Ž .
Chemical Geology 180 2001 99–115
www.elsevier.comrlocaterchemgeo
The effect of Fe-oxidizing bacteria on Fe-silicate
mineral dissolution
Cara M. Santelli
a,1, Susan A. Welch
a,), Henry R. Westrich
b, Jillian F. Banfield
aa
Department of Geology and Geophysics, UniÕersity of Wisconsin-Madison, 1215 W. Dayton St., Madison WI 53706 1692, USA
b
Geochemistry Department, Sandia National Laboratories, Albuquerque NM 87185-0750, USA
Abstract
Ž .
Acidithiobacillus ferrooxidans are commonly present in acid mine drainage AMD . A. ferrooxidans derive metabolic
energy from oxidation of Fe2qpresent in natural acid solutions and also may be able to utilize Fe2qreleased by dissolution
of silicate minerals during acid neutralization reactions. Natural and synthetic fayalites were reacted in solutions with initial pH values of 2.0, 3.0 and 4.0 in the presence of A. ferrooxidans and in abiotic solutions in order to determine whether these chemolithotrophic bacteria can be sustained by acid-promoted fayalite dissolution and to measure the impact of their metabolism on acid neutralization rates. The production of almost the maximum Fe3qfrom the available Fe in solution in
Ž 3q .
microbial experiments compared to no production of Fe in abiotic controls confirms A. ferrooxidans metabolism. Furthermore, cell division was detected and the total cell numbers increased over the duration of experiments. Thus, over the pH range 2–4, fayalite dissolution can sustain growth of A. ferrooxidans. However, ferric iron released by A. ferrooxidans metabolism dramatically inhibited dissolution rates by 50–98% compared to the abiotic controls.
Two sets of abiotic experiments were conducted to determine why microbial iron oxidation suppressed fayalite dissolution. Firstly, fayalite was dissolved at pH 2 in fully oxygenated and anoxic solutions. No significant difference was observed between rates in these experiments, as expected, due to extremely slow inorganic ferrous iron oxidation rates at pH 2. Experiments were also carried out to determine the effects of the concentrations of Fe2q, Mg2q and Fe3q on fayalite dissolution. Neither Fe2qnor Mg2qhad an effect on the dissolution reaction. However, Fe3q, in the solution, inhibited both
silica and iron release in the control, very similar to the biologically mediated fayalite dissolution reaction. Because ferric
Ž .
iron produced in microbial experiments was partitioned into nanocrystalline goethite with very low Si that was loosely associated with fayalite surfaces or coated the A. ferrooxidans cells, the decreased rates of accumulation of Fe and Si in solution cannot be attributed to diffusion inhibition by goethite or to precipitation of Fe–Si-rich minerals. The magnitude of
3q Ž .
the effect of Fe addition or enzymatic iron oxidation on fayalite dissolution rates, especially at low extents of fayalite reaction, is most consistent with suppression of dissolution by interaction between Fe3q and surface sites. These results suggest that microorganisms can significantly reduce the rate at which silicate hydrolysis reactions can neutralize acidic solutions in the environment.q2001 Elsevier Science B.V. All rights reserved.
Keywords: Fe-oxidizing bacteria; Fe-silicate mineral dissolution; Thiobacillus ferrooxidans
)Corresponding author. Fax:q1-608-262-0693.
Ž .
E-mail address: [email protected] S.A. Welch .
1
Current address: Woods Hole Oceanographic Institute, Woods Hole MA, USA.
1. Introduction
Microbes utilize Fe for a variety of purposes and can greatly influence biogeochemical cycling of Fe.
0009-2541r01r$ - see front matterq2001 Elsevier Science B.V. All rights reserved.
Ž .
In anoxic environments, microbes can use ferric iron oxyhydroxide phases as electron acceptors for
respi-Ž
ration of organic compounds see Ehrlich, 1996 for .
review , thus, greatly increase rates of Fe release to solution. Under oxidizing conditions, Fe-oxidizing Bacteria and Archaea can utilize reduced Fe as an electron donor in energy generation. This metabolism is generally thought to predominate at low pH, where rates of inorganic iron oxidation are slow. However, a growing body of evidence indicates that these organisms play a significant role in Fe cycling and CO2 fixation in more neutral pH solutions. For example, chemolithotrophic iron oxidizers have been detected in soils and ground water in microenviron-ments where pO2 is low or at oxicranoxic
inter-Ž
faces Emerson and Moyer, 1997; Emerson et al., .
1999 . Nitrate-reducing lithotrophic iron oxidizers Ž have also been detected in anoxic environments e.g.,
.
Straub et al., 2001 . Observations interpreted to sug-gest the activity of Fe-oxidizing microbes associated
Ž
with oceanic basalts have been reported Thorseth et .
al., 1995; Fisk et al., 1998 . In the case of near-neu-tral pH systems, iron-oxidizing microbes are usually detected in groundwater mixing and discharge zones, and the connection between reactions releasing re-duced iron and microbial populations is unclear. Specifically, the link between dissolution kinetics of iron silicates and growth of Fe-oxidizing microbes has received little attention. This is true both at low pH and in near-neutral pH solutions associated with weathered rocks.
Based on thermodynamic calculations, Jakosky
Ž .
and Shock 1998 predicted that approximately 2 g of microorganisms could be supported by the alter-ation of 1 mol of fayalite. However, the thermody-namic argument fails to consider whether the rate of the mineral alteration reaction is sufficient to sustain a community of Fe-oxidizing microorganisms. Re-cently, direct experimental evidence confirmed that populations of Fe-oxidizing can be sustained solely
Ž
by fayalite dissolution at neutral pH Welch et al., .
submitted .
Although Fe oxidizing chemolithotrophic mi-croorganisms may have limited importance in sili-cate mineral alteration now compared to the effect of heterotrophic organisms and plants, it is likely that they have been very important in earlier geologic periods. Phylogenetic analyses of small subunit
ribo-somal RNA sequences indicate that earliest organ-isms for which we have genetic signatures were autotrophs, organisms capable of converting CO to2
Ž .
organic carbon Barns and Nierzwicki-Bauer, 1997 . Furthermore, metabolic gene sequence analyses sug-gest that many autotrophic pathways evolved billions
Ž .
of years ago Bult et al., 1996 . Although the relative importance of inorganic energy sources has changed over geologic time with the evolution of higher life
Ž forms, energy associated with reduced metals
espe-.
cially Fe in silicate minerals may have been and remain quite significant.
The experiments reported here had two purposes. Firstly, we aimed to determine whether rates of
Ž .
dissolution of fayalite Fe SiO2 4 at pH 2–4 are rapid enough and inorganic ferrous iron oxidation reac-tions slow enough, that significant populareac-tions of Fe-oxidizing microorganisms can be sustained. Sec-ondly, we measured the impact of enzymatic iron oxidation on the kinetics of Fe-silicate mineral weathering reactions. Our results have relevance both to analyses of potential metabolic pathways for
sub-Ž .
surface biospheres here and on Mars and to models Ž
that predict acid neutralization rates acid mine .
drainage, acid rain .
2. Experimental methods
2.1. Minerals
Natural fayalite was obtained from the American
Ž .
Museum of Natural History AMNH 10928 and
Ž .
from the Smithsonian R 3516 . Sample R 3516 is from Cheyenne County, CO. Few millimeter-wide crystal aggregates were crushed, sieved, washed and characterized by X-ray diffraction and
high-resolu-Ž .
tion transmission electron microscopy HRTEM and SEM. The 25–75-mm size fraction was used for the experiments. Synthetic fayalite was prepared by slow cooling of a mixture of iron oxalate and quartz from 12758C to 11758C under a COrCO2 atmosphere Žf O –10y1 0.. Synthetic fayalite had a particle size
2
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 101
2.2. Microbes
A lithotrophic iron oxidizing bacterium, Thiobac-illus ferrooxidans, was used in the biotic experi-ments. A. ferrooxidans was cultured in a 9-K media ŽSilverman and Lundgren, 1959 as cited in Ehrlich,
.
1996 on an orbital shaker at 258C at 130 rpm for several weeks. The 9-K media consist of 3.0 g ŽNH4 2. SO , 0.1 g KCl, 0.5 g K HPO , 0.5 g4 2 4
Ž .
MgSO4P7H O, 0.01 g Ca NO2 3 2, 44.22 g FeSO4P
7H O and 1-l autoclaved, distilled water. The bacte-2 ria were separated from their culture media by a series of filtrations. The culture media with bacteria
Ž
was initially filtered using a paper filter approxi-.
mately 10-mm pore size to separate out the iron hydroxide precipitates from the media and bacteria. The filtrate was collected and then, filtered again using an autoclaved 0.22-mm polycarbonate filter to collect the bacterial cells. This filter was flushed with several milliliters of distilled water to remove residual dissolved Fe from the bacterial cells. The filter with bacterial cells was placed in a polycarbon-ate centrifuge tube with a few milliliters of sterile nanopure water. The sample was then vortexed and sonicated to remove cells from the filter. This solu-tion was then examined by light and epifluorescence microscopy for bacterial cells and for iron precipi-tates. Trace quantities of iron precipitates were de-tected in the cell slurry. The cell solution was stored at 48C until the bacteria could be added to the experiments, usually within several hours. All media and solids were cleaned and autoclaved to minimize possible contamination by other microorganisms. A. ferrooxidans was an ideal organism to use in the experiments because it can be grown in extremely
Ž .
simple media see below and because its possible growth substrates are limited to reduced iron and
Ž .
sulfur it cannot utilize organic molecules .
2.3. Biological experiments
Fayalite dissolution experiments were carried out in batch reactors. One hundred milligrams of fayalite was added to 50 ml of solution in acid-washed polycarbonate flasks. The solutions used for all bio-logical experiments consisted of dilute H SO , 12 4
Ž .
mM NaHCO , 1 mM NH3 4 2SO , 1004 mM K HPO2 4
and filtered nanopure water. This media was chosen since it would provide trace inorganic nutrients re-quired by the lithotrophic microorganisms without severely complicating the solution chemistry for the dissolution experiments. Initial solution pH was ad-justed by changing the molarity of sulfuric acid, either 1 or 10 mM and then, titrated with either dilute HCl or NaOH before experiments.
The experiments were done at an initial pH of 2, 3 and 4. Each set of experiments consisted of three different treatments: an abiotic treatment with fay-alite and solution; a biological treatment with ap-proximately 2–3 ml of the cell solution, fayalite and media and a fayalite-free control, which consisted of 2–3 ml of the cell solution in 50 ml of media. There were approximately 107 cellsrml in the bulk solu-tion in the biological and fayalite-free treatment. Cell numbers remained relatively constant throughout the course of the experiments. The fayalite-free control was done to determine the amount of Fe and Si associated with the bacterial cells. There were three replicates for all abiotic and the biological
experi-Ž ments, except for the fayalite-free treatment which
.
was not replicated . Flasks were placed on an orbital shaker at 120 rpm to keep solutions aerated and well mixed during the experiments. Five-milliliter aliquots was taken periodically from each bottle for about 2 weeks using a sterile syringe. These aliquots were filtered with a 0.22-mm acrodisc syringe filter to remove the bacteria and mineral particles. Five milliliters of fresh solution was then added to the flasks to keep the volume constant. All experiments were conducted at room temperature.
Experiments were performed at pH 2 using dead cell controls to determine whether they influenced rates of Fe and Si accumulation in solution. Cells
Ž .
wereAdeenergizedB inactivated, but not dead using
Ž .
1 mM Na N sodium azide from Fisher Chemical3 ŽMera et al., 1992 . For this set of experiments, there. were five treatments, three of which are identical to previous experiments: abiotic, live cells and fay-alite-free. Two sets of flasks contained 1 mM Na N:3 an abioticqazide treatment to determine if Na N3 affected the dissolution reaction and cellsqazide
Ž
treatment to determine the effect of dead or de-en-.
2.4. Abiotic experiments
Since A. ferrooxidans had a significant effect on the fayalite dissolution reaction, three sets of abiotic experiments were conducted to distinguish the effect of solution redox chemistry on fayalite dissolution from effects due to reaction products. All abiotic redox experiments were carried out at an initial solution pH of approximately 2, under conditions very similar to the abiotic controls in the previous experiments.
One set of experiments tested the effect of dis-solved oxygen on fayalite dissolution. The solution was the same used in inorganic controls for the pH 2 biological fayalite dissolution experiments. One hun-dred microliters of solution and 100 mg of natural fayalite were added to each reactor. In the oxic treatment, oxygen was bubbled through the solution for approximately 30 min, initially, and then, for 10 min everyday after a sample was taken. Similarly, nitrogen was bubbled through the solution in order to achieve essentially anoxic conditions. Bottles were not constantly agitated but were swirled gently by hand before samples were taken. A 5-ml aliquot was taken periodically from each bottle over 10 days. Samples were filtered with a 0.22-mm acrodisc filter. Experiments were not replenished with fresh media after sampling in order to minimize gas exchange.
A second set of abiotic fayalite dissolution expe-riments was carried out to determine the effect of solution iron speciation on fayalite dissolution. The Fe2q vs. Fe3q experiments consisted of four different treatments with two replicates each: 10 mM
Ž .
H SO2 4 as a control , 10 mM H SO2 4 plus 1 mM Mg2q, 10 mM H SO2 4 plus Fe2q, and 10 mM H SO plus Fe2 4 3q. One hundred microliters of solu-tion and 100 mg of synthetic fayalite were added to polycarbonate bottles that were placed on a shaker table for 8 days. Samples were taken periodically using a syringe and filtered with a 0.22-mm filter.
A third set of abiotic fayalite dissolution experi-ments was performed to determine the effect the varying amounts of Fe3q in solution had on fayalite
dissolution. Fayalite was reacted in media containing 100 mM Fe3q, 500 mM Fe3q, 1 mM Fe3q and 5
mM Fe3q. The duplicated batch experiments
con-sisted of 100 mg of fayalite reacted in 100 ml of solution in polycarbonate flasks. Samples were
col-lected periodically for 10 days using a syringe and filtered with a 0.22-mm filter. Fresh media was not added after sampling.
2.5. Analytical
Analysis for Fe2qand total Fe was done using the
Ž .
ferrozine method Stookey, 1970 . Absorbance was measured using a Perkin-Elmer UVrVIS spectro-photometer. Dissolved Si was measured using a Technicon AutoAnalyzer II with the molybdate blue method. Solution pH was determined with a Denver Instruments pH meter that had been calibrated using NBS buffers.
At the end of the dissolution experiments, a frac-tion of the mineral sample was removed from the reactor and mounted on a SEM stub with carbon tape. Mineral samples were gently rinsed and al-lowed to air dry. Samples were sputter coated with either platinum or gold and examined with a LEO 982 high-resolution scanning electron microscope. For some experiments, mineral samples were pre-pared for TEM analysis. Mineral grains were gently crushed using a mortar and pestle, suspended in deionized water and placed on a formvar-coated copper grid. Samples were examined using a Philips CM200 ultratwin transmission electron microscope operated at 200 kV.
3. Results
The net concentrations of iron and silica released were measured in both abiotic and biologic experi-ments. Results depended on solution pH and on the presence or absence of microbial populations. Total cell numbers in solution and on mineral surfaces were estimated by direct counts using epifluo-rescence microscopy
3.1. pH 2 experiments
Fayalite dissolution experiments were conducted
Ž . Ž .
with both natural Fig. 1 and synthetic Fig. 2 fayalite. In the abiotic treatments, results were simi-lar for both experiments, though concentrations were approximately 40% lower for the synthetic vs. natu-ral fayalite. Fe and Si release to solution was rapid in the first few day of the experiment and then, leveled
Ž
off or slightly decreased with time due to the addi-.
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 103
Ž . Ž . Ž .
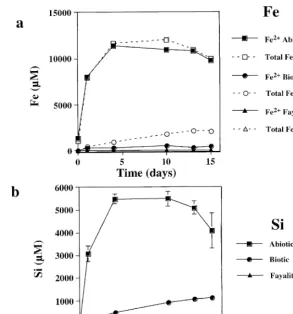
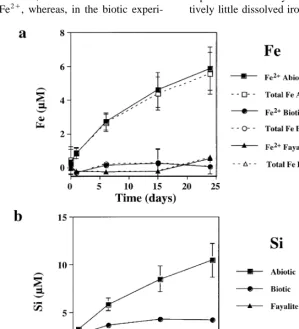
Fig. 1. a Total open symbols and reduced Fe filled symbols concentration for natural fayalite dissolution at initial pH 2 for the controls
ŽB., microbialŽv.and fayalite-freeŽ'.treatments. b Si concentration from dissolving natural fayalite at initial pH 2 for the controlsŽ . ŽB., microbialŽv.and fayalite-freeŽ'.treatments.
largely reflected the rapid consumption of protons in the batch reactor. Bulk solution pH increased rapidly from approximately 2 to 3.6 as the mineral dis-solved. The initial Fe release rate calculated from a linear fit of the Fe concentration curve over time Žbefore Fe concentration becomes relatively
con-. y1 1 2q 2
stant is ;1.3–1.6=10 mol Fe rcm s. Es-sentially, all of the iron in solution in the abiotic experiments was as Fe2q. The Si release curve is
very similar to Fe, though Si concentrations are approximately half those of Fe, reflecting net
stoi-Ž .
chiometric dissolution Figs. 1b and 2b . In the abiotic experiments, approximately 30% to 50% of the fayalite had reacted by the end of the experiment, thus, greatly limiting any contribution to the results from surface layers modified by sample preparation.
In the biological experiments, both Si and Fe release to solution was considerably lower than in the abiotic controls. In the experiments with natural fayalite, final concentrations were about a factor of five lower than the abiotic controls. However, for the experiments with synthetic fayalite, total concentra-tions were approximately 25 to 50 times less than the abiotic experiments. The release of both Fe and Si was nonlinear with time, but did not follow the same rapid initial dissolution observed in the abiotic con-trols. In both biotic experiments, most of the
measur-Ž 3q.
able dissolved Fe is oxidized Fe as opposed to the corresponding abiotic controls where Fe3q was
Ž . Ž . Ž .
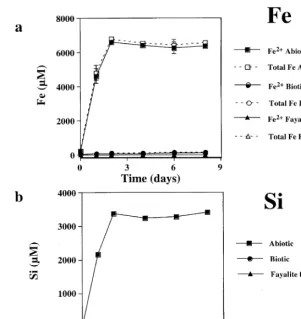
Fig. 2. a Total open symbols and reduced Fe filled symbols concentration for synthetic fayalite dissolution at initial pH 2 for the
Ž . Ž . Ž . Ž .
controls B, microbial v and fayalite-free ' treatments. b Si concentration from dissolving synthetic fayalite at initial pH 2 for the
Ž . Ž . Ž .
controls B, microbial v and fayalite-free ' treatments.
compared to Fe in the synthetic fayalite experiments. In the biotic experiments, solution pH increased from 2 to approximately 2.3. Even though the fayalite is in
Ž .
acidic solution maximum pH ;2.3 , it is appar-ently unreactive compared to the fayalite in the abiotic controls.
The Fe and Si released in the fayalite-free control was either undetectable or negligible compared to all
Ž .
mineral dissolution experiments Figs. 1 and 2 . Since live cultures of A. ferrooxidans appeared to significantly inhibit fayalite dissolution under acidic conditions, the experiments were repeated to deter-mine if the inhibition was the result of cell metabolic processes, or simply the result of adsorption of mate-rial to cell surfaces. The results of the control, biotic and fayalite-free treatments were nearly identical to the previous experiment done at pH 2 with synthetic
Ž .
fayalite Figs. 2 and 3 . Sodium azide had minimal effect on the reaction. Total and reduced Fe released from fayalite in the abiotic azide experiments was only approximately 10% lower than the abiotic con-trol, and Si release was indistinguishable from the control. Similarly, fayalite dissolution in solutions containing azide and dead cells were nearly identical to the abiotic azide treatments. These results indicate that dead cells had no significant effect on the dissolution reaction under these experimental condi-tions.
Solution saturation state with respect to several Fe and Si containing phases was calculated using
Ž .
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 105
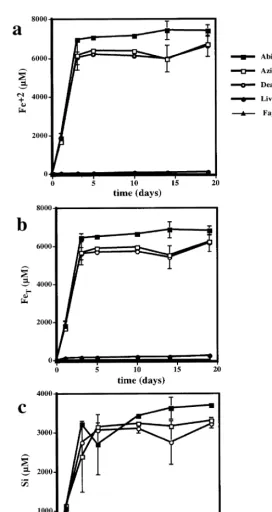
Ž . Ž . Ž . Ž .
Fig. 3. a Reduced, b total and c Si concentrations from dissolving fayalite in dead cell experiments. Treatments are abiotic B, azide
Ž . Ž . Ž . Ž . Ž .
Table 1
Ž .
Saturation index SI with respect to several possible secondary phases for conditions at the end of the dissolution experiments. SI
Ž .
calculated using PHREEQC Parkhurst, 1995
Goethite Hematite Quartz SiO2Ža.
Biological pH 2 1.41 4.83 0.18 y1.09
Biological pH 3 2.76 7.53 y1.12 y2.38 Biological pH 4 3.67 9.35 y1.41 y2.68
Abiotic pH 2 3.00 8.00 1.6 0.33
Abiotic pH 3 2.09 6.18 y0.90 y2.17
Abiotic pH 4 – – y0.98 y2.24
abiotic and biologic samples of the natural and syn-thetic fayalite experiments. Saturation index for con-ditions at the ends of the experiments are given in Table 1.
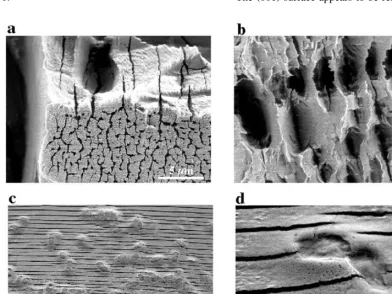
The solution chemical data, in terms of net total Fe and Si released to solution, indicates that approxi-mately 2% to 40% of the fayalite had reacted over the course of the experiments. Fig. 4 shows typical high-resolution SEM images of natural fayalite re-acted at pH 2 for 8 days. Although both sets of minerals show evidence of dissolution, it is evident from the SEM images that minerals in the abiotic experiments are much more extensively reactive than minerals in the biological experiments. Fig. 4a shows a very extensively reacted natural fayalite grain from the abiotic experiments. Deep etch channels and pits formed on the mineral surfaces. Although these etch channels are somewhat irregular, their orientation is generally normal to c). The extent and pattern of
etching varies substantially for different surfaces.
Ž .
The 001 surface appears to be relatively unreacted.
Ž .
Fig. 4. Scanning electron microscope images of natural fayalite reacted in abiotic and biological experiments at initial pH 2. a, b Fayalite reacted in abiotic experiments. The surfaces are characterized by very extensive etching and deep holes and channels forming normal to c). Ž .c Fayalite reacted in with A. ferrooxidans. Etch texture is characterized by narrow planar etch channels normal to c . The micron-sized)
Ž .
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 107
Fig. 4b shows another very rough etched surface
Ž .
with holes and channels forming parallel to 001 . In the biotic experiments, the etching is dominated by semi-periodic parallel channels formed parallel to Ž001. ŽFig. 4c . Based on SEM observations, these. etches appear to be typically 10s to 100s nm wide and spaced approximately 1mm apart. The micron-sized ovals are bacteria on the fayalite surface. Fig. 4d is a higher magnification image of the mineral surface showing dividing cells.
It was difficult to obtain accurate cell counts. Data indicate that the total number of cells in
sus-Ž .
pension increased slightly -a factor of 2 through-out the experiment. However, cell counts do not
include attached cells, as it was generally impossible to detect them using epifluorescence microscopy. Based on estimates from the SEM observations and total cell counts, approximately 10–50% of the cells are attached to mineral surfaces.
3.2. TEM and AEM characterization of reaction products
HRTEM images and energy-dispersive X-ray
Ž .
analyses analytical electron microscopy; AEM show
Ž .
that the nanocrystalline few nanometer in diameter particles of iron oxyhydroxides are produced in bio-logical dissolution experiments. Interplanar spacings in the product indicate a mixture of ferrihydrite and
Ž . Ž . Ž .
Fig. 5. pH 3 a Total open symbols and reduced Fe filled symbols concentration for natural fayalite dissolution at initial pH 3 for the
Ž . Ž . Ž . Ž .
controls B, microbial v and fayalite-free ' treatments. b Si concentration from dissolving natural fayalite at initial pH 3 for the
Ž . Ž . Ž .
goethite. Little Si was detected in secondary miner-als. Particles were typically flocculated into aggre-gates and sometimes adhered to cells. They do not form tightly adhering layers on altered fayalite sur-faces.
3.3. pH 3 experiments
At pH 3, iron and silica release into solution is about three orders of magnitude lower than for the experiment at pH 2 with natural fayalite. This corre-sponds to approximately 0.1% of the solid material reacting during the course of the experiments. Fig. 5 shows Fe and Si concentrations plotted against time. In the abiotic treatment, most of the iron released into solution is Fe2q, whereas, in the biotic
experi-ments with A. ferrooxidans, most of the iron in solution was Fe3q, very similar to what was ob-served in the more acidic experiments. The dissolu-tion reacdissolu-tion was not stoichiometric in both the biotic and abiotic experiments. Si accumulates in solution preferentially compared to Fe. The total
Ž
dissolved FerSi ratio was 0.5 a 4=concentration . of Si compared to Fe, relative to fayalite in the biotic experiments compared to 1.0 in the abiotic experiment. Fayalite dissolution rate at pH 3, based on Si release, is 6.2=10y1 5 molrcm2s in the abi-otic experiment. Overall, the presence of microor-ganisms inhibited the release of silica by approxi-mately a factor of two compared to the abiotic experiments. The fayalite-free control showed rela-tively little dissolved iron or silica in solution.
Ž . Ž . Ž .
Fig. 6. a Total open symbols and reduced Fe filled symbols concentration for natural fayalite dissolution at initial pH 4 for the controls
ŽB., microbialŽv.and fayalite-freeŽ'.treatments. Iron concentration in the biological and fayalite-free treatments are at the detection
Ž . Ž . Ž .
limit for the method. b Si concentration from dissolving natural fayalite at initial pH 2 for the controls B, microbial v and
Ž .
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 109
Ž . Ž .
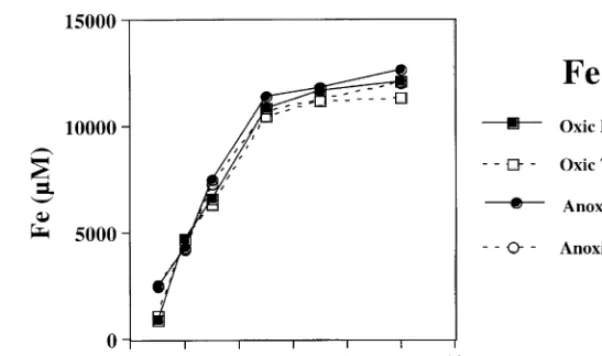
Fig. 7. Reduced and total Fe is solution from synthetic fayalite dissolution under oxic B and anoxic v conditions. Solution pH remained relatively constant with
time for both the abiotic and biologic experiments, increasing from approximately 3.0 to 3.1 throughout the course of the experiment. Solution saturation state with respect to several possible Fe and Si containing phases was calculated using PHREEQ. All experiments are supersaturated with respect to goethite and hematite by the end of the experiment ŽTable 1 ..
3.4. pH 4 experiments
At pH 4, iron and silica release from synthetic fayalite dissolution was approximately three orders
of magnitude lower than for the experiments at pH 2. The abiotic fayalite dissolution rate, based on a linear fit of the Si release curve, is 4.6=10y1 5 molrcm2s. The total Fe and Si released into solution was significantly greater in the abiotic controls
com-Ž .
pared to the biotic experiments Fig. 6 . However, in contrast with the pH 2 and 3 experiments, total Fe in the pH 4 experiments was dominated by Fe2q in both the abiotic and biologic experiments. However, Fe concentrations are extremely low in the biological experiments, very close to the detection limit for the analytical method. The reaction was not stoichiomet-ric. Silica accumulates in solution preferentially over
Ž iron, especially for the biologic experiments where
.
iron is barely detectable . The fayalite-free control
2qŽ . 2qŽ . 3qŽ . Ž .
showed no detectable iron or silica in solution. Solu-tion pH remained relatively constant increasing from approximately 4.0 to 4.3 for both the biologic and
abiotic samples. Solutions were supersaturated with respect to goethite and hematite in the biologic
ex-Ž .
periments Table 1 .
Ž . Ž . Ž . Ž . Ž .
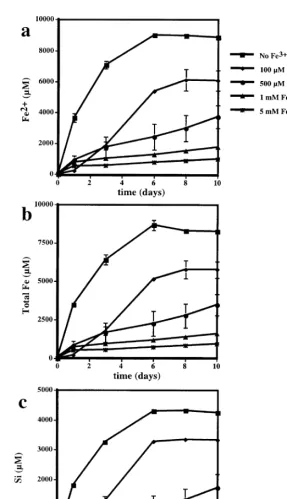
Fig. 9. a Reduced iron, b total iron and c Si from fayalite dissolution at initial pH 2 in a control B, and with 100mM l, 500mM
Žv., 1 mMŽ . Ž . 3q
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 111
4. Effect of solution redox conditions on fayalite dissolution
4.1. AnoxicÕs. oxic experiments
Abiotic experiments were carried out at pH 2 to determine if the dissolved oxygen concentration af-fected the fayalite dissolution rate. Fig. 7 shows iron released into solution for both oxygenated and anoxic Žor low oxygen conditions. Results were very simi-. lar to the natural fayalite abiotic experiments at pH 2. Dissolved Fe increased with time for the first few days of the experiment and then, the Fe concentra-tion was approximately constant. The total Fe was essentially all as Fe2q, even in the solutions that
were supersaturated with respect to O . There was2 no significant difference between Fe release rate, and Fe concentration in the two experiments, indicating that under these experimental conditions, dissolved O has a negligible effect on the fayalite dissolution2 reaction.
4.2. Fe2q and Mg2q experiments
Additional experiments were carried out at pH 2 to determine the effect of the concentration of other ions, Fe2qand Mg2q, on natural fayalite dissolution.
Fig. 8 shows average total iron released into solution over the course of the experiments. Results are nearly identical to the previous abiotic experiments. All dissolved iron released is Fe2q. Neither the added
Fe2q nor Mg2q in solution had a significant effect
on fayalite dissolution compared to the sulfuric acid control.
4.3. Fe3q addition experiments
Addition of 1 mM Fe3q greatly inhibited iron release from the fayalite. Total Fe released to solu-tion from natural fayalite in the Fe3q treatment was approximately 30% of the control at the end of the
Ž .
experiment Fig. 8 . This effect was comparable to the apparent biological inhibition observed for
fay-Ž .
alite dissolution for similar conditions Fig. 1 . Since both the biotic and abiotic experiments appear to show a link between Fe3q and inhibition
of the dissolution reaction, a series of experiments 3qŽ
were carried out with increasing amounts of Fe 0,
. Ž .
0.1, 0.5, 1, 5 mM in solution Fig. 9 . Fe and Si
concentrations released into solution in the control experiments were very similar to the abiotic control experiments done at pH 2 with synthetic fayalite ŽFigs. 2 and 3 . The reaction was stoichiometric. All. the dissolved iron was reduced and approximately, 40% of the fayalite dissolved by the end of the experiment. Addition of 100 mM Fe3q to solution decreased Fe and Si release by almost 1r3 compared to the controls. However, the reaction was not stoi-chiometric. The dissolved total Fe was approxi-mately 1000 mM lower than the concentration pre-dicted from Si for the stoichiometric dissolution of fayalite. The lost Fe was approximately equivalent to the precipitation of a goethite layer 150-nm thick over the initial mineral surface.
The addition of 500 and 1000 mM Fe3q to
solu-tion decreased net Fe and Si release from fayalite by 60% and 90% compared to the control. The reaction was approximately stoichiometric, though the mea-sured total Fe in solution was slightly lower than added Fe3q plus Fe2q predicted from Si release from fayalite dissolution. This difference between measured and predicted iron would be equivalent to a goethite layer that was only a few tens nanometers thick over the initial mineral surface area.
Net Si release from fayalite in the experiments with 5 mM Fe3q is only 3% of Si in the control at
the end of the experiment. Approximately, 80% to 90% of the added iron precipitates during the experi-ment. This could correspond to a goethite layer 600–700-nm thick over the mineral surface.
5. Discussion
5.1. Abiotic rates
sec-ondary phases and chemical affinity for fayalite dis-solution approaches zero. Rates also decrease with time because 30–50% of the available material re-acts within the first few days of the dissolution experiments.
Abiotic fayalite dissolution rates can be calculated from the initial stage of the reaction where Si and Fe are rapidly released to solution and no precipitation occurs. The abiotic dissolution rate of fayalite deter-mined from the increase in Fe or Si in solution ranged from approximately 1=10y1 1 molrcm2s at initial pH 2 to 5=10y1 5 molrcm2s at initial pH 4. These rates are comparable to other experimentally determined dissolution rates for Fe-silicate minerals.
Ž .
For example, Westrich et al. 1993 calculated a fayalite dissolution rate of 6=10y1 1 molrcm2
s at pH 2, slightly faster than the rates determined in this study. In our pH 2 experiments, acidity decreased with time as the mineral dissolved, therefore, the rate is for an average pH of approximately 2.4. Wogelius
Ž . Ž .
and Walther 1992 measured fayalite Fo6 dissolu-tion rates in flow through reactors of 5=10y1 2 to 7=10y1 4 molrcm2s from pH 2 to 5, rates very similar to those we estimated from our batch experi-ments. Experimentally determined mineral dissolu-tion rates typically vary by as much as on order of magnitude depending on experimental conditions, i.e., batch or flow through reactors, mineralrsolution ratio, mineral composition, mineral preparation, pre-treatment, etc., so our experimental rates are consis-tent with others. However, mineral dissolution ex-periments conducted, where all conditions are held constant and only one parameter is varied, tend to be
Ž .
very reproducible see for example Figs. 2 and 3 , and differences between experiments, either in terms of rate of release of mineral components or concen-trations in solution, can be attributed to the affect of
Ž
that variable i.e., changing solution pH, the presence .
of microbes, dissolved O content, etc. .2
5.2. Effect of T. ferrooxidans on fayalite dissolution
Fayalite dissolution experiments conducted in the presence of iron-oxidizing microbes showed a sup-pression of both iron and silica release into solution compared to abiotic controls. SEM images, which show mineral grains from the abiotic experiments are
much more reacted than those from the biologic experiments of the reacted minerals at pH 2, con-firming the solution geochemical results. The appar-ent inhibition of the reaction could be due to several factors.
A decrease in mineral dissolution rates over time has been attributed to the formation of a diffusion inhibiting leached layer or secondary precipitates on
Ž
the mineral surface Schott and Berner, 1983, 1985; .
Chou and Wollast, 1984 .
The formation of an iron-rich product on the fayalite surface is consistent with the solution chem-istry, which shows a preferential accumulation of Si compared to Fe in solution in the biological experi-ments. Furthermore, solutions are apparently super-saturated with respect to several possible secondary
Ž . 3q
Fe phases Table 1 . In the Fe addition experi-ments, the inhibition increases with increasing Fe3q
added to solution. In the biotic experiments at pH 2, total Fe concentration at the end of the dissolution experiment is approximately 150 mM less than what would have been predicted from Si concentration for stoichiometric fayalite dissolution. If all this iron precipitated as goethite, it would form a 15-nm-thick
Ž
layer over the fayalite surface assuming a fayalite surface area of 1000 cm2rg and no precipitation in
.
solution or the reaction vessel . However, there is no evidence, at least in the biotic experiments, that the goethite has formed a layer that could inhibit diffu-sion of ions to or from the mineral surface. TEM Žalso see Welch and Banfield, in press and SEM. images show no sign of an extensive leached layer or precipitated layer on the mineral surface. In fact, the particulate form of the goethite produced suggests that this product does not significantly modify the reactivity of the fayalite surface. Furthermore, sur-face reaction rates and therefore, dissolution rates, are generally thought to be considerably slower than diffusion rates. Thus, we conclude that a diffusion-inhibiting layer composed of iron oxyhydroxide min-erals does not explain the suppression of fayalite dissolution rates in the presence of iron-oxidizing microorganisms.
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 113
considering the total amount of material reacted in these experiments is not sufficient to produce a one unit cell thick layer over the mineral surface.
Another explanation for the suppression of fay-alite dissolution rates in the presence of microorgan-isms or Fe3qis that Fe3q absorbs to and chemically
passivates the mineral surface. The chemical passiva-tion of the surfaces has been suggested for other minerals. For example, Al3q adsorption to feldspar
surfaces appears to substantially decrease their reac-Ž tivity under some experimental conditions e.g.,
.
Oelkers et al., 1994 by forming stable cross-links between isolated Si tetrahedra. Alternatively, other experimental studies have failed to show a
signifi-Ž
cant inhibitory affect e.g., Chen and Brantley, 1997,
. 3q 3q
2000 . Fe , like Al , should be able to adsorb to mineral surfaces. The rates of solvent exchange around the Fe3qion are about three orders of magni-tude slower than rates surrounding Fe2q, therefore,
3q `
the reactivity of the Fe O bond should be orders 2q
` Ž
of magnitude slower than the Fe O bond Casey .
et al., 1993; Casey and Westrich, 1992 . Conse-quently, fayalite surfaces that have adsorbed Fe3qor
Fe ions that have been oxidized should be signifi-cantly less reactive than those with Fe2q surface
sites. Furthermore, naturally weathered olivine is highly reactive, except in regions containing planar defects of laihunite, an olivine in which Fe3q is
substituted primarily in the M2 octahedral sites and
Ž wx Ž 2q. Ž 3q. .
M1 vacancies i.e., x Fe 2y3 x Fe 2 xSiO4 ŽBanfield et al., 1989, 1990, 1992; Casey et al.,
.
1993 . The lower reactivity of ferric iron-rich olivine is consistent with a passivation mechanism due to adsorbed ferric iron as proposed here.
5.3. Effect of solution redox chemistry on fayalite dissolution
It is apparent from our experiments that solution redox reactions can dramatically affect rates and mechanisms of Fe-silicate mineral weathering. Our experiments clearly demonstrate that Fe3q in
solu-tion, either produced by A. ferrooxidans or added to solution abiotically, inhibited fayalite dissolution un-der acidic conditions. The effect increased with in-creasing Fe3q in solution. The presence of dissolved oxygen, which can abiotically oxidize Fe2qto Fe3q,
had no detectable affect on the reaction. However, under the experimental conditions, the abiotic oxida-tion rate of Fe2q is extremely slow, approximately
Ž .
0.1 mmolrlrday Nordstrom and Southam, 1997 , therefore, Fe3qwas not detected during the duration of the abiotic experiments. This example clearly emphasizes that microorganisms can dramatically al-ter the rates and products of geochemical reactions by ensuring that reactions occur under conditions that would not be encountered in the absence of a
Ž .
catalyst the enzymes of the electron transport chain . Others researchers have reported a decrease in Fe-silicate reaction rates under oxic conditions due to the oxidation of Fe2q. For example, Wogelius and
Ž .
Walther 1992 noted a decrease in fayalite dissolu-tion rates over time in their flow through reactors. They attributed this to the slow oxidation of reduced iron and precipitation of ferric hydroxides on the
Ž .
mineral surface. Siever and Woodford 1979 saw an increase in dissolution of many Fe containing rocks and minerals, fayalite, hypersthene, basalt and obsid-ian under slightly acidic anoxic conditions compared to air saturated or O2 saturated solutions. Total amount of material released to solution was up to an order of magnitude under anoxic conditions greater than comparable experiments in equilibrium with air or O , presumably due to the precipitation of iron2 phases on mineral surfaces. Alternatively, several researchers reported an increase in dissolutionr weathering rate of Fe silicates, due either to oxygen
Ž .
or another oxidizing agent. Hoch et al. 1996 saw a significantly higher flux of Si, Ca and Mg from augite in their 1.5-ppm O experiments at near-neu-2 tral pH compared to their 0.6-ppm O experiments,2 whereas, dissolved O content had no apparent effect2 on diopside dissolution. Diopside dissolution rates were lower than augite rates. They attributed the increase in reaction rate with increasing dissolved O2 and faster rates in Fe-rich minerals to the oxidative dissolution of Fe from the mineral. In a similar
Ž .
study, White and Yee 1985 measured an enhance-ment of augite dissolution in the presence of dis-solved O and Fe3q. They attributed this increase in
2
reaction rate to an electron transfer between Fe3q in
solution and Fe2qon the mineral surface. The
result-ing formation of a surface Fe3q leads to a charge
5.4. Microbial growth from Fe silicate minerals
If mineral dissolution rates and microbial iron oxidation rates are known, then, its possible to esti-mate the numbers of iron oxidizing microorganisms that could be sustained by iron oxidation. Based on an iron oxidation rate of ;6=10y1 8 mol Fe2qr
Ž .
cells Welch et al., in preparation and a fayalite
y1 0 y11 2 Ž dissolution rate of ;10 –10 molrcm s from
.
Westrich et al., 1993 , we predict that approximately 108–109cells can be sustained by fayalite dissolution at pH 2 in our experiments over the course of the experiment. This is consistent with the numbers of cells detected on our experiments for slightly slower reaction rates.
5.5. Implications for rates of acid neutralization by silicate mineral hydrolysis
The results of our experiments may be relevant to a variety of natural processes, as well as to reactions that occur in sites impacted by acid rain or acid drainage. Even though bulk solution pH for most natural environments is typicallyAnear neutralB, it is not unreasonable that in microbial microenviron-ments, conditions could be significantly more acidic ŽBarker et al., 1998 . Results of this study show that. under such conditions, iron silicate dissolution reac-tions could sustain significant microbial populareac-tions. Our results do not rule out the ability of silicate dissolution reactions to sustain microbial populations at near-neutral pH, but it is probable that the number of organisms would be lower due to the slower Fe-silicate dissolution rate and the communities would be confined to low oxygen environments where inorganic iron oxidation rates are slow. Neu-trophilic iron-oxidizing microbes are very difficult to cultivate in the laboratory, so they were not used in the current experiments. However, they may be
com-Ž mon in natural microaerophilic environments
Emer-. son and Moyer 1997; Emerson et al., 1999 includ-ing environments expected at Fe-silicate surfaces early in chemical weathering of rocks.
At near-neutral pH, the solubility of ferric iron is so low that, once formed by enzymatic routes, it
Ž precipitates very close to the oxidation site e.g., on
. the cell surface; see Banfield et al., 2000 . Under these conditions, it is likely that ferric iron
suppres-sion of fayalite dissolution will not occur. Thus, we predict that fayalite rates will not be as strongly inhibited by iron oxidation in near-neutral pH
solu-Ž
tions in contrast to the process at low pH, where ferric iron has a significant residence time in
solu-. tion .
6. Conclusion
The results of our experiments demonstrate that fayalite dissolution at low pH is able to sustain an active population of the iron-oxidizing bacterium T. ferrooxidans. Metabolic activity is evidenced by the production of almost the maximum concentration of ferric iron in solution, persistence and increase in size of populations over the duration of experiments, production of precipitates in proximity to cells and presence of dividing cells on the fayalite surface. Our results also show that the fayalite dissolution rate increases by three orders of magnitude between pH 4 and 2 implying a much larger microbial popu-lation can be sustained at low pH. However, the activity of the iron oxidizing bacterium suppresses the rates of fayalite dissolution at low pH. The results of abiotic, ferric iron-supplemented controls, are attributed to oxidation of Fe2q to Fe3q under conditions where ferric iron remains relatively solu-ble for long enough to interact with the fayalite surface. These results are relevant to conditions en-countered during neutralization of acid mine drainage or when acid rain interacts with silicate rocks Žolivine-group minerals being the most reactive of
. the common silicate phases .
References
Banfield, J.F., Veblen, D.R., Jones, B.F., 1989. TEM of laihunite-olivine intergrowths in oxidized olivine, and their influence on weathering reactions. Geological Society of
Ž .
America, 1989 annual meeting., 21 Ed. Anonymous , 240.
Ž .
Geological Society of America GSA .
Banfield, J.F., Veblen, D.R., Jones, B.F., 1990. Transmission electron microscopy of subsolidus oxidation and weathering of olivine. Contrib. Mineral. Petrol. 106, 110–123.
( )
C.M. Santelli et al.rChemical Geology 180 2001 99–115 115
Banfield, J.F., Welch, S.A., Zhang, H., Ebert, T.T., Penn, R.L., 2000. Crystal growth and microstructural evolution of FeOOH biomineralization products. Science 289, 751–754.
Barker, W.W., Welch, S.A., Chu, S., Banfield, J.F., 1998. Experi-mental observations of the effects of bacteria on aluminosili-cate weathering. Am. Mineral. 83, 1551–1563.
Barns, S.M., Nierzwicki-Bauer, D.H., 1997. Microbial diversity in ocean, surface and subsurface environments. In: Banfield, J.F.,
Ž .
Nealson, K.H. Eds. , Geomicrobiology: Interactions between Microbes and Minerals. Mineral. Soc. Am. Rev. Mineral., vol. 35, pp. 35–79.
Bult, C.J., White, O., Olsen, G.J., Zhou, L.X., Fleischmann, R.D., Sutton, G.G., Blake, J.A., FitzGerald, L.M., Clayton, R.A., Gocayne, J.D., Kerlavage, A.R., Dougherty, B.A., Tomb, J.F., Adams, M.D., Reich, C.I., Overbeek, R., Kirkness, E.F., Weinstock, K.G., Merrick, J.M., Glodek, A., Scott, J.L., Ge-oghagen, N.S.M., Weidman, J.F., Fuhrmann, J.L., Nguyen, D., Utterback, T.R., Kelley, J.M., Peterson, J.D., Sadow, P.W., Hanna, M.C., Cotton, M.D., Roberts, K.M., Hurst, M.A., Kaine, B.P., Borodovsky, M., Klenk, H.P., Fraser, C.M., Smith, H.O., Woese, C.R., Venter, J.C., 1996. Complete genome sequence of the methanogenic archaeon,
Methanococ-cus jannaschii. Science 273, 1058–1073.
Casey, W.H., Westrich, H.R., 1992. Control of dissolution rates of orthosilicate minerals by divalent metal–oxygen bonds. Nature
ŽLondon 355, 157–159..
Casey, W.H., Banfield, J.F., Westrich, H.R., McLaughlin, L., 1993. What do dissolution experiments tell us about natural weathering. Chem. Geol. 105, 1–15.
Chen, Y., Brantley, S.L., 1997. Temperature and pH-dependence of albite dissolution rate at acid pH. Chem. Geol. 135, 275– 292.
Chen, Y., Brantley, S.L., 2000. Dissolution of fosteritic olivine at 658C and 2-pH-5. Chem. Geol. 165, 267–281.
Chou, L., Wollast, R., 1984. Study of the weathering of albite at room temperature and pressure with a fluidized bed reactor. Geochim. Cosmochim. Acta 48, 2205–2217.
Ehrlich, H.L., 1996. Geomicrobiology. Marcel Dekker, New York, 717 pp.
Emerson, D., Moyer, C., 1997. Isolation and characterization of novel iron-oxidizing bacteria that grow at circumneutral pH. Appl. Environ. Microbiol. 63, 4784–4792.
Emerson, D., Weiss, J.V., Megonigal, J.P., 1999. Iron-oxidizing
Ž
bacteria are associated with ferric hydroxide precipitates
Fe-.
plaque on the roots of wetland plants. Appl. Environ. Micro-biol. 65, 2758–2761.
Fisk, M.R., Giovannoni, S.J., Thorseth, I.H., 1998. Alteration of oceanic volcanic glass: textural evidence of microbial activity. Science 281, 978–980.
Hoch, A.R., Reddy, M.M., Drever, J.I., 1996. The effect of iron content and dissolved O on dissolution rates of clinopyroxene2 at pH 5.8 and 258C: preliminary results. Chem. Geol. 132, 151–156.
Jakosky, B.M., Shock, E.L., 1998. The biological potential of Mars, the early Earth, and Europa. J. Geophys. Res. 103, 19359–19364.
Mera, M.U., Kemper, M., Doyle, R., Beveridge, T.J., 1992. The membrane-induced proton motive force influences the metal binding ability of Bacillus subtillis cell walls. Appl. Environ. Microbiol. 58, 3837–3844.
Nordstrom, D.K., Southam, G., 1997. Geomicrobiology of sulfide
Ž .
mineral oxidation. In: Banfield, J.F., Nealson, K.H. Eds. , Geomicrobiology: Interactions Between Microbes and Miner-als. Mineral. Soc. Am. Rev. Mineral., vol. 35, pp. 361–390. Oelkers, E.H., Schott, J., Devidal, J.L., 1994. The effect of
aluminum, pH, and chemical affinity on the rates of alumino-silicate dissolution reactions. Geochim. Cosmochim. Acta 58, 661–669.
Parkhurst, D.L., 1995. User’s guide to PHREEQC-A computer program for speciation, reaction-path, advective-transport, and inverse geochemical calculations. Water-Resources Investiga-tion Report 95-4227, U.S. Geological Survey, Lakewood, CO, pp. 143.
Schott, J., Berner, R.A., 1983. X-ray photoelectron studies of the mechanism of iron silicate dissolution during weathering. Geochim. Cosmochim. Acta 47, 2233–2240.
Schott, J., Berner, R.A., 1985. Dissolution mechanisms of
pyrox-Ž .
enes and olivines during weathering. In: Drever, J.I. Ed. , 1985. The Chemical of Weathering, vol. 149, Reidel, Dor-drecht, The Neatherlands, pp. 35–53.
Siever, R., Woodford, N., 1979. Dissolution kinetics and the weathering of mafic minerals. Geochim. Cosmochim. Acta 43, 717–724.
Silverman, M.P., Lundgren, D.G., 1959. Studies on the chemoau-totrophic iron bacterium Ferrobacillus ferrooxidans: I. An improved medium and a harvesting procedure for securing high cell yields. J. Bacteriol. 78, 326–331.
Straub, K.L., Marcus, B., Schink, B., 2001. Iron metabolism in anoxic environments at near neutral pH. FEMS Microbiol. Ecol. 34, 181–186.
Stookey, L.L., 1970. Ferrozine—a new spectrophotometric reagent for iron. Anal. Chem. 42, 779–781.
Thorseth, I.H., Torsvik, T., Furnes, H., Muehlenbachs, K., 1995. Microbes play an important role in the alteration of oceanic crust. Chem. Geol. 126, 137–146.
Welch, S.A., Banfield, J.F., in press. Ferric iron ordering and microstructural evolution of olivine during biological dissolu-tion: Reproduction of features of natural weathering. Welch, S.A., Newman, D.K., Labrenz, M., Santelli, C.M.,
Ed-wards, K.J., Bond, P.L., Banfield, J.F., in preparation. Micro-bial populations sustained by dissolution of Fe-silicates, Na-ture.
Westrich, H.R., Cygan, R.T., Casey, W.H., Zemitis, C., Arnold, G.W., 1993. The dissolution kinetics of mixed-cation orthosili-cate minerals. Am. J. Sci. 293, 869–893.
White, A.F., Yee, A., 1985. Aqueous oxidation-reduction kinetics associated with couple electron-cation transfer from iron-con-taining silicates at 258C. Geochim. Cosmochim. Acta 49, 1263–1275.