengan 2100 m dpl. Disamping itu dari hasil survei diketahui NSK dapat menyerang semua kultivar kentang yang ditanam di lapangan, yaitu Granola, Granola Kembang dan Atlantik mulai dari tanaman muda hingga tanaman yang siap panen. Lokasi sebaran geografi NSK selengkapnya dapat dilihat pada Tabel 2.1.
B
A
C
G bar 2.1 Gejala NSK di lapangan :
Spot-spot kerdil dan daun kekuningan (A dan B).
Betina dan sista pada permukaan akar tanaman kentang (C) am
A
C
Gambar 2.2 Lokasi pengambilan sampel pada pemetaan dan penyebaran NSK di Jawa Timur, Jawa Tengah dan Jawa Barat
No. Lokasi Ketinggian tempat (m dpl)
Sista Umur
tanaman
Kultivar Kode Tabel 2.1 Daerah sebar NSK di Pulau Jawa dan kode isolat
isolat 1. Desa Tulung Rejo
Kota Batu Jawa Timur (lokasi 1)
1.800 675 >100 Granola S1
kembang
2. Desa Tulung Rejo Kota Batu Jawa Timur (lokasi 2)
1.700 44 40 Granola S2
kembang
3. Desa Tulung Rejo Kota Batu Jawa Timur (lokasi 3) 1.600 21 40 Granola S3 kembang 4. Desa Pawuhan Banjarnegara Jawa Tengah
1.900 400 Siap panen Granola S4
5. Desa Karangtengah Banjarnegara Jawa Tengah
1.700 270 60 Granola S5
6. Desa Patak Banteng Wonosobo Jawa 1.700 2 60 Granola S6 Tengah 7. Desa Kepakisan Banjarnegara Jawa Tengah
1.600 21 Siap panen Granola S7
8. Desa Sukamanah Pengalengan Jabar (lokasi 3) 1.508 2 50 Granola S8 9. Desa Sukamanah Pengalengan Jawa Barat (lokasi 4) 1.544 19 70 Granola dan Atlantik S9 10. Desa Sukamanah Pengalengan Jawa Barat (lokasi 5) 1.456 28 70 Granola S10 11. Desa Mekarwangi Sindangkerta Jawa Barat (lokasi 1) 1.343 17 60 Granola S11
Gejala tanaman ya an yang
ng terinfeksi di lapangan sama seperti gejala pada tanam terinfeksi nematoda pada umumnya. Infeksi NSK pada tanaman
m tin b pertu uhan terh at,
tanam ingan, layu dan mati (Gambar 2.1A dan Gambar 2.1B), sedangkan
infeksi yang terjadi pada masa generatif, tana kan bi
tetapi an berukuran kecil, dan bila tanaman dicabut akan
terlihat nematoda betina dan sista yang mene el pa ermu tan n kenta
ah dan sampel
menu K yang terendah (2 sista/100 ml tanah) sampai yang
tertinggi (675 sista/100 ml tanah) dapat dilihat pada Tabel 2.1. Kepadatan populasi NSK tertinggi terdapat pada Desa Tulung Rejo (Kabupaten Kota Batu, Jawa ), yaitu 675 sista/100 ml tanah, dan yang terendah terdapat
pada Desa Patak Banteng, (Kabupaten W sobo wa Tengah dan Desa
Sukam dung, Jawa Barat) (lokasi 3) masing-masing (2
ista/100 ml tanah).
Pembahasan
Hasil survei menunjukkan NSK telah menyebar luas di beberapa wilayah pada p
ung ejo (Kabupaten Kota Batu, Jawa Timur) (lokasi 1), Desa P
menyatakan NSK merupakan nematoda utama yang menyebabkan kerusakan pada uda dengan populasi yang
an kekun
ggi menyeba kan mb tanaman amb
man masih dapat menghasil um umbi yang dihasilk
mp da p kaan akar ama
ng (Gambar 2.1C). Hasil ekstraksi tan njukkan kepadatan NS
akar yang diambil dari lapangan
Timur) (lokasi 1
ono , Ja
anah (Kabupaten Ban s
ertanaman kentang di Jawa Timur, Jawa Tengah dan Jawa Barat. Infeksi NSK dapat terjadi pada semua umur tanaman, mulai tanaman muda hingga tanaman siap panen. Infeksi yang terjadi pada masa vegetatif ditandai dengan gejala daun menguning, layu, pertumbuhan terhambat dan akhirnya tanaman mati muda, sedangkan infeksi pada tanaman yang telah memasuki fase generatif tanaman masih dapat menghasilkan, tetapi umbi yang dihasilkan kecil dengan produksi di bawah rata-rata produksi normal. Pada populasi NSK yang cukup tinggi seperti di Desa Tul R
awuhan dan Desa Karangtengah (Kabupaten Banjarnegara, Jawa Tengah) masing-masing terdapat 675, 400 dan 270 sista per 100 ml tanah, sista dapat dilihat secara visual pada tanah dan tampak nematoda betina dengan warna keemasan menempel berderet pada akar tanaman. Stevenson et al. (2001)
pertanaman kentang di seluruh dunia dan mempunyai sebaran geografi yang sangat luas. Sampai saat ini NSK telah menyebar sedikitnya di 65 negara di seluruh dunia, meliputi negara-negara di seperti Eropa, Asia, Afrika, Amerika Utara, Amerika Tengah, Amerika Selatan dan negara-negara lain. Lebih lanjut Stevenson et al. (2001) menyatakan pada kepadatan populasi NSK yang tinggi gejala pada tanaman ditandai den an tanaman terhambat, tanaman kerdil, m
ebih luas, dibutuhkan monitoring kejadian penyakit secara intensif pada daerah-daerah yang
gan pertumbuh
enguning, layu dan mati muda. Pada akar tanaman yang terinfeksi dapat dijumpai nematoda betina yang berwarna putih hingga kekuningan dan sista yang berwarna coklat.
Terdeteksinya NSK di sentra-sentra pertanaman kentang di Jawa Timur, Jawa Tengah dan Jawa Barat diduga NSK sudah ada di daerah tersebut untuk waktu yang cukup lama. Seperti diketahui penggunaan bibit kentang impor di Jawa Timur sudah berlangsung dalam waktu yang sangat lama. Hal yang sama juga terjadi di Jawa Tengah. Di Jawa Tengah, petani telah menanam kentang sejak tahun 1985 dengan menggunakan bibit asal Jerman (Suwardiwijaya et al. 2007). Kondisi ini memungkinkan bagi NSK untuk mantap di daerah tersebut. Menurut Brodie (1984), untuk dapat terdeteksi dan menyebabkan endemik di suatu daerah, NSK memerlukan waktu sekitar 7 tahun. Hasil yang didapat dari survei ini, dapat menjadi ancaman serius bagi pertanaman kentang lain di Indonesia. Hal ini disebabkan karena NSK merupakan patogen yang sulit dikendalikan. Stevenson et
al. (2001) menyatakan, sekali NSK terinfestasi pada suatu lahan, maka nematoda
akan tetap ada di lahan tersebut dan tidak mungkin lahan tersebut dapat bersih dari NSK. Oleh karena itu, walaupun jumlah sista NSK di lokasi survei bervariasi dari rendah sampai tinggi, kondisi ini tidak menghalangi untuk terjadinya ledakan penyakit di lokasi tersebut. Di Inggris dilaporkan ambang ekonomi untuk G.
rostochiensis adalah 15 telur/ gram tanah. Di Jerman dilaporkan infestasi G. rostochiensis menimbulkan kerugian 11, 27 dan 43% pada kepadatan populasi
100, 1.000 dan 10.000 larva/100 cm3 tanah (CABI 2002). Untuk mencegah penyebaran NSK yang l
sudah terinfestasi maupun yang belum terinfestasi NSK dan kerja sama dari berbagai pihak yang terkait. Mengingat NSK dapat tersebar secara pasif bersama tanah atau umbi atau bahan pembiakan
vegetatif yang lain, maka masuknya bibit ke suatu daerah perlu pengawasan yang lebih ketat oleh Badan Karantina. Jika tidak ada pengawasan yang ketat, dikhawatirkan penyebaran NSK akan semakin luas.
Hasil survei menunjukkan bahwa semua kultivar kentang yaitu Granola, Granola Kembang dan Atlantik dapat terinfeksi NSK dengan berbagai ketinggian tempat. Hal ini menandakan bahwa sampai saat ini belum ada varietas kentang yang resisten NSK di Indonesia. Tingginya populasi NSK di Jawa Timur (lokasi 1) diduga disebabkan pola tanam petani kentang di daerah tersebut. Di Jawa Timur pola tanam yang digunakan petani adalah monokultur, tetapi lahan akan diberakan pada musim kemarau. Kondisi ini memungkinkan terputusnya siklus hidup patogen lain, tetapi NSK dapat berada dalam kondisi dorman di dalam tanah. Pada saat kentang ditanam kembali sista akan aktif dan menetas, kondisi ini akan meningkatkan populasi NSK. Di Jawa Tengah, petani menanam kentang di sepanjang tahun. Pola tanam seperti ini memungkinkan keberadaan patogen tanah yang lain berkembang baik, sehingga dapat menjadi kompetitor bagi NSK untuk mendapatkan nutrisi dan ruang untuk hidupnya. Kehadiran suatu spesies nematoda dapat mempengaruhi pertumbuhan spesies nematoda yang lain (Norton 1978). Secara umum, kompetisi yang lebih ketat terjadi antara spesies yang mempunyai kebiasaan makan yang sama. Miller & Wihrheim (1968) melaporkan infeksi yang disebabkan oleh Heterodera tabacum akan menurun bila di lokasi tersebut terdapat juga Pratylenchus penetrans atau
Tylenchorhynchus claytoni. Esteros & Chen (1972) menemukan kepadatan P. penetrans dan Meloidogyne incognita lebih kecil bila mereka ditemukan bersama
di dala akarm tanaman tomat dibandingkan jika mereka hidup secara tunggal. Kehadiran M. incognita dapat menghambat penetrasi P. penetrans jika kedua patogen ada bersama-sama pada akar tanaman.
Walaupun kepadatan populasi NSK di Jawa Timur lebih tinggi dari Jawa Tengah, tetapi penyebaran NSK di Jawa Tengah lebih cepat dari Jawa Timur. Meurut laporan Rapat Kerja NSK Nasional (Pokja) 2007, saat ini ada sekitar 121 ha pertanaman kentang di Jawa Tengah yang terinfeksi NSK. Penyebaran ini sangat cepat, bila dibandingkan pada tahun 2003, luas lahan yang terinfeksi baru sekitar 23 ha. Hal ini diduga karena penggunaan pestisida oleh petani di Jawa
Tengah sangat tinggi, sehingga menyebabkan pengaruh yang tidak baik bagi organisme tanah lainnya. Penggunaan pestisida tanpa aturan menyebabkan organisme bukan sasaran terutama organisme tanah yang menguntungkan ikut mati. Kondisi ini menyebabkan kompetitor maupun musuh alami NSK di daerah tersebut berkurang, sehingga menyebabkan NSK menyebar cepat.
Simpulan
Hasil survei yang dilakukan di lapangan, membuktikan bahwa NSK telah tersebar luas di beberapa wilayah di Jawa Timur, Jawa Tengah dan Jawa Barat, dengan kepadatan populasi rendah hingga tinggi.
Infeksi NSK dapat terjadi pada tanaman muda hingga tanaman menjelang panen, dari ketinggian tempat 1400 m dpl hingga 2100 m dpl. Kultivar kentang yang terinfeksi NSK pda saat survei dilakukan adalah Granola, Granola Kembang dan Atlantik.
Kepadatan populasi NSK tertinggi (675 sista/100 ml tanah) terdapat di Desa Tulung Rejo (Kabupaten Kota Batu, Jawa Timur) Lokasi 1, sedangkan kepadatan populasi terendah (2 sista per 100 ml tanah)cterdapat di Desa Sukamanah (Kabupaten Bandung, Jawa Barat) (lokasi 3) dan Desa Petak Bateng, (Kabupaten Wonosobo, Jawa Tengah).
Abstrak
LISNAWITA. Identifikasi dan Karakterisasi Nematoda Sista Kentang
Indonesia Menggunakan Karakter Morfologi, Molekuler dan Klon Diferensial. Dibimbing oleh MEITY SURADJI SINAGA, GUSTAF ADOLF
WATTIMENA, SUPRAMANA dan GEDE SUASTIKA
Di Indonesia, NSK adalah patogen baru yang dapat menjadi faktor penghambat produksi kentang. Pengendalian NSK yang efektif memerlukan informasi tentang spesies dan patotipe yang ada, apakah Globodera rostochiensis (patotipe Ro1, Ro2, Ro3, Ro4, Ro5, Ra1, Ra2, Ra3 dan Rb1) atau G. pallida (patotipe Pa1, Pa2, Pa3, Pa1, Pa2, Pa3, Pa4, Pa5 dan Pb1). Penelitian ini bertujuan untuk mengetahui spesies dan patotipe NSK dengan menggunakan karakter morfologi, molekuler dan klon diferensial. Empat populasi NSK, masing-masing 3 populasi dari Jawa Timur dan satu populasi dari Jawa Tengah digunakan untuk pengujian menggunakan karakter morfologi dan klon diferensial. Pengamatan dan pengukuran dilakukan pada sista, juvenil stadia dua (J2), betina, jantan dan telur. Sedangkan pada pengujian teknik molekuler digunakan teknik polymerase chain
reaction yang berdasarkan pada amplifikasi daerah internal transcribed spacer
dengan menggunakan dua primer spesifik. Sebelas populasi NSK, masing-masing 3 populasi dari Jawa Timur, 4 populasi dari Jawa Tengah dan 4 populasi Jawa Barat digunakan dalam penelitian ini. Hasil analisis berdasarkan pengamatan karakter morfologi berhasil diidentifikasi spesies tunggal G. rostochiensis. Teknik PCR memberikan hasil yang lebih cermat dengan terdeteksinya campuran spesies
G. rostochiensis (238 bp) dan G. pallida (391 bp) dari populasi NSK Jawa
Tengah. Sedangkan populasi NSK Jawa Timur dan Jawa Barat hanya terdeteksi satu spesies yaitu G. rostochiensis (238 bp). Berdasarkan analisis keragaman genetik dengan program Clustal-W 1,83 didapat isolat Indonesia berada pada kelompok yang berbeda dengan isolat NSK negara lain yang ada di GeneBank. Isolat Indonesia terbagi dalam dua kelompok, yaitu satu kelompok isolat Jawa Timur, dan satu kelompok isolat Jawa Tengah. Pengujian patotipe dengan klon diferensial didapat populasi NSK Jawa Timur menujukkan G. rostochiensis dengan patotipe baru spesifik Indonesia, sedangkan populasi NSK Jawa Tengah merupakan G. rostochiensis patotipe Ro1.
Kata kunci : Nematoda sista kentang, Globodera rostochiensis, G. pallida,
polymerase chain reaction, karakter morfologi
1. Bagian dari disertasi ini telah dipublikasikan dalam : 1). J. ISSAAS, Vol. 11, No. 3, December 2005 (Supplement), dengan judul : Species Identification of Indonesian Potato Cyst Nematode Using Polymerase Chain Reaction (PCR), 2). J. ISSAAS, Vol. 13, No. 1, June 2007, dengan judul : Technique Identification of Potato Cyst Nematode Populations Prevailing in East and Central Java, Indonesia, Based on Morphometric and Morphological Characteristics.
III. IDENTIFICATION AND CHARACTERIZATION OF INDONESIAN POTATO CYST NEMATODE USING MORPHOLOGICAL CHARACTERS, MOLECULAR
AND DIFFERENTIAL CLONES
Abstract
LISNAWITA. Identification and Characterization of Indonesian Potato Cyst Nematode Using Morphological Characters, Molecular, and Differential Clones. Supervised by MEITY SURADJI SINAGA, GUSTAF ADOLF
WATTIMENA, SUPRAMANA and GEDE SUASTIKA
Potato Cyst Nematode (PCN), a recently discovered a serious pathogen in some potato production areas in Indonesia, is a major constraint to production. To control PCN effectively, it is very important to identify its species and the pathotype in other countries too species with many pathotypes have been identified, Globodera rostochiensis (pathotypes Ro1, Ro2, Ro3, Ro4, Ro5, Ra1, Ra2, Ra3 and Rb1), and G. pallida (pathotypes Pa1, Pa2, Pa3, Pa1, Pa2, Pa3, Pa4, Pa5 and Pb1). Identification of PCN species was based on the morphological and molecular characters. Pathotype testing was conducted using differential clones. Four PCN populations were for morphology and host range, i.e. three populations from Sumber Brantas (Kota Batu District, East Java) and one from Karang Tengah (Banjarnegara District, Central Java). Observation and measurement were conducted on cysts, second stage juveniles (J2), females, and males. For molecular typing, 11 PCN populations from East Java, 4 populations from Central Java, and 4 populations from West Java were used. Polymerase Chain Reaction with specific structure (one for each Globodera species) used to amplify the internal transcribed spacer region of PCN ribosomal DNA. Based on morphological character, all specimens collected from research areas were identified as a single species of golden potato cyst nematode (G. rostochiensis). Whereas amplified DNA fragments, G. rostochiensis 238 bp and G. pallida 391 bp, indicated mixed populations occurred in Central Java. Only a single species, G.
rostochiensis, indicated in samples from East Java and West Java by the
molecular methods. Based on genetic character analysis with Clustal-W 1.83 program, it was found Indonesian populations had low similarity compared to PCN species lodged in GeneBank. Indonesian populations were separated into two groups, i.e. East Java and Central Java isolates. Clone differentials test showed that PCN population from East Java was G. rostochiensis of a previously undescribed Indonesian specific pathotype, while the population from Central Java was G. rostochiensis with pathotype similar to Ro1.
Key woards : Potato cyst nematode, Globodera rostochiensis, G. pallida, polymerase chain reaction, morphology character
1. Part of the dissertation has been publishes in 1). J. of ISSAAS, Vol. 11, No. 3,
December 2005 (Supplement), with title : Species Identification of Indonesian Potato Cyst Nematode Using Polymerase Chain Reaction (PCR), 2). J . of ISSAAS, Vol. 13, No. 1, June 2007, with title : Technique Identification of Potato Cyst Nematode Populations Prevailing in East and Central Java, Indonesia, Based on Morphometric
Pendahuluan
Spesies Globodera yang dilaporkan menyerang kentang, yaitu G. rostochiensis (nematoda sista kuning atau golden potato cyst nematode) dan G. pallida (nematoda sista putih atau pale potato cyst nematode) (Berg et al. 2000).
Masing-masing spesies mempunyai patotipe-patotipe yang respon patologiknya terhadap kultivar kentang berlainan. Sampai saat ini diketahui ada 8 patotipe yang digunakan di Eropa, masing-masing 5 patotipe dari G. rostochiensis dan 3 patotipe dari G. pallida (Kort et al. 1977) dan 10 patotipe yang digunakan di Amerika Selatan, masing-masing 4 patotipe dari G. rostochiensis dan 6 patotipe dari G. pallida (Canto Saenz & Scurrah 1977).
Penanaman bibit kentang dari berbagai negara oleh petani kentang di Indonesia menyebabkan peluang masuknya kedua spesies dengan beberapa patotipe cukup besar. Sayangnya, informasi spesies dan patotipe serta karakteristik patogenik dari nematoda sista kentang asal Indonesia sampai saat ini belum dilaporkan. Informasi spesies dan patotipe dibutuhkan sebagai dasar di dalam merancang strategi pengendalian yang tepat dan efektif.
Ada beberapa teknik yang dapat dilakukan untuk identifikasi spesies NSK, yaitu secara konvensional, misalnya dengan pengamatan dan pengukuran ciri-ciri morfologi (morfometri), uji serologi (Schots 1988) dan teknik molekuler misalnya dengan menggunakan sidik jari DNA melalui PCR-RFLP (Fleming & Powers 1998). Sedangkan untuk mengetahui patotipe-patotipe NSK dapat dilakukan dengan pengujian menggunakan klon diferensial.
Morfologi dan morfometri masih digunakan bersama-sama dengan teknik molekuler untuk identifikasi spesies nematoda sista kentang (Baldwin & Mundo-ocampo 1991). G. rostochiensis dan G. pallida secara morfologi dan morfometri hanya memiliki sedikit perbedaan (Stone 1973a & 1973b). Oleh karena itu pengamatan beberapa karakter pada sista dan juvenil stadia dua (J2) merupakan kegiatan yang paling penting yang direkomendasikan untuk identifikasi spesies NSK secara konvensional. Kedua stadia ini selalu ditemukan pada sampel tanah yang telah terinfestasi NSK, di samping itu juga dilakukan pengamatan terhadap betina, jantan dan telur.
Klon diferensial atau tanaman indikator, merupakan salah satu cara untuk mengkarakterisasi masing-masing isolat berdasarkan kesesuaian pada suatu inang. Pada NSK klon diferensial mempunyai peranan yang sangat penting untuk membedakan patotipe. Patotipe tertentu hanya mampu bermultiplikasi pada inang tertentu, sehingga terdapat hubungan antara spesies yang resisten dengan patotipe NSK. Gen resisten pada tanaman kentang ada yang bersifat monogenik (gen major) atau poligenik (gen minor). Gen-gen monogenik tersebut adalah gen H1, H2, H3, gen A, B, dan gen Fa, Fb. Kespesifikan tanaman inang NSK menyebabkan pengujian klon diferensial sering dilakukan untuk karakterisasi spesies yang belum diketahui (Fleming & Powers 1998 ; Hull 2002).
Saat ini untuk keperluan identifikasi lebih cepat dan akurat, telah dikembangkan teknik identifikasi dengan menggunakan pendekatan biologi molekuler. Metode ini menggunakan struktur asam nukleat sebagai dasar untuk mengkarakterisasi dan mengidentifikasi patogen tanaman. Identifikasi nematoda sista kentang banyak menggunakan teknik polymerase chain reaction (PCR).
Polymerase chain reaction (PCR) adalah suatu metode in vitro untuk
mensintesis asam nukleat berdasarkan cetakan DNA. Dengan teknik PCR masalah jumlah DNA yang rendah per individu dapat teratasi. Fragmen DNA genom nematoda yang menjadi sasaran analisa diamplifikasi terlebih dahulu dengan PCR. Hasil analisis genom nematoda, adalah informasi bahwa bagian internal
transcribed spacer (ITS) dari rDNA merupakan daerah yang sangat variabel
sehingga dapat berfungsi sebagai penanda spesifik dalam studi taksonomi molekuler dan pilogenik sehingga sering digunakan untuk analisa patotipe suatu populasi nematoda (Thiery & Mugniery 1996).
Pengujian dengan teknik PCR memerlukan dua primer oligonukleotida spesifik yang akan menginduksi pembentukan dan perbanyakan asam nukleat atau untai DNA dengan bantuan enzim Taq polymerase dalam mesin PCR atau
thermocycler (Saiki 1990). Pemilihan primer yang tepat sangat menentukan
keberhasilan identifikasi suatu patogen. Dalam bidang fitopatologi, teknik PCR banyak digunakan untuk tujuan deteksi patogen, identifikasi patogen, karekterisasai keanekaragaman patogen, maupun untuk diferensiasi patogen tumbuhan.
Tujuan Penelitian
Tujuan penelitian ini adalah mengidentifikasi dan mengkarakterisasi spesies NSK dengan menggunakan karakter morfologi dan molekuler serta mengetahui patotipe NSK dengan menggunakan klon diferensial.
Metode Penelitian Tempat dan Waktu
Penelitian dilakukan di Laboratorium Nematologi Tumbuhan dan Laboratorium Virologi Tumbuhan Departemen Proteksi Tanaman Fakultas Pertanian IPB serta rumah kassa Kebun Percobaan desa Cipelang Cijeruk, dengan ketinggian tempat 1250 m dpl. Penelitian dilaksanakan dari bulan Agustus 2005 sampai Maret 2007.
Identifikasi Spesies NSK berdasarkan Karakter Morfologi Populasi nematoda sista kentang
Untuk identifikasi spesies dengan menggunakan karakter morfologi dan morfometri tidak semua isolat NSK digunakan. Dari 11 isolat NSK yang ada, hanya 4 isolat NSK yang digunakan pada percobaan ini, yaitu isolat-isolat yang ditemukan pada survei tahun pertama (S1, S2, S3 dan S4). Sedangkan isolat-isolat lainnya tidak diidentifikasi dengan karakter morfologi karena kepadatan populasi yang rendah dan akan diidentifikasi dengan menggunakan karakter molekuler.
Dua puluh lima betina dan 25 sista dikumpulkan pada tempat terpisah. Pembuatan preparat untuk betina dilakukan dengan memotong betina menjadi dua bagian, kemudian masing-masing bagian diletakkan pada gelas objek yang telah ditetesi dengan Hoyer (30 gram gum arabik, 200 gram kloral hidrat, 16 gram gliserol dan 200 ml akuades) lalu ditutup dengan gelas penutup.
Pembuatan preparat untuk sista dilakukan dengan memotong bagian posterior sista dengan menggunakan pisau bedah, kemudian telur dan juvenil yang ada di dalamnya dibersihkan dengan jarum pancing nematoda, setelah itu
potongan sista dipindahkan ke atas gelas objek yang telah ditetesi dengan Hoyer dan ditutup dengan gelas penutup.
Pembuatan preparat untuk juvenil stadia 2 (J2), jantan dan telur dilakukan dengan mengumpulkan masing-masing 50 J2, 50 jantan dan 50 telur pada tempat terpisah, kemudian nematoda dimatikan dengan pemanasan lalu diletakkan pada gelas objek yang telah ditetesi lactophenol cotton blue dan kemudian ditutup dengan gelas penutup.
Pengamatan
Pengukuran dilakukan dengan mengambil gambar setiap sampel di bawah mikroskop Olympus BX 51 dengan perbesaran 40x (10 x 4) untuk sista dan betina. Perbesaran 200x (10 x 20) untuk J2, jantan dan telur dengan menggunakan kamera digital mikroskop Olympus DP 11. Selanjutnya foto ditransfer menggunakan program morfometri tpsdig (Bennet & Hoffman 1998). Penentuan titik (digitasi) pada setiap gambar dilakukan dengan menentukan secara konsisten titik-titik masing-masing variabel. Setiap titik dari gambar pemotretan digitasi diubah dalam kordinat x dan y sehingga dapat diketahui jarak antar titiknya, kemudian dimasukkan dalam persamaan jarak dengan menggunakan program Microsoft Exel, sehingga diperolah jarak yang sesungguhnya, yaitu :
DV(mm) =
√
((X1 – X2)2 + (Y2 – Y2)2) (persamaan jarak-1)DS(mm) = DV / DP (persamaan jarak-2)
Dimana :
DV(mm) = jarak vektor,
DS(mm) = jarak sesungguhnya,
DP = jarak perbesaran mikroskop,
X1, X2, Y1, Y2 = titik-titik vektor pada sumbu X dan Y
Hasil digitasi setiap variabel berbentuk vektor yang kemudian dikonversi dalam ukuran sesungguhnya dengan cara nilai vektor dibagi dengan 399,99. Nilai ini diperoleh dari penentuan titik skala mikrometer (sepanjang 1 mm) pada perbesaran yang sama saat pemotretan , yaitu 10 x 20 (200x) atau 10 x 4 (40x).
Konfirmasi spesies dilakukan berdasarkan kunci determinasi Fleming & Powers (1998) ; Stone (1973a) ; Stone (1973b).
Pengamatan dan pengukuran dilakukan terhadap sista, betina, juvenil stadia 2 (J2), jantan dan telur dengan variabel seperti di bawah ini :
Betina
Pengamatan pada bagian anterior dilakukan dengan mengukur lebar kepala yang diukur pada dasar kepala dan panjang stilet, sedangkan pengamatan pada bagian posterior dilakukan dengan mengukur rata-rata diameter fenestra dan jarak dari anus ke fenestra.
Sista
Pengamatan dilakukan dengan menggunakan mikroskop stereo dan kompaun dengan melakukan pengukuran terhadap lebar sista, panjang sista termasuk leher, panjang leher, jumlah tonjolan kutikula, rasio granek yaitu jarak antara fenestra dengan anus dibagi dengan diameter fenestra, diameter fenestra dan jarak dari anus ke fenestra.
Juvenil stadia dua (J2)
Pengamatan pada J2 dilakukan dengan mengukur panjang dan lebar tubuh, panjang dan lebar kepala, panjang stilet, bentuk tonjolan stilet, jarak dari kepala ke median bulb, panjang dan lebar ekor.
Jantan
Pengamatan pada jantan dilakukan dengan mengukur panjang tubuh, panjang dan lebar kepala, panjang stilet, panjang spikula dan panjang ekor.
Telur
Pengamatan pada telur dilakukan dengan mengukur panjang dan lebar telur. Analisis ragam dilakukan terhadap data hasil pengamatan dengan menggunakan uji F dan jika diantara perlakuan terdapat perbedaan dilanjutkan dengan Uji Jarak Berganda Duncan (DMRT) pada ά=0,05 dengan bantuan SAS 6.12.
Identifikasi Spesies NSK berdasarkan Karakter Molekuler Ekstraksi DNA
Identifikasi NSK dengan menggunakan teknik PCR dilakukan untuk semua sampel yang positif NSK (Bab. II, Tabel 2.1). Metode ekstraksi DNA disusun berdasarkan metode Fullaondo et al. (1999) ; Subbotin et al. (2001) yang dimodifikasi. Dua puluh lima sista dikumpulkan secara acak dari setiap sampel, kemudian dimasukkan ke dalam eppendof steril yang berisi 150 μl buffer lisis (125 mM KCl ; 25 mM Tris-HCl, pH 8,0 ; 3,75 mM MgCl2 ; 2,5 mM DTT ; 1,125% Tween 20 dan 0,025% gelatin) dan ditambahkan 5 μl Proteinase K (600 μg/ml) (USB, UK). Sista digerus dengan pistil mikro plastik selama 2-3 menit, divorteks dan diinkubasi pada suhu 65°C selama 1 jam, dilanjutkan pada suhu 95°C selama 10 menit, setelah itu disentrifugasi dengan kecepatan 11000 rpm selama 10 menit. Supernatan dipindahkan pada tabung baru dan ditambahkan 1 volume kloroform : isoamil alkohol (24:1), divorteks dan disentrifugasi dengan kecepatan 11000 rpm selama 10 menit. Supernatan dipindahkan ke tabung baru dan ditambahkan 2.5 kali volume total supernatan NAOAc 3 M (pH 5.2), kemudian diinkubasi selama 20 menit pada suhu -20ºC. Supernatan dibuang setelah disentrifugasi 14000 rpm selama 10 menit. Pelet dicuci dengan 500 μl etanol 70% dan disentrifugasi 14000 rpm selama 10 menit. Etanol dibuang kemudian pelet dikeringkan di dalam pompa vakum selama 10 menit, selanjutnya pelet diresuspensi dengan 20 μl ddH2O. Jika DNA belum segera digunakan, dapat
disimpan pada temperatur -20°C. DNA hasil ekstraksi diamplifikasi dengan teknik PCR berdasarkan metode Fullaondo et al. (1999).
Setiap reaksi PCR (25 μl) terdiri atas 25 ng DNA template dari masing-masing sampel, yang terdiri dari 50 mM Tris-HCl (pH 9,0) ; 50 mM KCl ; 1,5 mM MgCl2 ; 0,1% Triton X-100 ; 0,2 mM setiap dNTP (New England Biolabs) ; 50 ng setiap primer dan 0,5 unit Taq polymerase (New England Biolabs).
Tiga primer digunakan, masing-masing primer spesifik untuk G.
rostochiensis yaitu ITS-1 (5’- TGT TGT ACG TGC CGT ACC TT-3’), primer
GT-3’) dan primer universal untuk NSK yaitu 5,8S rRNA (5’- GCA GAA GGC TAG CGA TCT TC-3’) (Invitro gen) (Mulholland et al. 1996).
Amplifikasi DNA dilakukan dengan denaturasi awal pada 96ºC selama 2 menit, kemudian dilanjutkan dengan 35 siklus yang melalui tiga tahapan, yaitu pemisahan utas DNA (denaturation) pada 94°C selama 1 menit, penempelan primer (anneling) pada 50°C selama 1 menit dan sintesis DNA (extention) pada
72°C selama 2 menit (Modifikasi Mulholland et al. 1996). Khusus untuk siklus
terakhir ditambah tahapan sintesis selama 7 menit, kemudian siklus berakhir dengan suhu 4ºC.
Tujuh μl fragmen DNA hasil amplifikasi PCR dianalisis dengan elektroforesis pada 2% gel agarose dalam buffer Trisborate EDTA (TBE) 1X dengan tegangan 100 Volt selama 70 menit (Subbotin et al. 2001) dan diamati dengan UV transiluminator setelah diberi warna dengan ethidium bromida (Maniatis et al. 1989).
Sekuensing
Sampel fragmen DNA hasil amplifikasi PCR disekuensing untuk melihat susunan DNA masing-masing populasi. Sekuensing dilakukan di Laboratorium
Research and Development Centre PT. Charoen Pokhpand, Jakarta dengan
menggunakan mesin sekuensing ABI-Prism 3100-Avant Genetic Analyzer.
Analisis Filogenetik
Hasil sekuensing populasi NSK yang digunakan dalam penelitian selanjutnya dianalisis tingkat kesamaannya dengan cara membandingkannya dengan sekuen NSK lain yang ada di GeneBank (Tabel 3.1). Analisis filogenetik dan jarak genetik dilakukan dengan menggunakan program Clustal-W 1.83.
Tabel 3.1 Lokasi geografi dan nomor asesi sekuen NSK pada GeneBank Identifikasi berdasarkan sekuen ITS Identifikasi original
Populasi Akronim Sumber (no asesi Genebank)
G. rostochiensis G. rostochiensis
Desa Tulung Rejo, Kecamatan Bumi Aji, Kabupaten Batu, Jawa Timur (lokasi 1), Indonesia
Desa Tulung Rejo, Kecamatan Bumi Aji, Kabupaten Batu, Jawa Timur (lokasi 2), Indonesia
Desa Tulung Rejo, Kecamatan Bumi Aji, Kabupaten Batu, Jawa Timur (lokasi 3), Indonesia
Desa Patak Banteng, Kecamatan Kejajar, Kabupaten Wonosobo, Jawa Tengah, Indonesia Desa Kepakisan, Kecamatan Batur, Kabupaten Banjarnegara, Jawa Tengah, Indonesia Kanada Falkland Islands, UK Cuapiaxtla, Meksiko
New York, USA
Allpachaka, Peru Peru Anta, Peru Liberice, Slovenia Gr1-JT1 Gr2-JT2 Gr3-JT3 Gr6-JTH Gr7-JTH Gr-Kan Gr-FI Gr-Meks Gr-NY Gr-Peru1 Gr-Peru2 Gr-Peru3 Gr-SA Penelitian ini Penelitian ini Penelitian ini Penelitian ini Penelitian ini Szalanski et al. (unpubli) (AFO16875) Szalanski et al. (unpubli) (AFO16876) Szalanski et al. (unpubli) (AFO16877) Szalanski et al. (unpubli) (AFO16878) Szalanski et al. (unpubli) (AFO16872) Szalanski et al. (unpubli) (AFO16873) Szalanski et al. (unpubli) (AFO16874) Szalanski et al. (unpubli) (AFO16874)
Pengujian Patotipe NSK dengan Klon Diferensial
Untuk mengetahui patotipe NSK dari empat isolat yang digunakan pada identifikasi karakter morfologi di atas, dilakukan pengujian klon diferensial. Klon diferensial yang digunakan adalah klon diferensial komersil yang merupakan pengganti dari klon diferensial klasik (Kort et al. 1977), yaitu kultivar Maritta, Saturna, KTT 60.12.19, ODV 32731, S. verrnei hibrid 62.33.3, AM 78.3778 dan P55/7.
Semua klon diferensial diperbanyak secara kultur jaringan dengan media perbanyakan MS0 (Lampiran 1). Planlet yang berumur dua minggu diaklimatisasi di rumah kassa Kebun Percobaan desa Cipelang, Cijeruk, dengan ketinggian tempat 1250 m dpl. Setelah itu tanaman dipindahkan ke dalam kantong plastik berukuran 30 cm x 30 cm dengan media tanah : pasir steril (1:1 / v:v). Untuk menjaga pertumbuhan tanaman dilakukan dengan pemberian pupuk NPK (15:15:15) (3 g/l) dan Bayfolan (2 ml/l) seminggu sekali dan disemprot insektisida (jika perlu). Inokulasi dilakukan setelah tanaman berumur dua minggu dengan menyebarkan 25 sista NSK di sekitar perakaran tanaman (Foot 1977).
Pengamatan dilakukan 12 minggu setelah inokulasi dengan membongkar tanaman secara hati-hati. Tanah diekstraksi dengan metode penyaringan (Shepherd 1985), sedangkan akar dicuci dengan hati-hati kemudian dikeringanginkan. Pengamatan dilakukan dengan menghitung laju reproduksi sista (Rf) yaitu dengan membandingkan jumlah sista pada akhir penelitian (Pf) dengan jumlah sista pada awal penelitian (Pi). Hasil yang didapat dibandingkan dengan skema yang dikemukan Tabel 3.2 di bawah ini.
Tabel 3.2 Skema Internasional untuk identifikasi patotipe G. rostochiensis (Kort
et al. 1977; Canto Saenz & de Scurrah 1977) pada klon diferensial
komersil.
Patotipe
Ra1 Rb1 Ra2 Ra3
Klon diferensial
komersil Tetua
Ro1 Ro4 Ro2 Ro3 Ro5
Maritta S.t.tuberosum + + + + +
Saturna S.t.andigena CPC 1673 – – + + +
KTT 60.21.19 S.kurtzianum hib.60.21.19 – + – + +
ODV 32731 S.vernei hib. 58.1642/4 – + – – +
S.vernei hib. 62.33.3 S.vernei hib. 62.33.3 – + – – +
AM 78.3778 S.vernei hib. 65.346/19 – – – – –
Keterangan : + = Pf/Pi >1
– = Pf/Pi ≤ 1
Hasil
Karakter Morfologi NSK Indonesia
Hasil identifikasi menggunakan karakter morfologi pada S1, S2, S3 dan S4 dapat dilihat pada Gambar 3.1 dan 3.2.
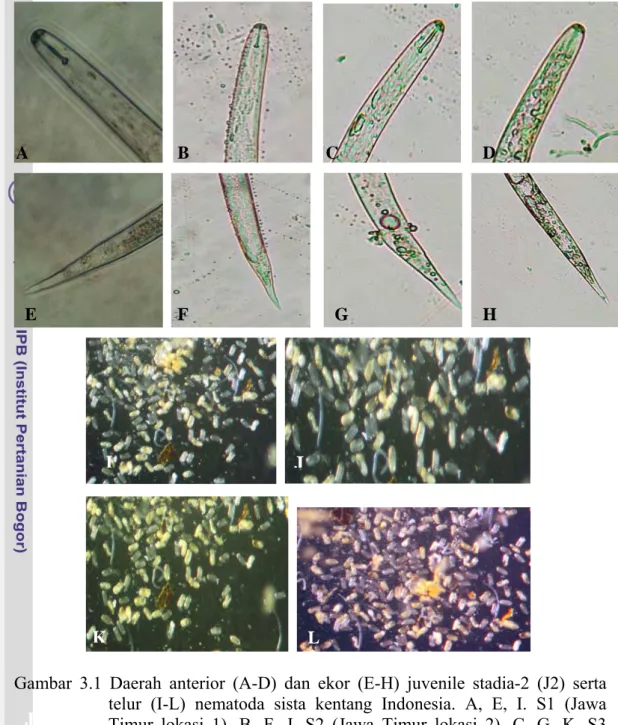
Juvenil stadia-2 (J2) (Gambar 3.1 A-G)
Hasil identifiksi menggunakan karakter morfologi pada J2 menunjukkan tidak ada perbedaan bentuk tubuh dari ke-4 sampel NSK yang digunakan. Tubuh J2 berbentuk seperti cacing. Kepala berbentuk bulat dengan kerangka yang kuat. Stilet berkembang baik dan kuat dengan tonjolan stilet berbentuk bulat pada keempat sampel. Bentuk ekor makin ke ujung makin mengecil. Sebagian tubuh posterior tampak hialin. Tidak terlihat perbedaan yang jelas dari pengamatan secara morfologi diantara keempat sampel yang digunakan.
Telur (Gambar 3.1 I-L)
Telur berada di dalam sista. Jika sista dipecahkan, maka akan terlihat jumlah telur yang cukup banyak. Telur berbentuk oval. Dinding telur halus dan tidak terdapat mikrovilli.
A B C D E F G H J I
Gambar 3.1 Daerah anterior (A-D) dan ekor (E-H) juvenile stadia-2 (J2) serta telur (I-L) nematoda sista kentang Indonesia. A, E, I. S1 (Jawa Timur lokasi 1). B, F, J. S2 (Jawa Timur lokasi 2). C, G, K. S3 ( Jawa Timur lokasi 3). D, H, L). S4 (Pawuhan Jawa Tengah).
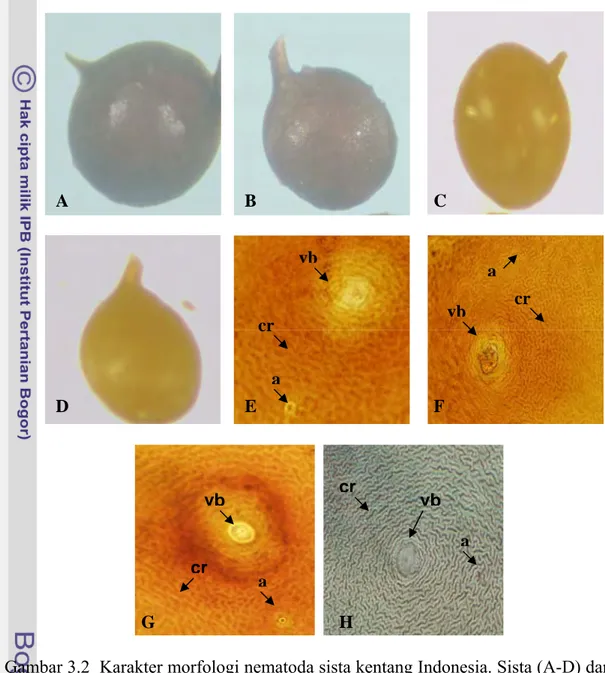
ambar 3.2 C-D)
Tubu
cone pada ke
erwarna putih, kemudian berubah menjadi kuning keemasan. Permukaan tubuh
K L J L I Betina (G I
h betina berbentuk bulat, sedikit ellips atau lonjong, tanpa vulval empat sampel. Tubuh betina yang baru keluar dari korteks awalnya b
bagian posterior betina ditutupi tonjolan kutikula. Sama seperti sista tonjolan kutikula menyatu dengan lehernya yang pendek (Gambar 3.2 C-D).
G D E F A B C cr vb a cr vb a cr vb vb cr a a H
Gambar 3.2 Karakter mo atoda sista kentang Indonesia. Sista (A-D) dan sidik pantat (perineal pattern) (E-H). a Timur lokasi 1). B, F). S2 (Jawa Timur lokasi 2). C, G). S3 (Jaw ur lokasi 3). D, H). S4 (Pawuhan Jawa Tengah). (vb = vulva basin, cr = cutcular
ridges, a=anus,GR besar dari 3,2)
rfologi nem
A, E). S1 (Jaw a Tim
Sista (Gam
Sama seperti betina, keempat sampel mempunyai sista dengan bentuk membulat, sedikit ellips atau ovoid. Tonjolan kutikula pada sista mempunyai pola yang sama seperti pada betina. Sista merupakan betina yang mati dengan kutikula ang keras, di dalamnya berisi telur yang akan menjadi generasi berikutnya. Sista g kemudian berubah menjadi coklat tua atau kehitaman.
Berdasarkan pengamatan morfologi dengan ciri-ciri seperti yang n di atas, maka keempat sampel NSK tersebut adalah G. rostochiensis.
g-masing isolat di atas. Hasil pengukuran
k erbeda nyata pada keempat isolat uji. Hasil yang sama juga didapat pada betina. Sedang
tonjolan stilet yang berbentuk bulat. Penguk
imur dan Jawa Tengah berdasarkan morfometrinya.
bar 3.2 A-B)
y
berwarna coklat muda yan
Tidak terdapat vulval cone, leher dan kepala kecil dan menonjol.
Jantan
Pada keempat sampel didapatkan jantan berbentuk cacing (vermiform). Tubuh jantan akan berbentuk C atau S jika dipanaskan atau difiksasi. Ekor terlihat tumpul dengan spikula di dekat ujung.
dijelaska
Di samping pengamatan morfologi, dilakukan juga pengukuran pada beberapa variabel morfologi dari masin
dapat dilihat secara lengkap pada Tabel 3.3.
Berdasarkan uji jarak Duncan, semua karakter yang diukur pada sista tida b
kan pada J2 didapat panjang stilet dan lebar tubuh S1 berbeda nyata dengan S2, S3 dan S4, tetapi pada karakter yang lain semua isolat tidak berbeda nyata, selain itu keempat isolat mempunyai
uran pada jantan dan telur juga didapat beberapa karakter berdasarkan uji jarak Duncan berbeda nyata antar isolat
Selanjutnya dilakukan analisis berdasarkan wilayah yaitu membandingkan isolat Jawa Timur (S1, S2 dan S3) dengan isolat Jawa Tengah (S4) (Tabel 3.4). Berdasarkan uji jarak Duncan didapat isolat Jawa Timur (S1, S2 dan S3) secara umum tidak berbeda nyata dengan isolat Jawa Tengah (S4), dengan kata lain tidak ada perbedaan antara isolat Jawa T
Tabel 3.3 Ukuran sista, juvenil, betina, jantan dan telur NSK isolat S1 (Jawa Timur lokasi 1), S2 (Jawa Timur lokasi 2), S3 (Jawa Timur lokasi 3) dan S4 (Pawuhan Jawa Tengah) (μm)
Sampel NSK * No
Sista
. Karakter S1 S2 S3 S4
1. Lebar 371,0 ± 103,8a 380,0 ± 96,5a 378,0 ± 95,0a 374,0 ± 6 2. Panjang
6,5a 456,0 ± 31,8a 457,0 ± 33,4a 470,0 ± 44,3a 450,0 ± 37,1a 3. Di eter fenestra 19,4 ± 4,8a 19,0 ± 4,6a 20,0 ± 4,2a 19,0 ± 4,6a ± 8,3a
5. Panjan ± 47,8a
6. Rasio g 8 ± 1,1a 3,6 ± 0,9a 3,9 ± 0,9a
ticular ridges 28,0a
,0 ) 3 ,0 ) 0 ) 30,0a 0 ) il stadia dua (J2) g tubuh uh
ilet bulat bulat bulat bulat
Betin
val basin
a
b
20. 5,7 ± 0,5a 5,5 ± 0,5a 5,8 ± 0,7a 5,6 ± 0,8a 6,5 b 6, b 6, b 7 8a ebar kepala 12,0 ± 1,5a 11,7 ± 1,5a 13,1 ±1,3a 12,0 ±1,4a
am
4. Jarak (anus – fenestra) 65,0 ± 10,6a 66,0 ± 8,3a 65,0 ± 7,0a 66,0 g leher 104,0 ± 79,2a 104,0 ± 79,2a 110,0 ± 75,7a 105,0
ranek 4,4 ± 1,8a 3, 7. Jumlah cu (rata-rata) 16,0 – ( 22 15,0 – ( 20 0,0a 16,0 – 32,0a ( 21, 14,0 – ( 23, Juven
8. Panjan 457,0 ± 97,9a 452,0 ± 96,9a 462,0 ±90,5a 460,0 ± 81,5a 9. Lebar tub 17,5 ± 1,1ab 17,2 ± 0,9b 17,8 ± 0,9a 17,7 ± 0,6a 10. Panjang stilet 24,2 ± 1,1a 23,0 ± 1,3b 23,0 ± 1,7b 22,0 ± 1,4c 11. Panjang ekor 45,4 ± 5,0a 44,0 ± 5,2a 45,0 ± 4,3a 45,0 ± 4,6a 12. Lebar ekor 11,5 ± 1,4a 11,7 ± 1,4a 12,0 ± 1,1a 11,8 ± 1,4a 13. Lebar kepala 4,3 ± 1,2a 4,3 ± 1,2a 4,5 ± 0,5a 4,7 ± 0,9a 14. Panjang bagian hialin
(pada ekor)
27,6 ± 4,1a 28,0 ± 3,5a 27,0 ± 4,2a 27,0 ± 3,9a
15. Bentuk tonjolan st
a
16. Diameter vul 21,0 ± 4,0a 20,0 ± 3,4a 22,0 ± 3,1a 21,0 ± 3,8a
Jant n
17. Panjang tubuh 898,0 ± 0,2b 875,0 ± 0,9b 1059,0 ± 0,4a 1109,0 ± 0,3a 18. Lebar tubuh 28,0 ± 2,6a 27,9 ± 3,0a 27,2 ± 2,7a 27,4 ± 3,0a 19. Panjang stilet 24,3 ± 1,5b 25,3 ± 2,3a 25,4 ± 2,1a 25,0 ± 2,1a
Panjang ekor
21. Panjang kepala ± 0,9 5 ± 0,9 8 ± 0,6 ,3 ± 0,
22. L
23. Panjang spikula 36,0 ± 2,1a 32,4 ± 4,2b 30,0 ± 5,7b 31,0 ±5,5b
Telur 24. 25. Panjang Lebar 105,0 ± 2,70a 47,0 ± 4,6bc 104,0 ± 4,2a 45,0 ± 3,0c 102,8 ± 1,3b 50,9 ± 0,9a 103,8 ± 1,7b 48,4 ± 1,3a
* Ra (sista), 50 tina) dan
ti dengan h a p g s bed
berdasarkan uji DMRT
ta-rata dari 25 (J2), 25 (be , 50 (jantan), 25 (telur) Ang
Tabe sista, j , j elur sark
h Jawa T dan Te
NS l 3.4 Ukuran
wilaya uvenil, betinaimur (S1, S2 antan dan t S3) dan Jawa ngah (S4) (μm) NSK berda an Sampel K *
No. Karakter Jawa Timur (S1, S2, S3) Jawa Tengah (S4)
Sista
1. Lebar 376,3 a
g 61,0 a
6. Rasio gra
mlah cuticular ridges 15,0 – 32,0 14,0 – 30,0a
il stadia dua (J2)
g tubuh uh
ilet Bulat Bulat
Betin val basin a 21. la 6,6 ± 0,8a 7,3 ± 0,8a 1,6a a ng spikula 32,8 ± 4,9b 31,0 ±5,5b Telur njang 103,9 ± 3,0b 103,8 ± 1,7b ± 97,3a 374,0 ± 66,5 2. Panjan 4 ± 37,0a 450,0 ± 37,1
3. Diameter fenestra 19,5 ± 4,5a 19,0 ± 4,6a 4. Jarak (anus – fenestra) 65,3 ± 8,7a 66,0 ± 8,3a
5. Panjang leher 106,0 ± 77,5a 105,0 ± 47,8a
nek 3,9 ± 1,4a 3,9 ± 0,9a
7. Ju (rata-rata)
a
(21,0) ( 23,0 )
Juven
8. Panjan 457,0 ± 94,6a 460,0 ± 81,5a
9. Lebar tub 17,5 ± 1,0a 17,7 ± 0,6a
10. Panjang stilet 23,4 ± 1,5a 22,0 ± 1,4c
11. Panjang ekor 44,8 ± 4,9a 45,0 ± 4,6a
12. Lebar ekor 11,7 ± 1,3a 11,8 ± 1,4a
13. Lebar kepala 4,4 ± 1,0a 4,7 ± 0,9a
14. Panjang bagian hialin )
(pada ekor
27,5 ± 3,9a 27,0 ± 3,9a
15. Bentuk tonjolan st
a
16. Diameter vul 21,0 ± 3,6a 21,0 ± 3,8a
Jant n
17. Panjang tubuh 1048,0 ± 130,9a 1109,0 ± 0,3a
18. Lebar tubuh 27,7 ± 4,8a 27,4 ± 3,0a
19. Panjang stilet 25,0 ± 2,0a 25,0 ± 2,1a 20. Panjang ekor 5,7 ± 0,6a 5,6 ± 0,8a
Panjang kepa
22. Lebar kepala 12,3 ± 12,0 ±1,4
23. Panja
24. Pa
25. Lebar 47,6 ± 4,0a 48,4 ± 1,3a
* Ra (sista), 50 (J2), 25 (be ), dan 25
ti dengan huruf yang sa ang sam a
nyata pada taraf 5% berdasarkan uji DMRT
ta-rata dari 25 tina), 50 (jantan (telur)
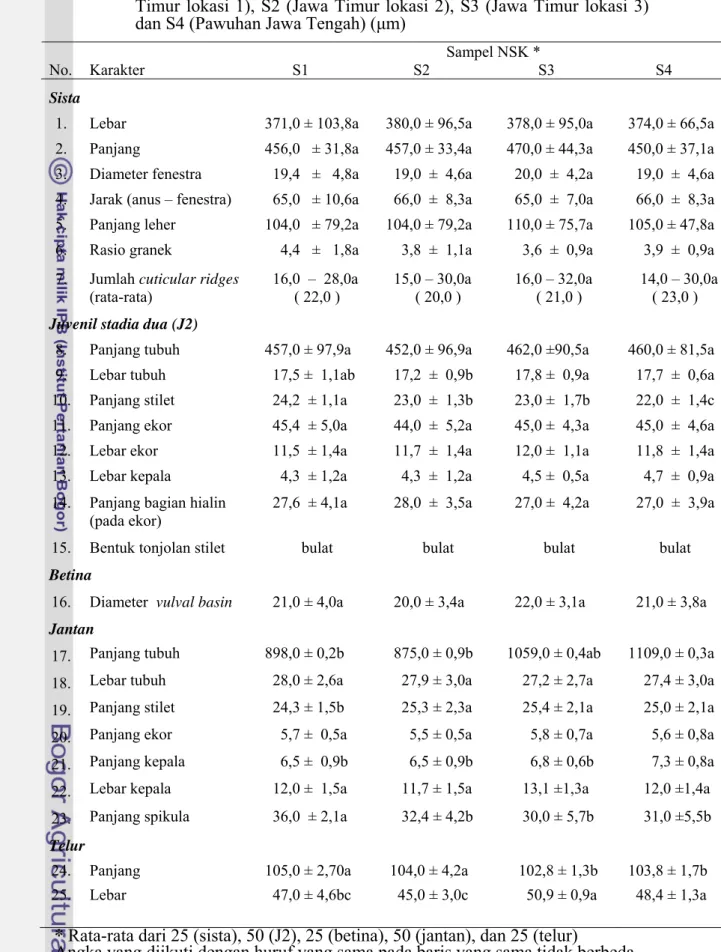
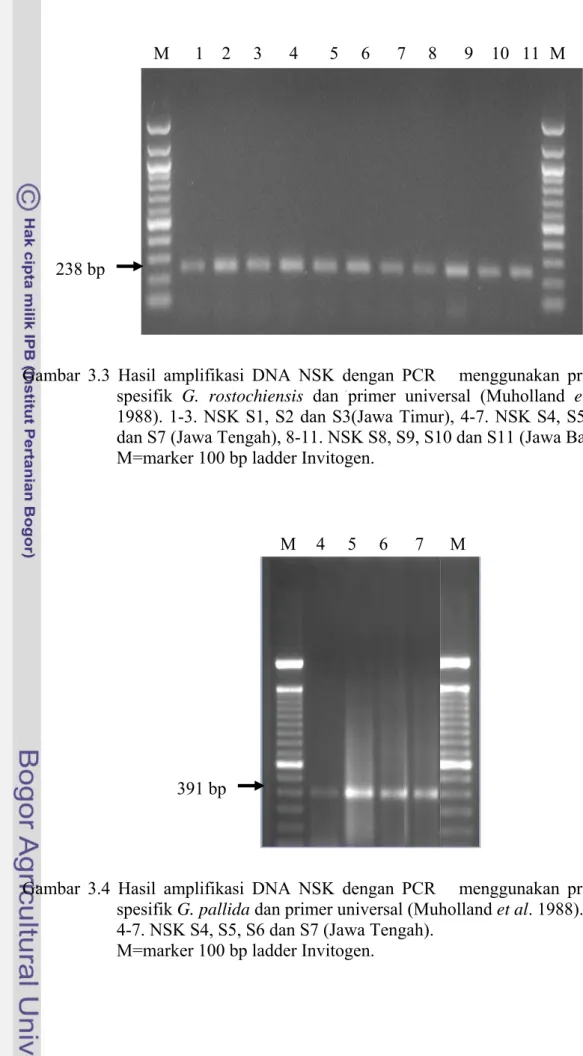
Karakter Molekuler NSK Indonesia a Daerah ITS
Hasil amplifikasi DNA dengan teknik PCR menggunakan primer spesifik dan G. pallida serta prim
NSK menghasilkan 2 pola pita, masi asing dengan ukuran 238 bp dan 391 bp. a Timur i
merupakan spesies G pita 391 bp hanya
ditemukan pada isolat Jawa Tengah (S4-S7). Pita dengan ukuran ini adalah
Perunu
kan sebagai bahan analisis tingkat kesamaan genetik NSK yang telah dikoleksi dari daerah geografi lainnya, yaitu dengan ang tersedia dalam GeneBank
(http://
Amplifikasi rDNA pad
untuk G. rostochiensis er universal terhadap 11 populasi
ng-m
Ukuran pita 238 bp ditemukan pada semua populasi NSK, yaitu dari Jaw
(S1-S3), Jawa Tengah (S4-S7) dan Jawa Barat (S8-S11). Pita dengan ukuran in
. rostochiensis (Gambar 3.3). Ukuran
spesies G. pallida (Gambar 3.4).
tan rDNA pada Daerah ITS
Perunutan DNA dilakukan terhadap fragmen DNA hasil amplifikasi PCR spesies G. rostochiensis dari isolat Jawa Timur (S1-S3) dan Jawa Tengah (S4-S7) sedangkan isolat Jawa Barat (S8-S11) belum dilakukan analisis perunutan DNA. Perunutan dilakukan dengan menggunakan mesin sekuensing ABI-Prism
3100-Avant Genetic Analyzer. Hanya data S1, S2, S3 (Jawa Timur), S6 dan S7 (Jawa
Tengah) yang selanjutnya diguna
memanfaatkan informasi perunutan DNA y
www.ncbi.nlm.nih.gov dan http://www.ebi.ac.uk) dan program Clustal-W
1,83.
Hasil analisis 5 isolat NSK dengan menggunakan program Clustal-W 1,83 menunjukkan ada beberapa bagian pada sekuen gen NSK pada daerah ITS-1 yang memiliki homologi. Isolat NSK Jawa Timur (S1, S2 dan S3) mempunyai tingkat kesamaan (similarity) yang tinggi yaitu di atas 90% tetapi berbeda jauh dengan isolat Jawa Tengah (S6 dan S7).
Gambar 3.3 Hasil amplifikasi DNA NSK dengan PCR menggunakan primer spesifik G. rostochiensis dan primer universal (Muholland et al. 1988). 1-3. NSK S1, S2 dan S3(Jawa Timur), 4-7. NSK S4, S5, S6 dan S7 (Jawa Tengah), 8-11. NSK S8, S9, S10 dan S11 (Jawa Barat). M=marker 100 bp ladder Invitogen.
Gambar 3.4 Hasil amplifikasi DNA NSK dengan PCR menggunakan primer spesifik G. pallida dan primer universal (Muholland et al. 1988). 4-7. NSK S4, S5, S6 dan S7 (Jawa Tengah).
M=marker 100 bp ladder Invitogen. 238 bp
M 1 2 3 4 5 6 7 9 8 10 11 M
M 4 5 6 7 M
1
GR1-JT1 ---TTGT 4 Gr6-JTH --- GR3-JT3 ---TTGT 4 GR2-JT2 ---TTGT 4 Gr-NY GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAGGCTGCTACTCCATGT 60 Gr-SA GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAGGCTGCTACTCCATGT 60 Gr-Meks GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr-Peru1 GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr-Kan GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr-Peru2 GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr-FI GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr-Peru3 GAGCGGTTGTTGCGCCTTGCGCAGATATGCTAACATGGAGTGTAG-CTGCTACTCCATGT 59 Gr7-JTH --- GR1-JT1 TGTACGTGCCGTACCTTGCGGCATGTTTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 64 Gr6-JTH ---GGGGCAACCCATCCCCTTTGAGATGCTACGCTCCGCAGGACGATG-AATAA 50 GR3-JT3 TGCACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 64 GR2-JT2 TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 64 Gr-NY TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 120 Gr-SA TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 120 CGGCATGTCTGCGCTTGTGTGCTACGTCCG GGCCGTGATGA 119 CGGCATGTCTGCGCTTGTGTGCTACGTCCG GGCCGTGATGA 119 r-Kan TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTATGTGCTACGTCCGTGGCCGTGATGA 119 * * * * 1-JT1 GTGGTTTAAGACT-TGA 115 Gr6-JTH GACGAC-GTGTTA---GGACCCGT--GCC-TGGAAATGGACACGAGGATTAAGACTATGA 103 R3-JT3 GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 115 R2-JT2 GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 115 r-NY GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 171 . 3 Gr-Meks TGTACGTGCCGTACCTTG T Gr-Peru1 TGTACGTGCCGTACCTTG T G Gr-Peru2 TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 119 Gr-FI TGTACGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 119 Gr-Peru3 TGTATGTGCCGTACCTTGCGGCATGTCTGCGCTTGTGTGCTACGTCCGTGGCCGTGATGA 119 Gr7-JTH ---CGGGTTTTCGGTCCTTT----GTTGCTACGTCCCGTGTGCCCGTGTTGTGC 47 * * * * * * * * GR GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CAC G G G Gr-SA GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 171 170 Gr-Meks GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA Gr-Peru1 GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 170 Gr-Kan GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 170 Gr-Peru2 GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 170 Gr-FI GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 170 Gr-Peru3 GACGAC-GTGTTA---GGACCCGT--GCC-TGGCATTGG-CACGTGGTTTAAGACT-TGA 170 Gr7-JTH GCTGACCGTGTTCTATGGACCCGTTGGCCGTGGGAACGGAGACGTGGTTTAAGACT-TGA 106 * *** ***** ******** *** *** * ** *** ** ******** *** GR1-JT1 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT 171 Gr6-JTH TGAGTGCCCGCAGGCAACCGACCAGCTTTTTCCCATTTTATATTTATTTTTTA-TGCAAT 162 GR3-JT3 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT 171 171 GR2-JT2 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT AAT 2 Gr-NY TGAGTGCCC-CAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTAATGC 27 Gr-SA TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTAATGCAAT 228 Gr-Meks TGAGTGCCGGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT 226 Gr-Peru1 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT 226 Gr-Kan TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATTTTTTA-TGCAAT 226 Gr-Peru2 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATCTT-TATTTATTTTTTA-TGCAAT 226 Gr-FI TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATATTTTA-TGCAAT 226 Gr-Peru3 TGAGTGCCCGCAGGCACC--GCCAGCTTTTTCCCATTTT-TATTTATCTTTTA-TGCAAT 226 Gr7-JTH TGAGTGGCCGGAGGAAAGAAGGGGGCC--- 133 ****** * *** * ** GR1-JT1 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 216 Gr6-JTH TCGAATGCTA-AAATATTCTAGCCTTCAACAAAAGGATCACTCGGCTC 209 GR3-JT3 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 216 GR2-JT2 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 216 Gr-NY TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 272 Gr-SA TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 273 Gr-Meks TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr-Peru1 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr-Kan TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr-Peru2 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr-FI TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr-Peru3 TCGATTGCTA-AAATATTCTAGTCTT--ATCGGTGGATCACTCGGCTC 271 Gr7-JTH
---Gambar 3.5 Perbandingan hasil perunutan nukleotida5 isolat `NSK spesies G
rostochiensis Indonesia. GR1-JT1 = S1, GR2-JT2 = S2, GR3-JT
= S3, GR6-JTH = S6, dan GR7-JTH = S7. *) Menunjukkan nukleotida yang sama.
Hasil perunutan DNA selanjutnya digunakan untuk analisis keragaman i i r ensial kdiferensial 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
Jawa Timu GeneBank
Patotipe NSK Indonesia
genetik dengan menggunakan program Clustal-W 1,83. Hasil analisis untuk nila koefisien perbedaan jarak ketidaksamaan didapat isolat-isolat Indonesia terbag dalam dua kelompok. Satu kelompok terdiri dari isolat-isolat NSK Jawa Timu (S1, S2 dan S3) dan satu kelompok lainnya terdiri dari isolat-isolat Jawa Tengah (S6 dan S7). Kedua kelompok ini terpisah dari isolat-isolat NSK asal negara lain (Gambar 3.6).
er Pengujian Patotipe dengan Klon dif
S1
S1
0 0.1 0.2
Jarak Kesamaan Genetik
r Jawa Tengah
ambar 3.6 Dendogram isolat-isolat NSK asal Indonesia terhadap NSK dari lokasi geografi lain yang ada di GeneBank (AFO168
G
73 ; AFO 16876 ; AFO 16875 ; AFO 16873 ; AFO 16877 ; AFO 16878 ; A
) berdasarkan analisis menggunakan program CFO 16874lustal-W ; AFO
16872 1,8 3 S2 S3 S6 S7 Gr-NY Gr-SA Gr-Peru1 Gr-SA Gr-Peru3 Gr-Meks Gr-Peru2 Gr-Kan
Hasil pengujian patotipe dengan menggunakan klon diferensial enunjukkan isolat 1, 2 dan 3 (S1, S2 dan S3) kemungkina kan G.
ostochiensis dengan patotipe yang spesifik Indonesia, berbeda dengan patotipe
ang ada di Eropa maupun di Amerika Selatan karena terdapat hasil patogenisitas ang berbeda pada klon diferensial AM 78.3778 (S. vernei hib. 65.346/19) pada
et al. 1977)
977), NSK tidak dihasilkan sangat rendah bila pada klon diferensial AM 78.3778 (S. vernei hib. 65.346/19)
n ini sista baru yang
erupakan e
ang ada di
Tabel 3.5 Analisis patotipe isolat S1, S2, S3 (Jawa Timur) dan S4 (Pawuhan
Jawa Tengah) pa il
m n merupa
r
y y
ke-3 isolat. Berdasarkan uji patotipe yang terdapat di Eropa (Kort maupun yang di Amerika Selatan (Canto Saenz & de Scurrah 1 dapat menghasilkan sista baru atau sista baru yang
diinokulasikan
sehingga diberi tanda negatif (–) (Tabel 3.5). Pada pengujia
terbentuk cukup tinggi maka diberi tanda positif (+). Sedangkan isolat S4
G. rostochiensis dengan patotipe Ro1 karena sesuai dengan patotip
Eropa (Tabel 3.5). m
y
da klon diferensial komers
Isolat Klon diferensial komersil Tetua S1 S2 S3 S4 Maritta S.t.tuberosum + + + + Saturna S.t.andigena CPC 1673 - - - - KTT 60.21.19 S.kurtzianum hib.60.21.19 - - - -
ODV 32731 S.vernei hib. 58.1642/4 - - - -
S.vernei hib. 62.33.3 S.vernei hib. 62.33.3 - - - -
AM 78.3778 Sin. S.vernei hib. 65.346/19 + + + -
Patotipe baru Baru baru Ro1
Pembahasan
Identifkasi spesies berdasarkan karakter morfologi didapat keempat sampel yang digunakan (S1, S2, S3 dan S4) adalah G. rostochiensis. Hasil ini
apat dilihat dari setiap sampel (S1, S2, S3 dan S4) menunjukkan rata-rata peubah
erbedaan antara satu isolat dengan isolat yang lain, tetapi umumnya karakter d
yang diamati relatif sama. Walaupun ada beberapa karakter yang mempunyai p
yang diamati berdasarkan uji jarak Duncan tidak berbeda nyata. Pengamatan pada juvenil dua (J2) dan sista merupakan kegiatan yang paling penting dalam m de i secara k i samping pengamatan pada jantan, betina dan
iller & Gra senback 1993), enggunakan daerah
an fe edakan antara spesies Globodera
virginia .
n tonjolan kutikula pada isolat S1, S2 dan
S4. Sem la p lel de an ju ah rata-rata
utikula pad S1, S2, S3 dan S4 berturut-turut sebanyak 22 ; 20 ; 21 dan asing-masing isolat adalah 4,2 ; 3,8 ; 3,6 dan 3,9. Nematoda sista kentang dengan karakter sista seperti ini merupakan NSK spesies
G. ros
um tidak terdapat perbedaan antara NSK as
eto identifikas onvensional, d
telur. M y (1972) ; Mota & Ei ( m
antara anus d nestra sista untuk memb
tabacum, G. e dan G. solanacearum
Pada pengamatan sista ditemuka
S3 maupun ua isolat mempunyai po ara ng ml
tonjolan k a
23. Rasio granek untuk m
tochiensis. Fleming & Powers (1998) menjelaskan pola tonjolan stilet
antara anus dan vulval basin dapat digunakan untuk membedakan spesies. Lebih lanjut Fleming & Powers (1998) melaporkan, G. rostochiensis mempunyai pola tonjolan kutikula yang paralel sedangkan G. pallida dan G. achilleae mempunyai pola tojolan kutikula mirip G. rostochiensis tetapi berukuran lebih besar, di samping itu G. rostochiensis mempunyai jumlah tonjolan kutikula di atas 14 dengan rasio granek lebih besar dari 3. Hasil identifikasi ini juga diperkuat dengan ciri-ciri morfologi seperti bentuk tonjolan stilet yang bulat, panjang stilet panjang tubuh J2 dan karakter lainnya, semua menunjukkan kepada G. rostochiensis.
Banyaknya persamaan yang didapat pada keempat isolat berdasarkan karakter morfologi dipertegas dengan hasil uji jarak Duncan berdasarkan wilayah (Tabel 3.4). Berdasarkan wilayah secara um
al Jawa Timur dengan Jawa Tengah. Hal ini disebabkan karena isolat Jawa Timur maupun Jawa Tengah sama-sama terindentifikasi sebagai G. rostochiensis, sehingga tidak didapatkan perbedaan yang nyata.
Identifikasi spesies dengan berdasarkan karakter molekuler memberikan hasil yang lebih akurat. Hasil amplifikasi dengan teknik PCR pada daerah ITS dengan menggunakan pasangan primer ITS-1 (5’- TGT TGT ACG TGC CGT ACC TT-3’) dengan 5,8S rDNA (5’- GCA GAA GGC TAG CGA TCT TC-3’) menghasilkan fragmen DNA berukuran 238 bp pada semua isolat-isolat Jawa Timur, Jawa Tengah dan Jawa Barat, sedangkan pasangan primer ITS-1 (5’- GGT GAC TCG ACG ATT GCT GT-3’) dengan 5,8S rDNA (5’- GCA GAA
GGC TAG CGA TCT TC-3’) menghasilkan fragmen DNA berukuran 391 bp hanya pada isolat-isolat Jawa Tengah. Menurut Muholland et al. (1996) ukuran fragmen DNA dengan pasangan primer di atas berturut-turut adalah G.
rostochiensis dan G. pallida. Hasil ini menunjukkan isolat-isolat Jawa Timur dan
Jawa B
tuk mengidentifikasi spesies
empunyai mempunyai persamaan jarak genetik yang sangat dekat yaitu d
arat merupakan populasi tunggal G. rostochiensis, sebaliknya isolat Jawa Tengah terdiri dari populasi campuran G. rostochiensis dan G. pallida (Gambar 3.3 dan 3.4).
Tingginya tingkat akurasi pada identifikasi spesies berdasarkan karakter molekuler, disebabkan pada metode ini perbedaan antara isolat dilacak pada tingkat gen, sehingga memiliki sensifitas yang tinggi un
campuran. Sebaliknya identifikasi spesies berdasarkan karakter morfologi hanya dapat mengidentifikasi spesies yang paling dominan dari rata-rata sampel yang digunakan pada pengamatan. Artinya, walaupun pada isolat S4 berdasarkan karakter molekuler merupakan spesies campuran, tetapi berdasarkan karakter morfologi rata-rata ukuran morfologinya masuk sebagai G. rostochiensis. Seperti diketahui ekspresi morfologi merupakan kombinasi antara genotipe tanaman dengan faktor lingkungan.
Analisis perunutan DNA menggunakan hasil amplifikasi PCR menunjukkan isolat-isolat Indonesia tidak berada dalam satu kelompok dengan isolat NSK dari negara lain. Isolat-isolat Indonesia terbagi dalam 2 kelompok, masing-masing kelompok pertama terdiri dari isolat Jawa Timur (S1, S2 dan S3) dan kelompok kedua terdiri dari isolat Jawa Tengah (S6 dan S7). Ketiga isolat Jawa Timur m
i atas 90%, jarak genetik seperti ini dapat dikatakan populasi NSK Jawa Timur adalah sama. Pada pengujian ini ketiga isolat tersebut memberikan reaksi yang sama pada semua klon diferensial. Tingginya tingkat persamaan runutan nukleotida ini dapat disebabkan karena umbi kentang yang ditanam berasal dari sumber yang sama sehingga ada kemungkinan nematoda sista kentang yang terinfestasi di lokasi pengambilan sampel juga mempunyai karakter genetik yang sama.
Terpisahnya isolat Jawa Timur dengan isolat Jawa Tengah menunjukkan populasi NSK di Indonesia mempunyai keragaman genetik yang tinggi. Hasil
klon diferensial, walaupun jumlah sista yang dihasilkan bervariasi pada
masing-ial sering digunakan sebagai alternatif pengujian untuk melengkapi pengam
Scurrah 1977). Patotipe pada S1, S2 dan S3 kemungkinan merupakan patotipe spesifik Indonesia, yang terjadi ka
dari Eropa dan Australia untuk waktu yang cukup lama (petani, komunikasi pribadi). Seperti diketahui Granola dan Atlantik adalah dua kultivar kentang yang yang didapat dari perunutan DNA ini sejalan dengan hasil pengujian patotipe dengan klon diferensial. Pada pengujian patotipe dengan klon diferensial didapat patotipe populasi NSK Jawa Timur berbeda dengan Jawa Tengah. Kondisi ini menunjukkan populasi NSK Indonesia mempunyai patotipe-patotipe yang berbeda. Tidak tertutup kemungkinan merupakan patotipe baru yang spesifik Indonesia.
Hasil pengamatan uji tanaman klon diferensial menunjukkan bahwa keempat isolat NSK yang digunakan dapat menghasilkan sista baru pada semua
masing inang. Matthews (1991) mendefinisikan tanaman diferensial adalah tanaman yang mampu mengekspresikan reaksi yang khas sehingga dapat membedakan patotipe antara isolat yang digunakan. Tingginya sifat spesifik pada kultivar tanaman inang tertentu untuk NSK menyebabkan pengujian klon diferens
atan yang berdasarkan karakter morfologi.
Pada Tabel 3.5 terlihat isolat 4 (S4) memberikan hasil yang sama seperti skema yang didiskripsikan oleh Canto Saenz & de Scurrah (1977) ; Kort et al. (1977) (Tabel 3.2) untuk G. rostochiensis dengan patotipe Ro1. Sebaliknya pada S1, S2 dan S3 memberikan hasil yang berbeda pada klon AM 78.3778 (S. vernei hib. 65.346/19). Hasil ini mengindikaskan S1, S2 dan S3 bukan seperti patotipe yang terdapatdi Eropa (Kort et al. 1977) maupun yang di Amerika Selatan (Canto Saenz & de
rena adanya mutasi pada isolat-isolat tersebut.
Dugaan kuat telah terjadi variasi genetik pada isolat-isolat Jawa Timur adalah berhubungan erat dengan kultivar kentang yang diimpor dan negara asal impor. Sebelum adanya laporan keberadaan NSK pada Maret 2003 yang lalu (Daryanto 2003), petani daerah ini telah menggunakan bibit umbi kentang impor
paling banyak diimpor dan ditanam di Indonesia. Granola diimpor dari negara Eropa. Sedangkan kentang kultivar Atlantik diimpor dari Australia. Impor bibit kentang
sis Ro1. Hasil penelitian menunjukkan Granola dan Atlantik dapat
diseran
dalam jumlah yang cukup besar untuk waktu yang cukup lama tanpa prosedur karantina, memberi peluang besar masuknya NSK ke daerah ini. Mengingat Jawa Timur merupakan salah satu sentra pertanaman kentang di Indonesia, maka penanaman kentang pasti dilakukan dalam skala yang luas secara terus menerus sepanjang tahun. Hal ini merupakan kondisi yang sangat mendukung untuk terjadinya ledakan dan terbentuknya patotipe baru di daerah ini. Menurut Joosten (1991), Granola merupakan kultivar kentang yang resisten terhadap G. rostochiensis Ro1 dan Ro4, sedangkan Atlantik resisten terhadap G.
rostochien
g oleh NSK, bahkan di beberapa lokasi pertanaman kentang di Kota Batu, Jawa Timur tanaman kentang mati pada umur tanaman kurang dari satu bulan. Kondisi ini memperkuat dugaan NSK yang terdapat di Jawa Timur merupakan patotipe baru. Agrios (2005) mengemukan ada 3 faktor yang harus dimiliki oleh patogen untuk dapat menginfeksi tanaman yaitu 1) Kemampuan penetrasi dan menginfeksi tanaman, 2) Kemampuan untuk mengatasi ketahanan tanaman, 3) Kemampuan patogenisitas. Lebih lanjut dilaporkan salah satu aspek biologis yang sangat dinamis dan sangat menentukan adalah karakter individu dalam satu spesies yang selalu mengalami perubahan baik morfologis maupun fisiologisnya, kemampuan berubahnya bervariasi dari satu individu ke individu yang lain sebagai suatu kemampuan beradaptasi. Sinaga (2003) menyatakan bahwa perubahan-perubahan yang terjadi pada patogen, dapat meningkatkan sifat patogenitasnya, sehingga suatu saat dapat membentuk suatu strain baru yang virulen dan dapat menyerang inang yang resisten.
Selain alasan di atas, dugaan kuat terdapatnya patotipe baru spesifik Indonesia adalah adanya adaptasi lingkungan karena perbedaan temperatur tanah dan panjang hari, seperti yang terjadi pada negara Eropa. NSK merupakan patogen yang berasal dari Kepulauan Andes (Amerika Selatan), yang merupakan daerah berhari pendek yaitu panjang siang adalah 12 jam. Introduksi NSK melalui