17
PERAN FGF21 PADA PROSES PENCOKELATAN
JARINGAN ADIPOSA
THE ROLE OF FGF21 IN THE ADIPOSA TISSUE BROWNING PROCESS Nisa Kartika Komara
Program Studi D IV-Teknik Laboratorium Medik, Politeknik Kesehatan Hermina, Jakarta Timur, Indonesia E--mail: [email protected]
Diterima : 2 Maret 2021 Direvisi : 12 April 2021 Disetujui : 29 April 2021 Abstrak
Konsumsi makanan yang berlebihan tanpa disertai dengan aktivitas fisik akan menyebabkan akumulasi lemak di dalam jaringan adiposa. Jaringan adiposa yang berperan dalam menyimpan kelebihan energi adalah jaringan adiposa putih. Salah satu mekanisme untuk mengurangi kadar profil lemak tubuh yang berlebihan dalam jaringan adiposa putih adalah melalui mekanisme pencokelatan. Mekanisme pecokelatan ditandai dengan adanya peningkatan ekspresi gen Uncoupling Protein 1 (UCP1). Aktivasi UCP1 terjadi melalui beberapa persinyalan yang mengaktivasi beberapa protein transkripsi, salah satunya adalah PPAR gamma
coactivator 1α (PGC-1α). PPAR gamma coactivator 1α (PGC-1α) distimulasi oleh protein sirtuin 1 (SIRT1)
yang diaktivasi Fibroblast Growth Factor 21 (FGF21) melalui ikatannya dengan kompleks reseptor FGFR1/β-Klotho. Fibroblast Growth Factor 21 (FGF21) memiliki peran penting dalam menjaga keseimbangan energi dengan cara meningkatkan proses termogenesis.
Kata kunci: β-Klotho, FGF21, FGFR1, Jaringan adiposa, Pencokelatan, UCP1
Abstract
Excessive food consumption without physical activity causes the accumulation of fat in adipose tissue. Adipose tissue plays a role in storing the excess energy is white adipose tissue. A mechanism to reduce levels of excess body fat profiles in white adipose tissue is through the mechanism of browning. The browning mechanism is characterized by an increase in the expression of the Uncoupling Protein 1 (UCP1) gene. UCP1 activation occurs through multiple signaling and some would ak transcription proteins, one of which is a PPAR gamma co-activator 1α (PGC-1α). The gamma coactivator PPAR 1α (PGC-1α) is stimulated by the protein sirtuin 1 (SIRT1) which is activated by Fibroblast Growth Factor 21 (FGF21) by binding to the FGFR1 / β-Klotho receptor complex. Fibroblast Growth Factor 21 (FGF21) has an important role in maintaining energy balance by increasing thermogenesis process.
Keywords: β-Klotho, FGF21, FGFR1, Adipose tissue, Browning, UCP1
Pendahuluan
Ketidakseimbangan antara energi yang masuk dan keluar tubuh serta kurangnya aktivitas fisik akan menimbulkan akumulasi lemak di dalam jaringan adiposa putih. Akumulasi lemak yang berlebihan dapat berekembang menjadi obesitas yang dapat menimbulkan gangguan metabolik, diantaranya diabetes melitus 2, penyakit kardiovaskular,
kelainan pada saraf, dan kanker.1 Beberapa penelitian sudah banyak dilakukan untuk
mengurangi jumlah lemak yang berelebihan di dalam jaringan adiposa. Menurut Castillo et al. (2017) diferensiasi jaringan adiposa putih menjadi jaringan adiposa beige dapat
menurunkan berat badan pada penderita obesitas.2 Penelitian Neyrinck et al. (2017) juga
menunjukkan bahwa diferensiasi jaringan adiposa putih yang diinduksi oleh ekstrak daun
18
hasil penelitian tersebut, mekanisme diferensiasi jaringan adiposa putih menjadi beige dapat
dijadikan target pengobatan untuk obesitas.1
Diferensiasi jaringan adiposa putih menjadi jaringan adiposa beige disebut dengan pencokelatan atau browning. Jaringan adiposa beige memiliki karateristik yang sama
dengan jaringan adiposa coklat, sehingga dapat berperan dalam proses thermogenesis.4
Mekanisme pencokelatan dapat distimulasi oleh beberapa faktor, seperti paparan dingin,
exercise, dan environmental enrichment.5 Masing-masing kondisi tersebut kemudian akan
menginduksi pencokelatan melalui jalur persinyalan yang mengaktivasi faktor transkripsi utama dalam pencokelatan, yaitu PPARγ, PRDM16, dan PGC1α. Aktivasi dari faktor-faktor transkripsi tersebut kemudian akan meningkatkan ekspresi dari UCP1 yang merupakan gen
yang bertanggung jawab dalam proses pencokelatan.6
Peningkatan ekspresi UCP1 dipengaruhi oleh beberapa protein serum yang dapat menginduksi aktivitas dari faktor transkripsi pada pencokelatan, diantaranya, Fibroblast
Growth Factor 21 (FGF21).6 Menurut Kim et al. (2017) peningkatan ekspresi UCP1 dalam
proses pencokelatan melalui aktivasi PGC1α diinduksi oleh peningkatan sekresi FGF21 oleh
jaringan adiposa coklat.5 Fibroblast Growth Factor 21 (FGF21) dapat diekspresikan juga di
hati dan berperan dalam menjaga keseimbangan energi di dalam tubuh.4 Peningkatan
FGF21 secara farmakologis telah terbukti dapat meningkatkan proses pencokelatan,
sehingga dapat menurunkan berat badan pada tikus yang mengalami obesitas.7 Selain itu,
penelitian lain juga menunjukkan bahwa tikus obese knockout FGF21 mengalami kanker
hati, karena akumulasi lemak di dalam hati yang berlebihan.8 Peran FGF21 dalam
pengaturan energi di dalam tubuh dapat terjadi apabila terbentuk ikatan kompleks dengan
reseptornya, yaitu FGFR1c dan β-Klotho.9
Ikatan antara FGF21 dan kompleks FGFR1c/β- Klotho akan membentuk transduksi sinyal yang meningkatkan aktivitas Sirtuin-1 (SIRT1), sehingga menstimulasi PGC1α untuk meningkatkan ekspresi UCP1. Ikatan yang terjadi antara FGF21 dan reseptornya sangat penting untuk menjalankan peran FGF21 dalam menginsiasi terjadinya pencokelatan. Penelitian yang dilakukan pada tikus obesitas menunjukkan bahwa ikatan antara FGF21
dan reseptornya tidak terbentuk, terlihat dari menurunnya ekspresi UCP1.10
Tipe Jaringan Adiposa
Jaringan adiposa merupakan organ utama yang menyimpan kelebihan energi dalam bentuk triagliserol. Jaringan tersebut terdiri atas sel lemak atau adiposit dan stroma-vascular
fraction. Jaringan adiposa berperan untuk menyimpan cadangan lemak, sehingga berlebihnya
lemak yang masuk dalam tubuh akan menyebabkan pembesaran struktur pada jaringan adiposa. Jaringan adiposa terdiri atas jaringan adiposa putih, jaringan adiposa coklat, dan
19
Gambar 1. Jenis jaringan adiposa13
Klasifikasi Jaringan Adiposa
Jaringan adiposa putih atau white adipose tissue merupakan jaringan adiposa yang banyak ditemukan di jaringan subkutan. Jaringan tersebut memiliki struktur yang tersusun atas sel-sel lemak droplet unilokuler, sedikit mitokondria, dan memiliki tingkat oksidatif yang rendah. Jaringan adiposa putih berasal dari sel-sel mesenkimal dan sudah mengalami perkembangan pada saat di rahim, namun perkembangan tersebut akan semakin meningkat saat lahir. Perkembangan jaringan adiposa putih saat lahir menjadi meningkat dikarenakan adanya respon dari tubuh untuk mengatur keseimbangan energi dalam tubuh, sehingga ukuran dari jaringan adiposa putih akan terus mengalami perkembangan sepanjang masa hidup organisme. Oleh karena itu, jaringan adiposa putih memiliki peran dalam penyimpanan
energi dan disimpan dalam bentuk triagliserol (TAG). 4
Jaringan adiposa coklat memiliki ukuran yang lebih kecil daripada jaringan adiposa putih dan tersususun atas sel-sel lemak multilokular, namun memiliki sitoplasma yang relatif lebih banyak. Selain itu, jaringan adiposa coklat membutuhkan oksigen yang lebih besar, sehingga mengandung lebih banyak pembuluh kapiler daripada jaringan adiposa putih. Jaringan adiposa cokelat merupakan jaringan lemak yang berwarna cokelat hingga sedikit kemerahan yang disebabkan oleh banyakanya pembuluh darah dan mitokondria. Mitokondria tersebut
memiliki uncoupling protein-1 (UCP1) yang berperan dalam proses pengeluaran energi.5
Jaringan adiposa coklat terbentuk melalui sel yang sama dengan jaringan adiposa putih, yaitu berasal dari sel-sel mesenkimal. Namun, dalam perkembangannya jaringan adiposa cokelat lebih banyak pada bayi yang baru lahir dan akan berkurang seiring dengan bertambahnya usia. Jumlah jaringan adiposa coklat yang banyak pada bayi yang baru lahir berperan untuk adaptasi terhadap perubahan suhu lingkungan, yaitu saat terkena paparan
dingin.4 Pada saat dewasa jaringan adiposa putih yang terbentuk sudah semakin banyak,
akumulasi jaringan lemak tersebut yang akan menurunkan aktivitas dari jaringan adiposa coklat. Hal tersebut dapat dilihat dari jumlah jaringan adiposa coklat pada penderita
obesitas yang sangat sedikit sekali jika dibandingkan dengan berat badan normal.14 Ilustrasi
komposisi jaringan adiposa coklat dapat dilihat pada gambar 2.
20
Gambar 2. Jaringan adiposa coklat berdasarkan usia14
Pada orang dewasa kemampuan termogenesis jaringan adiposa coklat dapat dilakukan oleh jaringan adiposa putih yang dapat berdifernsiasi. Proses tersebut terjadi ketika ada paparan dingin, maka jaringan adiposa putih akan berdiferensiasi menjadi jaringan adiposa beige. Jaringan adiposa beige memiliki struktur seperti dengan jaringan adiposa cokelat, yaitu memiliki droplet lipid multilokuler, jumlah mitokondria banyak, dan tingginya ekspresi gen
yang spesifik pada lemak cokelat seperti UCP1.15 Pembentukan jaringan adiposa beige juga
berasal dari sel-sel mesenkimal yang sama dengan jaringan adiposa coklat, namun yang
membedakan adalah sel prekusornya.4
Mekanisme Pembentukan Jaringan Adiposa
Pembentukan jaringan adiposa putih, beige, dan coklat berawal dari sel induk mesenkimal yang sama. Perbedaan yang terjadi dalam proses pembentukan jaringan adiposa terdapat pada sel prekusornya. Pada awal perkembangan embrio jaringan adiposa yang paling banyak dibentuk adalah jaringan adiposa coklat. Jaringan adiposa coklat berasal dari sel prekursor
dalam mesoderm embrionik yaitu sel Myf5+ dan Pax7. Beberapa anggota protein yang
berperan dalam pembentukan tulang, seperti BMP4, BMP8b dan BMP7 terlibat dalam perubahan pre adiposit jaringan adiposa coklat menjadi adiposit coklat. Beberapa faktor transkripsi, seperti PRDM16, PPARγ, dan PGC1α juga berperan dalam diferensiasi jaringan adiposa coklat. PRDM16 akan menstimulasi diferensiasi jaringan adiposa coklat melalui pengikatannya terhadap PGC1α dan PPARγ yang kemudian akan mengaktifkan gen
pencokelatan, yaitu UCP1. 4,16
Jaringan adiposa putih dan beige berasal dari sel prekusor yang sama yaitu Myf5-. Proses
pembentukannya juga dibantu oleh beberapa protein, seperti BMP2 dan BMP4. Pembentukan jaringan adiposa putih kemudian terjadi melalui pengikatan faktor transkripsi PPARγ dengan c- AMP-response element-binding protein (CREBP), sehingga mengaktifkan transkripsi gen dari jaringan adiposa putih. Diferensiasi dari jaringan adiposa putih akan membentuk jaringan adiposa yang mirip dengan adiposa cokelat, yaitu adiposa beige. Jaringan adiposa beige dapat terbentuk melalui deasetilasi PPARγ oleh SIRT1 yang diperlukan untuk menstabilkan dan mengaktivasi PRDM16 untuk selanjutnya berinteraksi
dengan PGC1α, sehingga mengaktifkan gen pencokelatan. 4,5,14
21
diekspresikan baik dalam jaringan adiposa coklat maupun adiposa beige. MicroRNA yang dapat PRDM16, yaitu miR-193b-365 yang dibutuhkan untuk adipogenesis jaringan adiposa coklat. Sebaliknya, miR-133a akan menekan ekspresi PRDM16 dan menghambat diferensiasi jaringan adiposa coklat. Inaktivasi miR-133a tidak akan berefek pada perkembangan jaringan adiposa cokelat, namun meningkatkan perkembangan dari adiposa
beige pada jaringan adiposa putih. Beberapa microRNA lainnya yang terlibat dalam proses
pembentukan jaringan adiposa adalah miR-155, miR-196a, dan miR26a/26b.18
Mekanisme Pencokelatan
Mekanisme pencokelatan merupakan mekanisme yang menyebabkan adanya perubahan
struktur dari jaringan adiposa putih menjadi jaringan adiposa beige.4 Proses pencokelatan
dapat muncul setelah mendapatkan beberapa stimulus, seperti paparan dingin, latihan fisik,
dan enviromental enrichment. 5 Mekanisme pecokelatan ditandai dengan adanya
peningkatan ekspresi gen Uncoupling Protein 1 (UCP1) yang berperan dalam fosforilasi ATP. UCP1 dapat mentranslokasi proton melewati membran dalam mitokondria jaringan adiposa cokelat, melalui proses tersebut UCP1 melepaskan energi dari sintesis ATP dan kemudian menyebabkan pelepasan energi dalam bentuk panas, dan juga menstimulasi
oksidasi asam lemak. 13
Gambar 3. Jalur yang memengaruhi proses pencokelatan11
Ekspresi UCP1 dapat terjadi melalui beberapa jalur yang diregulasi oleh faktor-faktor transkripsi utama, seperti, PPAR γ, PGC-1α, dan PRDM 16. PPARγ merupakan faktor transkripsi yang penting dalam diferensiasi jaringan adiposa cokelat dan putih. PPARγ dibutuhkan untuk rekrutmen PRDM 16 ke kompleks transkripsi PPARγ, yang akan membuat proses pencokelatan. Regulator lain yang dapat menginduksi pencokelatan adalah PGC-1α yang merupakan koaktivator transkripsi yang terlibat dalam mengontrol
metabolisme energi yang banyak diekspresikan di jaringan adiposa cokelat.6 Ekspresi
PGC-1α yang meningkat dalam respon metabolik dapat terjadi karena adanya stimulasi dapat disebabkan oleh Beta 3-adenergik. Peningkatan ekspresi tersebut dapat terjadi melalui fosforilasi PGC-1α yang distimulasi oleh activating transcription factor 2 (ATF2) yang diaktifkan oleh protein kinase teraktivasi mitogen p38. Hal tersebut menyebabkan PGC-1α akan menginduksi biogensis mitokondria dan aktivasi faktor transkripsi PPAR, sehingga
22
Regulator FGF21
Fibroblast Growth Factor 21 (FGF21) merupakan salah satu protein dari anggota fibroblast
growth factors (FGFs) yang terletak pada kromosom 19 dengan berat molekul sekitar 17
sampai 34 kDa. FGF21 diekspresikan oleh beberapa organ, seperti hati, adiposa putih,
pankreas, otak dan otot rangka yang dapat dilihat di gambar 2.2.2(1).12 Hati dan jaringan
adiposa merupakan organ yang menjadi tempat kerja utama untuk FGF21.
Mekanisme kerja FGF21 pada organ-organ tersebut dapat berupa parakrin, autokrin, dan endokrin. Hal tersebut menyebabkan FGF21 dapat mengontrol homeostasis energi dalam
tubuh, sehingga dapat meningkatkan pengeluaran energi pada penderita obesitas.19
Fibroblast Growth Factor 21 (FGF21) dapat melakukan perannya melalui pembentukan
sinyal. Aktivitas transduksi sinyal dari FGF21 bergantung pada ikatannya dengan reseptor
FGF (FGFR) dan koreseptor yang disebut β-Klotho (KLB).20
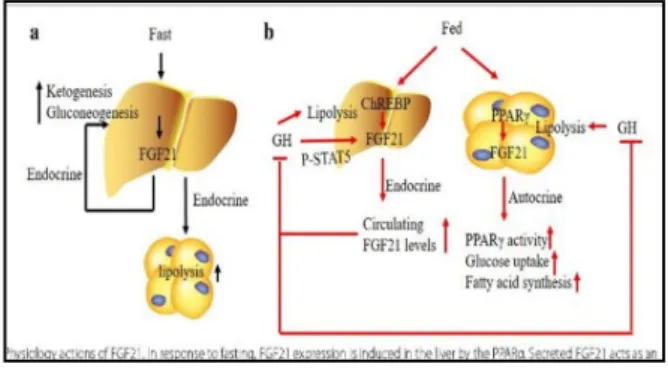
Hati merupakan organ terbesar yang memroduksi dan meregulasi FGF21. Ekspresi FGF21 pada hati dan plasma akan meningkat ketika puasa dan diet ketogenik. Ekspresi FGF21 pada hati saat puasa dimediasi oleh faktor transkripsi proliferator peroxisome alpha receptor (PPARα). FGF21 yang disekresikan oleh hati bekerja sebagai faktor endokrin. Faktor transkripsi tersebut diaktifkan oleh asam lemak yang dikeluarkan oleh adiposit selama puasa ataupun diet ketogenik. Selain itu, FGF21 juga dapat meregulasi faktor transkripsi lain, yaitu PPAR gamma coactivator 1α (PGC-1α) yang dapat meregulasi oksidasi asam lemak melalui peningkatan fungsi mitokondria dan biogenesis. Ekspresi FGF21 juga dapat ditemukan saat tidak puasa, namun faktor transkripsi yang meginduksinya berebeda dengan saat puasa. Faktor transkripsi tersebut adalah proliferator peroxisome alpha receptor (PPARγ) yang terdapat pada jaringan adiposa putih dan ChREBP pada hati. FGF21 yang disekresikan oleh hepatosit dapat bekerja secara endokrin, parakrin, ataupun autokrin yang memodulasi ketogenesis, glukoneogenesis, oksidasi asam lemak, hormon pertumbuhan, dan
karsinogenesis.15,19,21
Gambar 4. Regulasi FGF21 pada hati21
Organ penting lain yang berperan dalam sekresi FGF21 adalah jaringan adiposa. FGF21 pada jaringan adiposa berperan dalam pemgambilan glukosa, lipolisis, dan meningkatkan kapasitas oksidatif mitokondria. Jaringan adiposa dapat dibedakan menjadi 3 struktur, yaitu jaringan adiposa putih, jaringan adiposa krem atau beige, dan jaringan adiposa cokelat. Ekspresi dan sekresi jaringan adiposa putih diregulasi oleh proliferator peroxisome alpha
23
receptor (PPARγ). 15,19 Pada jaringan adiposa putih FGF21 dapat bekerja secara autokrin
untuk menstimulasi PPARγ dan pengambilan glukosa, sedangkan secara endokrin dalam
menekan aktivitas lipolisis.21 FGF21 di jaringan adiposa putih juga dapat bereperan dalam
homeostasis energi, terutama dalam proses thermogenesis. FGF21 akan meregulasi interskapular dari jaringan adiposa cokelat untuk meningkatkan aktivitas termogenesis
melalui induksi protein UCP1.19
Jaringan adiposa yang menjadi lokasi untuk proses termogenesis adalah jaringan adiposa cokelat. Ekspresi FGF21 pada proses termogenesis distimulasi oleh paparan dingin yang akan merangsang pelepasan norepinefrin. Norepinefrin kemudian akan berikatan dengan reseptor adenergik untuk meningkatkan cAMP. Peningkatan cAMP akan mengaktifkan transkripsi gen FGF21 melalui elemen pengikat ATF2. Pelepasan neurotransmiter tersebut akan meningkatkan aliran darah, sehingga mengaktifkan enzim lipase untuk memecahkan
trigliserida menjadi asam lemak dan gliserol.19
Penelitian mengenai peran FGF21 pada jaringan adiposa telah banyak dilakukan, salah satunya adalah penelitian yang menggunakan adiposit 3T3- L1. Pada penelitian tersebut ditemukan bahwa pemberian FGF21 secara kronis akan meningkatkan ekspresi PPARγ. Selain itu, pemberian FGF21 pada adiposit tersebut akan meningkatkan ekspresi mRNA dan protein Glut-1 yang berhubungan dengan peningkatan pengambilan glukosa ke dalam sel sel adiposa. FGF21 akan menstimulasi p44/42, sehingga mengaktifkan fosforilasi dan faktor transkripsi serum respon factor (SRF) serta Ets- like protein-1 (Elk-1) pada adiposit 3T3-L1. Hal tersebut yang menyebabkan peningkatan pengambilan glukosa pada jarigan adiposa putih. Pada penderita obesitas ekspresi GLUT-1 pada jaringan adiposa akan menurun,
sehingga terjadi resistensi terhadap FGF21.15
Peran FGF21 Pada Proses Pecokelatan
Fibroblast growth factor 21 (FGF21) merupakan faktor endokrin yang memiliki peran
dalam menurunkan kadar glukosa, insulin, trigliserida, dan berat badan pada penderita
obesitas.22 Potensi FGF21 dalam menjalankan peran tersebut harus diawali dengan
pembentukan sinyal. FGF21 akan membentuk sinyal apabila berikatan dengan reseptor. Reseptor FGF (FGFRs) merupakan reseptor permukaan sel yang termasuk ke dalam reseptor
tirosin kinase. FGF21 dapat berikatan dengan beberapa jenis FGFRs, diantaranya
FGFR1c, -2c, -3c, dan -4). Ikatan FGF21 dan FGFRs dengan afinitas yang paling tinggi terdapat pada FGFR1c. FGF21 yang sudah berikatan dapat membentuk trasnduksi sinyal, sehingga dibutuhkan koreseptor.
24
Koreseptor yang berperan penting dalam persinyalan FGF21 adalah β-Klotho (KLB). Koreseptor β- Klotho merupakan protein transmembran tunggal yang akan membentuk
kompleks dengan FGFR1c dan FGF21.19 Fibroblast growth factor 21 memiliki dua sisi
pengikatan dalam membentuk ikatan dengan FGFR1c dan β-Klotho yang terletak di C dan N terminus. Ujung C FGF21 akan berinterkasi dengan β-Klotho dan ujung N akan berinteraksi dengan domain imunoglobulin D2 dan D3 dari reseptor FGFR1c. Ikatan kompleks antara FGF21 dengan reseptornya akan memicu terbentuknya transduksi sinyal untuk memulai
proses pencokletan pada jaringan adiposa.20
Gambar 5. Kompleks FGF21 dan FGFR1c/ β-Klotho20
Proses pencokelatan dapat meningkatkan pembakaran energi secara alami, sehingga
berpotensi untuk mengurangi profil lemak dalam tubuh.4 Ekspresi FGF21 pada proses
termogenesis, salah satunya distimulasi oleh paparan dingin yang akan merangsang pelepasan norepinefrin menstimulasi reseptor β-adenergik pada jaringan adiposa cokelat. Norepinefrin kemudian akan berikatan dengan reseptor adenergik untuk meningkatkan cAMP. Peningkatan cAMP akan mengaktifkan protein kinase A (PKA) yang akan memfosforilasi p38 MAPK, kemudian mengaktifkan transkripsi gen FGF21 melalui elemen pengikat ATF2 protein kinase A (PKA). Selain itu, fosforilasi p38 MAPK juga dapat menginduksi transkripsi gen yang merupakan mediator untuk gen thermognesis, seperti UCP1. Aktivasi PKA juga akan menginduksi pemecahan lipase atau lipolisis. Selain itu aktivasi p38 MAPK juga dipengaruhi oleh miR-32 yang berperan dalam menghambat
Tob1. Mekanisme tersebut dapat dilihat pada gambar 2.4. (2). Selanjutnya FGF21 yang
disekresikan akan bekerja secara endokrin maupun autokrin.23,24 FGF21 akan mengaktifkan
protein transkripsi, yaitu PPAR gamma coactivator 1α (PGC-1α). Peningkatan aktivitas PGC-1α distimulasi oleh SIRT1 yang diaktivasi oleh FGF21 yang berikatan secara kompleks dengan reseptornya. Proses tersebut akan menginduksi ekspresi UCP1 pada
25
Gambar 6. Mekanisme sekresi FGF21 pada pencokelatan24
Kesimpulan
Akumulasi lemak yang berlebihan akan tersimpan di dalam jaringan adiposa putih. Lemak yang berlebihan dalam jaringan adiposa dapat dikurangi melalui mekanisme pencokelatan. Mekanisme pencokelatan ditandai dengan adanya peningkatan ekspresi gen
Uncoupling Protein 1 (UCP1). Aktivasi UCP1 terjadi melalui beberapa persinyalan yang
mengaktivasi beberapa protein transkripsi, diantaranya PPAR gamma coactivator 1α (PGC-1α). Proses pencokelatan dapat diinduksi oleh FGF21 yang akan berikatan secara kompleks dengan FGFR1c/β-Klotho. Ikatan tersebut kemudian akan menghasilkan trasnduksi sinyal yang akan meningkatkan aktivitas PGC-1α distimulasi oleh SIRT1, sehingga mengaktivkan gen UCP1 untuk selanjutnya menginduksi proses diferensiasi jaringan adiposa putih menjadi beige.
DAFTAR PUSTAKA
1. Straub L, Wolfrum C. FGF21 energy expenditure and weight loss-How much brown fat do you need. MolMetab. 2015;4(9):605–9.DOI:10.1016/j.molmet.2015.06.008
2. Vargas-castillo A, Fuentes-romero R, Rodriguez- lopez LA, Torres N, Tovar AR. Understanding the Biology of Thermogenic Fat : Is Browning A New Approach to the Treatment of Obesity?. Arch Med Res.2017:1–13.DOI: 10.1016/j.arcmed.2017.10.002
3. Neyrinck AM, Bindels LB, Geurts L, Van M, Cani PD, Delzenne NM. J Nutr Biochem. 2017:1–23. DOI: 10.1016/j.jnutbio.2017.07.008
4. Giralt M, Villarroya F. White,Brown,Beige/Brite: Different Adipose Cells for Different Functions?. Endocrinol. 2018;15:2992–3000. DOI:10.1210/en. 2013- 140
5. Kim SH, Plutzky J. Brown Fat and Browning for the Treatment of Obesity and Related. Diabetes Metab J. 2016;40:12–21.
6. Lo KA, Sun L. Turning WAT into BAT : a review on regulators controlling the browning of white adipocytes. Biosci Rep. 2013:711–9. DOI: 10.1042/BSR20130046
7. Kharitonenkov A, Adams AC. Inventing new medicines : The FGF21 story. Mol Metab. 2014;3:221–9.
8. Singhal G, Kumar G, Chan S, Fisher M, Ma Y, Vardeh HG, et al. Deficiency of fibroblast growth factor 21 (FGF21) promotes hepatocellular carcinoma (HCC) in mice on a long term obesogenic diet.MolMetab.2018;13:5666.DOI:10.1016/j.molm
26 et.2018.03 .002
9. Kilkenny DM, Rocheleau J V. The FGF21 Receptor Signaling Complex : Klotho β,FGFR1c,and Other Regulatory Interactions. 1st ed. Elsevier Inc, 2016. 17-58 p. DOI: 10.1016/bs.vh.2016.02.008
10. Fu T, Seok S, Choi S, Huang Z, Suino-powell K, Xu HE, et al. MicroRNA 34a Inhibits Beige and Brown Fat Formation in Obesity in Part by Suppressing Adipocyte Fibroblast Growth Factor 21 Signaling and SIRT1 Function. Mol Cell Biol. 2014;34(22):4130–42.
11. Cristina T, Bargut L, Souza-mello V, Aguila MB, Mandarim-de-lacerda CA. Browning of white adipose tissue : lessons from experimental models. Horm Mol Biol Clin Invest. 2017:1– 13. DOI: 10.1515/hmbci-2016-0051
12. Kim KH, Lee M. FGF21 as a mediator of adaptive responses to stress and metabolic benefits of antidiabetic drugs. J Endrocinol. 2015;226: R1–R16. DOI: 10.1530/JOE-15-0160
13. Nedergaard J, Cannon B. Brown Adipose Tissue: Development and Function. 5th ed. Elsevier Inc, 354-363p. DOI: 10.1016/B978-0-323-35214-7.00035-4
14. Saito M. Brown Adipose Tissue as a Regulator of Energy Expenditure and Body Fat in Humans. Diabetes Metab J. 2013;22–9.
15. Ge X, Wang Y, Sl K, Xu A. Metabolic actions of FGF21 : molecular mechanisms and therapeutic implications. Acta Pharm Sin B. 2012;2(4):350– 7.DOI:10.1016/j.apsb.2012.06 011
16. Becerril S, Ezquerro S, M L. Crosstalk between adipokines and myokines in fat browning. Act Physiol. 2016;1–20. DOI: 10.1111/apha.12686
17. Reddy NL, Tan BK, Barber TM, Randeva HS. Brown adipose tissue : endocrine determinants of function and therapeutic manipulation as a novel treatment strategy for obesity. BMC Obesity. 2014;1:1–13.
18. Lee SB. Intricate transcriptional networks of classical brown and beige fat cells. Front endocrinol. 2015;6:1–7. DOI: 10.3389/fendo.2015.00124
19. Fisher FM, Maratos-flier E. Understanding the Physiology of FGF21. Annu Rev Physiol. 2016;78:223–41. DOI: 10.1146/annurev-physiol-021115-105339
20. Davis D, Garg C, Lahiri SS. Therapeutic potential of FGF21 in diabetes. JMPS. 2017;5(3):364– 72.
21. Guan D, Zhao L, Chen D, Yu B, Yu J. Regulation of fibroblast growth factor 15/19 and 21 on metabolism : in the fed or fasted state. J Transl Med. 2016;1–7.
22. Yie J, Wang W, Deng L, Tam L, Stevens J, Chen M, et al. Understanding the Physical Interactions in the FGF21/FGFR/β-Klotho Complex : Structural Requirements and Implications in FGF21 Signaling. Chem Biol Drug Des. 2012;398–410.
23. Hondares E, Iglesias R, Giralt A, Gonzalez FJ, Giralt M, Mampel T, et al. Thermogenic Activation Induces FGF21 Expression and Release in Brown Adipose Tissue. J Biol Chem. 2011;286(15):12983– 90.
24. Ng R, Hussain NA, Zhang Q, Chang C, Li H, Fu Y, et al. miRNA-32 Drives Brown Fat Thermogenesis and Trans-activates Subcutaneous White Fat Browning in Mice Article miRNA-32 Drives Brown Fat Thermogenesis and Trans-activates Subcutaneous White Fat Browning in Mice. Cell Reports.2017;19(6):1229–46.DOI:10.1016/j.celrep. 2017.04.035