EXPERIMENT MESELSON STAHL ROLLING CIRCLE REPLICATION
REVERSE TRANSCRIPTION REPLIKASI APARATUS PHAGE T4
RESUME
Disusun untuk memenuhi tugas matakuliah Genetika Lanjut yang dibimbing oleh Prof. Dr. Siti Zubaidah, M.Pd
Oleh:
Kelompok 4 / Kelas B
Dewi Cahyaningrum (140341807059)
Ari Ashari (150341808373)
Frengky Neolaka (150341806355)
UNIVERSITAS NEGERI MALANG PROGRAM PASCASARJANA
PROGRAM STUDI PENDIDIKAN BIOLOGI FEBRUARI 2016
EXPERIMENT MESELSON STAHL
Dasar Replikasi DNA secara in Vivo
Berdasarkan hasil penemun ahli genetika, pada manusia sintetis 1 untai DNA baru terjadi kira-kira dengan kecepatan 3000 nukleotida per-menit. Sedangkan pada sel bakteri sekitar 30.000 nukleotida yang ditambahkan pada untai DNA yang baru dibentuk per-menitnya. Terdapat mekanisme seluler yang bertanggung jawab pada proses replikasi DNA yang mengharuskan replikasi terjadi dengan cepat dan akurat. Keakuratan replikasi DNA sangat menakjubkan, diperoleh rata-rata hanya terjadi 1 kesalahan 1 milyar pada saat penggabungan nukleotida yang terjadi setelah sintesis dan pengoreksian kesalahan pada saat selama replikasi.Sintesis DNA seperti halnya sintesis RNA, dan protein yang terjadi dalam 3 tahap yaitu: (1) Inisiasi, (2) Elongasi, (3) Terminasi.
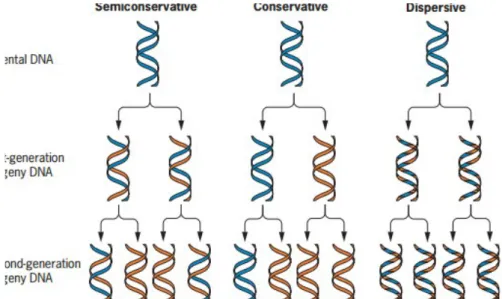
DNA merupakan molekul hidup karena mampu melakukan penggandaan diri (replikasi). Fungsi ini disebut fungsi autokatalisis karena DNA mampu mensistesis dirinya sendiri. Replikasi merupakan peristiwa sintesis DNA. Replikasi DNA dapat terjadi dengan adanya sintesis rantai nukleotida baru dari rantai nukleotida lama. Prosesnya dengan menggunakan komplementasi pasangan basa untuk menghasilkan suatu molekul DNA baru yang sama dengan molekul DNA lama. Proses yang terjadi tersebut dipengaruhi oleh enzim helikase, enzim polimerase, dan ligase. Ada tiga kemungkinan terjadinya replikasi DNA, yaitu semikonservatif, konservatif, dan dispersif.
Replikasi Semikonservatif
Ketika Watson dan Crick mempublikasikan mengenai DNA yang menyatakan bahwa DNA memiliki struktur double helix mengusulkan bahwa dua rantai yang komplemen pada double heliks membuka dan terpisah. tiap-tiap untainya membentuk untai baru. Urutan basa tiap untai DNA “parent” digunakan sebagai template dan basa akan berpasangan dengan komplemennya sehingga membentuk untai baru. Sebagai contohnya Adenin yang terdapat pada untai DNA “parent” yang akan bertindak sebagai template yang akan dihubungkan oleh ikatan hidrogen agar dapat berpasangan dengan Timin yang terdapat pada untai DNA yang baru terbentuk. Mekanisme ini
disebut dengan replikasi semikonservatif (karena setengah molekul “induk/ parent” masih tetap terjaga).
Gambar 1. Model replikasi semikonservatif DNA oleh Watson dan Crick
Pada replikasi konservatif, DNA indukan akan tetap terjaga/dipertahankan, sedangkan untai baru dari DNA akan disintesis. Sedangkan pada model replikasi dispersive, segmen dari kedua molekul untai DNA akan tetap dipertahankan dan digunakan sebagai template untuk mensintesis segmen yang komplemen yang setelah itu akan bergabung untuk menghasilkan keturunan untai DNA yang baru.
Gambar 2. Tiga model yang memungkinkan terjadi pada saat replikasi DNA yaitu (a) semikonservatif, (b) konservatif, (c) dispersif
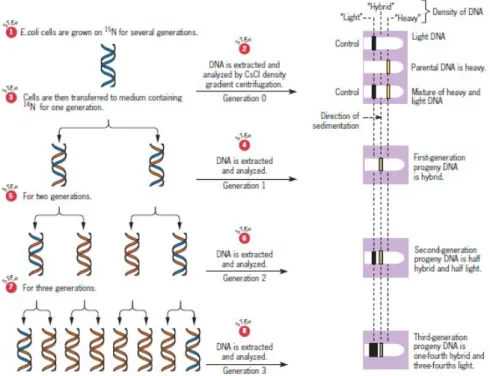
Pada tahun 1958, Matthew Meselson dan Franklin Stahl mendemonstrasikan sebuah replikasi semikonservatif pada kromosom Eschericia Coli. Kemudian pada 1962, John Cairns mendemonstasikan bahwa kromosom dari E. coli memiliki DNA dengan untai ganda. Hasil presentasi dari Cairns, Meselson, dan Stahl bersama-sama mereka ingin membuktikan bahwa sebenarnya DNA bereplikasi secara semikonservatif pada E.coli.
Meselson dan Stahl menumbuhkan sel bakteri E.coli di dalam medium isotop berat nitrogen (15N) yang telah disubstitusi menjadi normal, yaitu isotop ringan (14N)
sebagai penanda pada masing-masing medium. Basa purin dan pirimidin pada DNA mengandung nitrogen. Kemudian DNA sel bakteri tumbuh pada medium yang mengandung isotop (15N) dan memiliki kepadatan yang lebih tinggi (massa/unit
volume) dari pada DNA sel bakteri yang tumbuh pada medium yang mengandung isotop (14N). Molekul DNA dengan kepadatan yang berbeda dapat dipisahkan oleh
equilibrium density-gradient centrifugation. Dengan menggunakan teknik inilah, Meselson dan Stahl mampu membedakan diantara ketiga model replikasi DNA melalui bentuk perubahan kepadatan pada DNA sel bakteri yang ditumbuhkan pada medium (15N) dan medium (14N) yang disebut dengan eksperimen perpindahan
kepadatan.
Meselson dan Stahl mengambil sel-sel yang telah tumbuh pada medium yang mengandung (15N) selama beberapa generasi (sehingga terkandung "berat" DNA),
kemudian sel-sel tersebut dicuci untuk menghapus medium yang mengandung (15N),
dan dipindahkan pada medium yang mengandung (14N). Selanjutnya sel dibiarkan
tumbuh pada medium (14N) pada periode waktu, kemudian DNA diekstraksi dan
dianalisis dalam CsCl untuk mencapai keseimbangan gradien kepadatan. Hasil penelitian Meselson dan Stahl konsisten menunjukkan bahwa replikasi hanya dengan replikasi semikonservatif. Semua DNA yang diisolasi dari sel setelah satu generasi pertumbuhan pada medium yang mengandung (14N) memiliki setengah kerapatan
antara atau kerapatan antara. Kepadatan menengah ini biasanya disebut sebagai kepadatan "hybrid". Setelah pertumbuhan dua generasi pada medium yang mengandung (14N), setengah dari DNA merupakan kerapatan hybrid.
Gambar 3. Meselson dan Stahl mendemonstrasikan replikasi DNA semikonservatif di E. coli. Diagram tersebut menunjukkan bahwa hasil percobaan Meselson dan Stahl yang diharapkan jika kromosom E. coli bereplikasi semikonservatif.
Salah satu dari generasi hasil replikasi semikonservatif dari helix ganda induk yang mengandung 15N pada medium yang mengandung 14N hanya akan menghasilkan
dua keturunan heliks ganda, yang keduanya memiliki 15N dalam satu untai (strand
yang lama) dan 14N di untai lainnya (strand yang baru). Molekul tersebut akan
menjadi kepadatan hybrid.
Replikasi konservatif tidak menghasilkan molekul DNA dengan kepadatan hybrid, karena setelah dihasilkan satu generasi DNA hasil dari replikasi konservatif berat DNA dalam medium menjadi ringan, sehingga setengah dari DNA masih akan memiliki berat yang lebih apabila dibandingkan dengan setengah lainnya yang akan menjadi lebih ringan. Jika replikasi dispersif, Meselson dan Stahl mengamati perubahan berat DNA terhadap cahaya dalam setiap generasi (yaitu dengan mengamati setengah berat atau hybrid setelah satu generasi, kuartal berat setelah dua generasi, dan sebagainya). Hasil dari percobaan Meselson dan Stahl memiliki kemungkinan-kemungkinan yang tidak konsisten. Replikasi DNA selanjutnya terbukti terjadi secara semikonservatif di beberapa mikroorganisme.
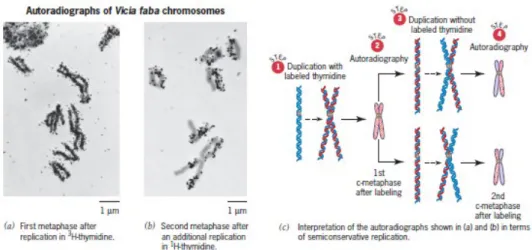
Replikasi semikonservatif pada kromosom eukariotik pertama kali dibuktikan pada tahun 1957 dengan hasil percobaan yang dilakukan oleh J. Herbert Taylor, Philip Woods, dan Walter Hughes pada sel-sel ujung akar Vicia faba. Taylor dan rekannya memberikan label pada kromosom Vicia faba yang diambil bagian pertumbuhan ujung akarnya selama delapan jam (kurang dari satu generasi sel) pada medium yang mengandung radioaktif 3H-timidin. Sel-sel pada ujung akar tersebut kemudian
dipindahkan dari medium yang mengandung radioaktif, dicuci, dan dipindahkan pada medium non radioaktif yang mengandung alkaloid colchicine. Hal tersebut dilakukan karena diketahui bahwa colchicine dapat mengikat mikrotubulus dan mencegah pembentukan spindle fungsional. Akibatnya, kromosom keturunannya tidak mengalami anafase normal. Dengan demikian, jumlah kromosom per inti akan berlipat ganda per siklus sel apabila menggunakan colchicine. Hal tersebut mengakibatkan jumlah kromosom dua kali lipat untuk setiap generasi sel. Taylor dan rekan-rekannya menentukan berapa banyak DNA ulangan dari setiap sel yang telah mengalami penggabungan tersebut dari timidin radioaktif. Pada metafase pertama di colchicine (c-metafase), inti akan berisi 12 pasang kromatid (masih bergabung di sentromer). Pada kedua c-metafase, inti akan berisi 24 pasang, dan sebagainya.
Taylor dan rekan-rekannya menggunakan teknik yang disebut autoradiografi untuk memeriksa distribusi radioaktivitas dalam kromosom pada sel pada saat c-metafase yang pertama, c-c-metafase kedua, dan sebagainya. Autoradiografi adalah metode untuk mendeteksi dan lokalisasi radioaktif isotop dalam persiapan sitologi atau makromolekul oleh pencahayaan fotografi terhadap emulsi yang sensitif terhadap radiasi energi rendah. Emulsi tersebut mengandung halida perak yang menghasilkan bintik-bintik hitam kecil disebut butir perak. Ketika butiran perak terkena partikel bermuatan yang dipancarkan selama peluruhan isotop radioaktif. Autoradiografi sangat berguna dalam mempelajari metabolisme DNA karena DNA dapat diberi label khusus menurut sel yang tumbuh di 3H-timidin, adeoxyribonucleoside timin yang berisi isotop radioaktif hydrogen (tritium).
Ketika Taylor dan rekannya menggunakan autoradiografi untuk memeriksa distribusi radioaktivitas dalam kromosom Vicia faba pada saat pertama c-metafase, kedua kromatid dari setiap pasangan merupakan radioaktif. Namun pada c-metafase yang kedua, hanya salah satu kromatid dari setiap pasangan yang merupakan radioaktif. Hasil tersebut merupakan yang diharapkan jika replikasi DNA adalah
semikonservatif, memberikan molekul DNA satu per kromosom. Pada tahun 1957, Taylor dan rekan-rekannya mampu menyimpulkan bahwa DNA kromosom pada Vicia faba dipisahkan dalam semikonservatif. Kesimpulan bahwa double helix direplikasi semikonservatif dalam kacang harus menunggu bukti selanjutnya menunjukkan bahwa setiap kromosom mengandung molekul DNA. Percobaan annalog kemudian dilakukan dengan beberapa eukariota lainnya, dan, dalam semua kasus, hasilnya menunjukkan bahwa replikasi terjadi secara semikonservatif.
Gambar 4: Bukti replikasi semikonservatif kromosom pada kacang, Vicia faba. Hasil yang diperoleh Taylor, Woods, dan Hughes (a, b) diprediksi oleh replikasi semikonservatif dari DNA (c).
Point penting dari Replikasi DNA :
a. Model konservatif, yaitu dua rantai DNA lama tetap tidak berubah, berfungsi sebagai cetakan untuk dua rantai DNA baru. Replikasi ini mempertahankan molekul dari DNA lama dan membuat molekul DNA baru.
b. Model semikonservatif, yaitu dua rantai DNA lama terpisah dan rantai baru disintesis dengan prinsip komplementasi pada masing-masing rantai DNA lama. Akhirnya dihasilkan dua rantai DNA baru yang masing-masing mengandung satu rantai cetakan molekul DNA lama dan satu rantai baru hasil sintesis.
c. c. Model dispersif, yaitu beberapa bagian dari kedua rantai DNA lama digunakan sebagai cetakan untuk sintesis rantai DNA baru. Oleh karena itu, hasil akhirnya
diperoleh rantai DNA lama dan baru yang tersebar pada rantai DNA lama dan baru. Replikasi ini menghasilkan dua molekul DNA lama dan DNA baru yang saling berselang-seling pada setiap untai.
ROLLING CIRCLE REPLICATION
Rolling circle replication adalah salah satu tipe dari replikasi DNA dan terjadi dalam replikasi pada organisme virus, bakteri, dan amfibi. Mekanisme rolling circle replication digunakan untuk :
1. menggandakan genom pada virus (menduplikasi genom mereka),
2. pada bakteri, mekanisme ini digunakan untuk mentransfer DNA dari sel donor ke sel resipie (penerima) selama satu jenis pertukaran genetik dan
3. pada amfibi, mekanisme ini terjadi saat oogenesis dalam rangka mengamplifikasi DNA ekstrakromosomal yang membawa kumpulan gen-gen RNA ribosomal.
Mekanisme rolling circle replication ini merupakan mekanisme replikasi DNA yang berbentuk sirkuler. Hal yang unik dalam rolling circle replication adalah bahwa DNA parental yang berbentuk sirkuler tetap utuh dan menggulung selama replikasi berlangsung dan berputar (roll/spin) sehingga mekanisme ini dinamakan demikian. DNA parental yang sirkuler ini menjadi template untuk sintesis unting komplemennya.
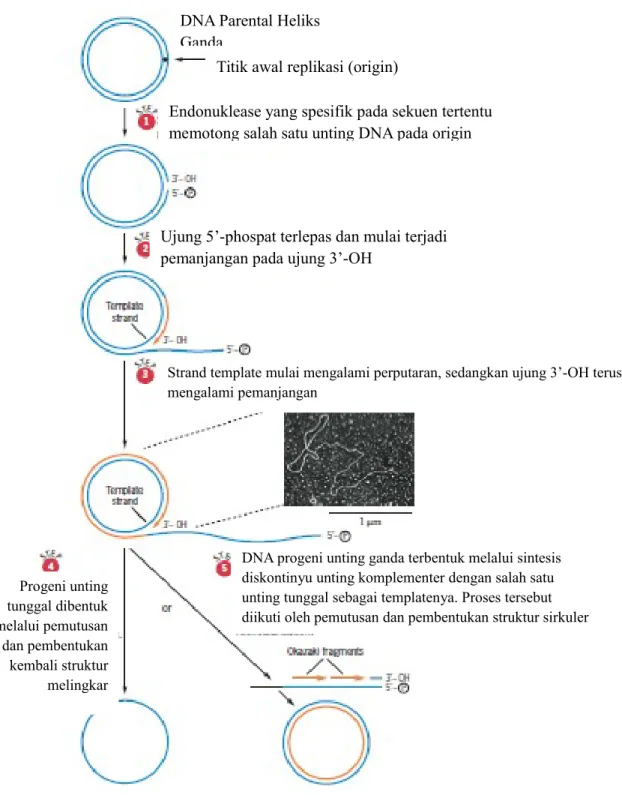
Rolling circle replication dimulai ketika enzim endonuklease yang spesifik terhadap sekuen tertentu memutus satu unting pada titik awal replikasi (origin). Aktivitas enzim endonuklease ini menghasilkan ujung 3’-OH dan ujung 5’-phospat. Ujung 5’-phospat kemudian dilepaskan dari lingkaran DNA, dan pada saat yang sama unting yang dijadikan sebagai template berputar pada sumbunya. Pemanjangan DNA terjadi pada ujung 3’-OH dari strand yang terputus.
Rolling circle replication dapat menghasilkan progeni (keturunan) DNA unting tunggal maupun unting ganda. Progeni unting tunggal yang sirkuler dapat dihasilkan melalui pemutusan spesifik tapak pada titik awal replikasi yang diikuti dengan pembentukan kembali struktur melingkar. Sedangkan, untuk menghasilkan molekul progeni yang terdiri dari unting ganda, DNA unting tunggal yang dihasilkan berperan sebagai template untuk pembentukan unting komplementernya. Pembentukan unting komplementer terjadi melalui pembentukan fragmen Okazaki (sintesis diskontinyu) diikuti dengan pemutusan dan pembentukan struktur melingkar. Proses terjadinya rolling circle replication dapat diilustrasikan pada gambar 1.
Gambar 5. Mekanisme Rolling circle replication (Sumber: Snustad dan Simmons, 2012)
DNA progeni unting ganda terbentuk melalui sintesis diskontinyu unting komplementer dengan salah satu unting tunggal sebagai templatenya. Proses tersebut diikuti oleh pemutusan dan pembentukan struktur sirkuler Progeni unting tunggal dibentuk melalui pemutusan dan pembentukan kembali struktur melingkar
Strand template mulai mengalami perputaran, sedangkan ujung 3’-OH terus mengalami pemanjangan
Ujung 5’-phospat terlepas dan mulai terjadi pemanjangan pada ujung 3’-OH
Endonuklease yang spesifik pada sekuen tertentu memotong salah satu unting DNA pada origin
Titik awal replikasi (origin) DNA Parental Heliks
REVERSE TRANSCRIPTION
Pada semua sel eukariotik hampir ditemukan elemen atau komponen genetik, yang dapat bertransposisi yang dikenal dengan transposon atau transposable elemen dan mekanisme perpindahan elemen ini tergantung pada reverse transcriptions yang mana merupakan perpindahan RNA menjadi DNA, siklus hidup beberapa virus tergantung pada Reverse transcription. Sel virus itu sendiri terdiri atas untaian RNA tunggal, ketika virus menginfeksi sel, untaian tunggal itu akan disalin ke dalam DNA beruntai ganda, sehingga informasi perpindahan genetik ini disebut retroviruses.
Retrovirus dari beberapa penelitian menyelediki penyebab jenis tumor pada ayam, kucing dan tikus. Pada setiap kasus produksi tumor selalu diakibatkan oleh RNA virus. Pemahaman ini berkembang sejak tahun 1970 saat david Baltimore, Howard Temin, dan Satoshi Mizutani menemukan sebuah reverse transcriptase yang memungkinkan virus itu menyalin RNA menjadi DNA. Penelitian ini tentang bagaimana proses transkripsi balik atau reverse transcription dimana urutan DNA berasal dari transkripsi balik, sehingaa dikatakan bahwa reverse transcription bertanggung jawab untuk mengisi gen dengan berbagai jenis urutan DNA termasuk retrovirus.
Contoh salah satu retrovirus yang kita kenal pada manusia yaitu penyakit AIDS yang sisebabkan oleh Human Immunodeficiency Virus (HIV). AIDS merupakan penyakit yang secara bertahap merusak kekebalan tubuh manusia yang mana pada tahap perkembangannya seseorang akan kehilangan kemampuan untuk melawan infeksi pathogen sehingga jika dibiarkan tanpa pengobatan penderita ini akan mati. AIDS ditularkan dari satu orang ke orang lain melalui cairan tubuh seperti darah atau air mani yang telah terkontaminasi dengan HIV. Gejala-gejala awal penyakit ini seperti terserang flu sehingga individu yang terinfeksi HIV akan mengalami nyeri, demam dan kelelahan, setelah beberapa minggu gejala-gejala ini mereda dan kesehatan tampaknya pulih, namun gejala-gejala ini dapat berlangsung selama beberapa tahun, virus terus berkembangbiak dan akan menyebar keseluruh tubuh, menargetkan sel-sel khusus yang mempunyai peranan menjaga homeostatis tubuh.
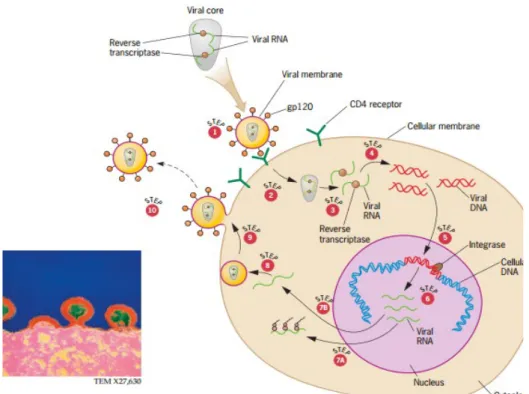
Berikut ini akan dijelaskan mengenai siklus hidup HIV (Gambar 1)
Gambar 6. Siklus hidup virus
a. Tahap 1: HIV berikatan/terkait dengan sel target melalui interaksi antara protein gp120 yang terdapat pada virus dan protein reseptor seluler CD4 pada sel target. b. Tahap 2: terjadi fusi yang memungkinkan inti virus masuk ke dalam sel target. c. Tahap 3: RNA yang terkait dengan protein dilepaskan dari inti virus.
d. Tahap 4: reverse transcriptase mengkatalisis sintesis double strand DNA virus dari single strand RNA virus dalam sitoplasma.
e. Tahap 5: Integrase mengkatalisis penyisipan DNA virus ke dalam DNA sel target dalam nukleus.
f. Tahap 6: RNA polimerase seluler mentranskripsi DNA virus menjadi RNA virus. g. Tahap 7a: Beberapa RNA virus berfungsi sebagai mRNA untuk sintesis protein virus. h. Tahap 7b: Beberapa RNA virus membentuk genom virus keturunan.
i. Tahap 8: Partikel virus keturunan dirakit di dekat membran sel.
j. Tahap 9: Partikel virus progeni dikeluarkan dari sel dengan membentuk budding. k. Tahap 10: Partikel virus bebas untuk menginfeksi sel-sel lain.
HIV ini merupakan virus yang sangat mematikan sehingga menjadi fokus dari banyak peneliti di dunia, salah satu hasil dari penelitian adalah pemahaman secara rinci bagaimana siklus hidup HIV. HIV memasuki sel inang memalui interaksi dengan protein receptor CD4 yang terletak di permukaan sel, interaksi ini dimediasi oleh glikoprotein yang biasa disebut gp120. Protein ini tertanam dalam lipid yang
mengelilingi virus, sesudah gp120 berikatan dengan receptor CD4, akan terjadi fusi virus dengan sel target. Di dalam sel, membran lipid dan mantel protein yang mengelilingi partikel virus akan terlepas, dan bahan-bahan dalam inti virus dilepaskan ke dalam sitoplasma sel. Inti ini berisi dua untai tunggal RNA-virus yang identik dan sejumlah kecil protein yang memfasilitasi replikasi genom, termasuk dua molekul reverse transcriptase dari virus, satu terikat untuk setiap untai RNA virus.
Reverse transcriptase HIV dan reverse transcriptase lain juga mengkonversi single strand RNA menjadi double strand DNA. Double strand DNA yang dihasilkan kemudian dimasukkan pada posisi acak dalam kromosom dari sel yang terinfeksi, pada dasarnya untuk mengisi genom sel dengan banyak salinan genom virus. Salinan ini kemudian disalin oleh RNA polimerase untuk menghasilkan sejumlah besar RNA virus, yang berfungsi untuk memediasi sintesis protein virus dan juga menyediakan RNA genomik untuk perakitan partikel virus baru. Partikel ini dikeluarkan dari sel melalui proses budding melalui membran sel sehingga dapat menginfeksi sel-sel lain melalui interaksi dengan reseptor CD4 pada permukaan sel target. Dengan cara ini, materi genetik HIV direplikasi dan disebarkan melalui sel-sel yang rentan.
Genom HIV panjang lebih dari 10 kilobasa, didalamnya ada beberapa gen, tiga gen disebut dengan gag, pol dan wnv, ditemukan di semua retrovirus lain, gen gag mengkode protein, gel pol mengkodekan enzim reverse transcriptase dan enzim lain yang disebut integrase yang mengkatalis penyisipan bentuk DNA genom HIV ke dalam kromosom, gen env mengkodekan glikoprotein.
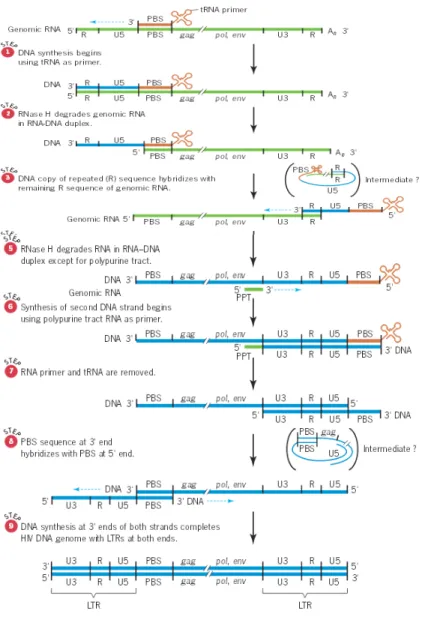
Gambar 7. Konversi RNA HIV kedalam DNA Beruntai Ganda
Proses replikasi genom HIV dikatalis oleh reverse transcriptase, dimulai dengan mensintesis untaian DNA single komplemen ke RNA untai tunggal dari genom virus, sintesis DNA dimulai dengan menggunakan tRNA sebagai primer yang menempel pada sekuen primer binding site yang berada di sebelah kiri pusat RNA HIV (Langkah 1) tRNA ini terbungkus terikat dengan PBS dalam inti HIV, setelah reverse transcriptase mengkatalis sintesis dari ujung 3 DNA virus ribonuklease H (RNase H) mendegradasi RNA genomik dalam duplex RNA-DNA (langkah 2). Degradasi ini membolak-balikkan pengulangan sekuen (R) untai DNA baru bebas untuk berhibridisasi dengan urutan R pada ujung 3 RNA HIV. Hasil akhirnya adalah
wilayah R dari untai DNA yang baru "melompat" dari 5 ujung RNA HIV ke ujung 3 dari RNA HIV (langkah 3). Reverse transcriptase selanjutnya memperluas salinan DNA dengan menggunakan 5 wilayah RNA HIV sebagai template (langkah 4).
Pada langkah 5, RNaseH mendegradasi semua RNA dalam duplex RNA-DNA kecuali daerah kecil yaitu Polypurine Track (PPT), yang sebagian besar terdiri dari purin adenin dan guanin. Polypurine Track ini digunakan sebagai primer untuk sintesis untai DNA yang kedua dari genom HIV (langkah 6). Setelah tRNA dan RNA genom hadir dalam duplex RNA-DNA dihapus (langkah 7), DNA kedua "melompat" terjadi selama PBS pada ujung 5 untai DNA kedua berhibridisasi dengan PBS komplemen pada ujung 5 untai DNA pertama (langkah 8). 3 -hydroxyl termini dari dua untai DNA yang kemudian digunakan sebagai primer sintesis DNA untuk menyelesaikan sintesis untai ganda DNA HIV (langkah 9). Konversi RNA virus ke DNA virus menghasilkan urutan tanda di kedua ujung molekul DNA. Urutan ini disebut long terminal repeats (LTRs), diperlukan untuk integrasi genom virus ke dalam DNA sel inang.
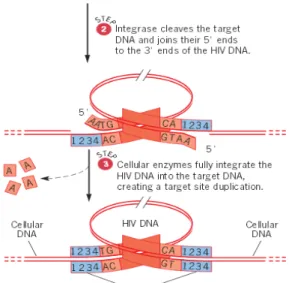
Gambar 8. Integrasi DNA HIV Beruntai Ganda ke dalam DNA Kromosom Sel Inang
Integrasi dari DNA virus dikatalisis oleh enzim integrase, yang memiliki aktivitas endonuklease. Enzim integrase akan memotong DNA HIV di dekat ujung 3' dari setiap untai atau pada kedua ujung LTRs (langkah 1). Ujung ini selanjutnya digunakan untuk integrase-mengkatalis ikatan fosfodiester dalam sekuens target dalam DNA sel inang. Proses ini menghasilkan formasi ikatan fosfodiester baru antara
ujung 3 DNA HIV dan 5 fosfat dalam DNA inang (langkah 2). Pada tahap akhir integrasi, enzim seluler sepenuhnya mengintegrasikan DNA HIV ke dalam DNA target, dan akan menciptakan situs duplikasi target (langkah 3). Genom HIV yang terintegrasi kemudian menjadi bagian permanen dari genom sel inang, replikasi sama seperti segmen lain dari DNA inang
Ketika fag T4 mengambil alih sel E. coli, ia menyediakan beberapa fungsi sendiri yang mengganti atau menambah fungsi inang. fag menempatkan sedikit ketergantungan pada fungsi ekspresi inang. Degradasi DNA inang penting dalam melepaskan nukleotida yang digunakan kembali dalam sintesis DNA fag. (DNA fag berbeda dalam komposisi dasar dari DNA sel dalam menggunakan hydroxymethylcytosine bukan sitosin adat.
Fungsi kode-fag yang bersangkutan dengan sintesis DNA dalam sel yang terinfeksi dapat diidentifikasi dengan mutasi yang menghambat produksi fag matang. fungsi fag penting diidentifikasi oleh mutasi yang mematikan bersyarat, yang jatuh ke dalam tiga kelas fenotipik:
Mereka di mana tidak ada sintesis DNA sama sekali mengidentifikasi gen yang produknya baik adalah komponen dari apparatus replikasi atau terlibat dalam penyediaan prekursor (terutama yang hydroxymethylcytosine).
Mereka yang timbulnya sintesis DNA tertunda prihatin dengan inisiasi replikasi. Mereka yang sintesis DNA dimulai tapi kemudian ditangkap termasuk fungsi
regulasi, ligase DNA, dan beberapa enzim yang bersangkutan dengan degradasi DNA inang.
Ada juga gen yang tidak penting berkaitan dengan replikasi, termasuk mereka yang terlibat dalam glikosilasi hydroxymethylcytosine dalam DNA.
Sintesis dari DNA T4 dikatalisis oleh agregat multienzim dirakit dari produk dari sekelompok kecil gen penting.
Gen 32 protein (GP32) adalah protein yang mengikat yang sangat kooperatif untai tunggal, yang dibutuhkan dalam jumlah stoikiometri. Ini adalah contoh pertama dari jenisnya yang akan ditandai. Geometri dari cabang replikasi T4 dapat secara khusus memerlukan protein fag-kode, karena E. coli SSB tidak dapat menggantikan. GP32 yang membentuk kompleks dengan polimerase DNA T4; Interaksi ini bisa menjadi penting dalam membangun cabang replikasi.
Sistem T4 menggunakan R A priming acara yang mirip dengan inangnya. Dengan DNA T4 singlestranded sebagai template, produk gen 41 dan 61 bertindak bersama-sama untuk mensintesis primer singkat. Perilaku mereka adalah analog dengan DnaB dan DnaG di E. coli. Gen 41 protein adalah mitra untuk DnaB. Ini
adalah helikase hexameric yang menggunakan hidrolisis GTP untuk memberikan energi untuk bersantai DNA. The P41 / P61 kompleks bergerak processively di 5'-3 'arah dalam sintesis untai tertinggal, secara berkala memulai fragmen Okazaki. protein lain, produk dari gen 59, beban P41 / P61 kompleks ke DNA; itu diperlukan untuk menggantikan protein P32 untuk memungkinkan helikase untuk berkumpul di DNA.
Gen 61 protein yang dibutuhkan dalam jumlah yang jauh lebih kecil daripada kebanyakan protein replikasi T4. Ada sedikitnya sepuluh salinan gp61 per sel. (Ini menghambat karakterisasinya. Hal ini diperlukan dalam jumlah kecil seperti yang awalnya itu terjawab sebagai komponen penting, karena cukup hadir sebagai kontaminan dari persiapan GP32!) Gene 61 protein memiliki aktivitas primase, yang analog dengan DnaG dari E. coli. Primase mengakui urutan template yang 3'-TTG- 5 'dan mensintesis primer pentaribonucleotide yang memiliki urutan umum pppApCpNpNpNp. Jika apparatus replikasi lengkap hadir, primer ini diperluas ke dalam rantai DNA.
Gen 43 DNA polymerase memiliki aktivitas sintetis yang biasa 5'-3 '. yang berhubungan dengan aktivitas proofreading 3'-5 'exonuclease. Ini mengkatalisis sintesis DNA dan menghapus primer. Ketika T4 DNA polimerase menggunakan DNA beruntai tunggal sebagai template, laju kemajuan tidak merata. enzim bergerak cepat melalui daerah untai tunggal, tetapi hasil jauh lebih lambat melalui daerah yang memiliki basis dipasangkan intrastrand struktur sekunder. Protein aksesori membantu polimerase DNA dalam melewati rintangan tersebut dan mempertahankan kecepatan.
Tiga protein lainnya disebut sebagai "protein aksesori polimerase." Mereka meningkatkan afinitas polimerase DNA untuk DNA. serta meningkatkan processivity dan kecepatan. Produk gen 45 adalah trimer yang bertindak sebagai penjepit geser. Struktur trimer yang mirip dengan yang ada pada E. coli dimer. dalam hal ini membentuk lingkaran di sekitar DNA yang memegang subunit DNA polymerase lebih erat pada template.
Produk gen 44 dan 62 membentuk kompleks ketat yang memiliki aktivitas ATPase. Mereka adalah setara dengan kompleks loader y8 penjepit, dan peran mereka adalah untuk memuat P45 ke DNA. Empat molekul ATP dihidrolisis dalam memuat penjepit P45 dan P43 DNA polymerase pada DNA. Struktur keseluruhan replisome adalah mirip dengan E. coli. Ini terdiri dari dua kompleks holoenzyme digabungkan,
salah satu mensintesis terkemuka untai dan lainnya sintesis untai tertinggal. Dalam hal ini, dimerisasi yang melibatkan interaksi langsung antara subunit polimerase DNA P43, dan P32 berperan dalam mengkoordinasikan tindakan dari dua unit DNA polimerase.
Sejauh ini kita telah berurusan dengan replikasi DNA hanya dalam hal perkembangan replikasi untuk Kebutuhan untuk fungsi lain ditunjukkan oleh DNA-delay dan DNA-penangkapan mutan. Tiga dari empat gen mutan DNA DNA-delay adalah 39, 52, dan 60, yang kode untuk tiga subunit dari T4 topoisomerase II. suatu kegiatan yang diperlukan untuk menghapus superkoil dalam template (lihat Bagian 19,13, topoisomerase Relax atau Perkenalkan superkoil dalam DNA). Peran penting dari enzim ini menunjukkan bahwa T4 DNA tidak tetap dalam bentuk linear, melainkan menjadi topologi dibatasi selama beberapa tahap replikasi. topoisomerase yang bisa diperlukan untuk memungkinkan rotasi DNA depan cabang replikasi.
Perbandingan apparatus T4 dengan apparatus E. coli menunjukkan bahwa replikasi DNA menimbulkan serangkaian masalah yang diselesaikan dengan cara-cara analog dalam sistem yang berbeda. sekarang kita bisa membandingkan enzimatik dan kegiatan struktural ditemukan di cabang replikasi di E. coli, T4, dan HeLa (manusia) sel. GAMBAR 18,26 merangkum fungsi dan memberikan mereka untuk protein individu. Kita bisa menafsirkan sifat dikenal replikasi protein kompleks dalam hal fungsi serupa yang melibatkan unwinding, priming, katalitik, dan reaksi penyegelan. Komponen setiap sistem berinteraksi dengan cara yang terbatas, seperti yang ditunjukkan oleh fakta bahwa fag T4 membutuhkan sendiri helikase, Primase, penjepit, dan sebagainya, dan oleh fakta bahwa protein bakteri tidak dapat menggantikan rekan-rekan fag mereka.
PERTANYAAN DAN JAWABAN A. Dewi Cahyaningrum
1. Mengapa rolling circle replication hanya dapat terjadi pada replikasi DNA yang berbentuk sirkuler? Jelaskan mekanismenya?
Jawaban:
Rolling circle replication merupakan mekanisme replikasi DNA yang umum terjadi pada DNA dengan bentuk sirkuler. Hal ini dapat terjadi karena pada DNA sirkuler saja lah yang dapat membentuk pemanjangan sembari DNA templatenya tetap utuh dan memutar. Mekanisme rolling circle replication dapat ditemukan pada replikasi genom virus, transfer DNA bakteri, dan amplifikasi gen-gen pengkode RNA ribosomal selama oogenesis amfibi.
Mekanismenya Rolling circle replication, yaitu:
Dalam replikasi DNA adalah kompleks, memerlukan partisipasi sejumlah besar protein.
Sintesis DNA adalah kontinyu pada untai keturunan yang sedang diperpanjang di keseluruhan arah 5 '→ 3', tetapi terputus pada strand yang berkembang di keseluruhan arah 3 '→ 5'.
Rantai DNA baru adalah diprakarsai oleh RNA primer pendek yang disintesis oleh DNA primase.
Sintesis DNA dikatalisis oleh enzim yang disebut DNA polimerase.
Semua DNA polimerase memerlukan untai primer, yang diperpanjang, dan untai cetakan, yang disalin.
Semua DNA polimerase memiliki syarat mutlak untuk 3'-OH bebas pada untai primer, dan semua sintesis DNA terjadi pada arah 5 'ke 3'.
Kegiatan 3'→ 5' exonuclease DNA polimerase mengoreksi untai baru yang lahir seperti yang merka sintesis, menghapus setiap nukleotida mispaired di ujung 3' untai primer.
Enzim dan protein pengikat DNA yang terlibat dalam replikasi merakit menjadi replisome pada setiap garpu replikasi dan bertindak dalam “konser” sebagai garpu yang bergerak sepanjang molekul DNA induk.
2. Di dalam penembentukan rolling circle replication ada enzim yang berperan dalam peristiwa tersebut. Enzim apakah yang bdrperan dalam peristiwa tersebut dan jelaskan carakerja enzim tersebut dalam perannya di rolling circle replication?
Jawaban :
Rolling circle replication dimulai ketika aktivitas spesifik enzim nuklease dari virus ФX174 itu, protein membelah rantai positif dari rantai induk RF pada daerah
ori dari replikasi. Aktifitas enzim ini sangat spesifik dengan memotong kromosom hanya pada satu sisi pada daerah ori. Enzim tersebut membentuk ujung 3’OH dan 5’Fosfat pada sisi yang dipotong itu pada untai positif dan untai negatifnya tetap utuh. Ujung 5’ dari untai positif tersebut terbuka dan terlepas sementara untai negatifnya berputar disekitar daerah aksis (sehingga namanya disebut rolling circle). Jadi enzim aktifasi yang pertama kali bekerja dan memiliki peran penting dalam proses rolling circle replication adalah enzim nuklease.
B. Frengky Neolaka
1. Jelaskan pengertian reverse transcription, retrovirus, bagaimana mekanismenya dan berikan salah satu contoh dari retrovirus!
Jawab :
Reverse transcription merupakan perpindahan RNA menjadi DNA pada sel eukariotik salah satunya adalah virus, siklus hidup beberapa virus tergantung pada Reverse transcription. Sel virus itu sendiri terdiri atas untaian RNA tunggal, ketika virus menginfeksi sel, untaian tunggal itu akan disalin ke dalam DNA beruntai ganda, sehingga informasi perpindahan genetik ini disebut retroviruses. Salah satu penelitian yang dilakukan oleh Howard Temin, dan Satoshi Mizutani tentang bagaimana proses transkripsi balik atau reverse transcription dimana urutan DNA berasal dari transkripsi balik, sehingaa dikatakan bahwa reverse transcription bertanggung jawab untuk mengisi gen dengan berbagai jenis urutan DNA termasuk retrovirus.
Salah satu contoh retrovirus yang kita kenal pada manusia yaitu penyakit AIDS yang sisebabkan oleh Human Immunodeficiency Virus (HIV). HIV memasuki sel inang memalui interaksi dengan protein receptor CD4 yang terletak di permukaan sel, interaksi ini dimediasi oleh glikoprotein yang biasa disebut gp120. Protein ini tertanam dalam lipid yang mengelilingi virus, sesudah gp120 berikatan dengan receptor CD4, akan terjadi fusi virus dengan sel target. Di dalam sel, membran lipid dan mantel protein yang mengelilingi partikel virus akan terlepas, dan bahan-bahan dalam inti virus dilepaskan ke dalam sitoplasma sel dan menyerang beberapa sel imun sehingga mengakibatkan seseorang kehilangan kekebalan tubuh dan mengakibatkan kematian.
2. Jelaskan perbedaan prinsip dari model replikasi semikonservatif, konservatif, dan dispersif? Teknik eksperimen apakah dalam pengamatannya?
Replikasi semikonservatif: replikasi yang terdapat pada untai DNA “parent” yang akan bertindak sebagai template yang akan dihubungkan oleh ikatan hidrogen agar dapat berpasangan dengan untai DNA yang baru terbentuk.
Replikasi konservatif: replikasi yang terjadi dengan mempertahankan DNA indukan, sedangkan untai baru dari DNA akan disintesis.
Replikasi dispersif: replikasi yang terjadi pada kedua segmen molekul untai DNA akan tetap dipertahankan dan digunakan sebagai template untuk mensintesis segmen yang komplemen yang setelah itu akan bergabung untuk menghasilkan keturunan untai DNA yang baru.
Molekul DNA dengan kepadatan yang berbeda dapat dipisahkan oleh equilibrium density-gradient centrifugation. Dengan menggunakan teknik inilah, ketiga model replikasi DNA melalui bentuk perubahan kepadatan pada DNA sel bakteri yang ditumbuhkan pada medium (15N) dan medium (14N).
C. Ari Ashari
1. Pada proses sintesis dari DNA T4 ada beberapa jenis protein yang digunakan salah satunya adalah “protein aksesori polimerase”. Deskripsikan secara singkat tentang peran dari protein ini?
Jawab
Peran dari protein aksesori polimerasi adalah untuk meningkatkan afinitas polimerase DNA untuk DNA. serta meningkatkan processivity dan kecepatan. Kemudian produk gen 45 adalah trimer yang bertindak sebagai penjepit geser. Struktur trimer yang mirip dengan yang ada pada E. coli dimer. dalam hal ini membentuk lingkaran di sekitar DNA yang memegang subunit DNA polymerase lebih erat pada template.
2. Pada proses replikasi bakteri FAG T4, Gen 43 DNA polymerase yang biasa memiliki aktivitas sintetis 5'-3 ' yang mana ini berhubungan dengan aktivitas proofreading 3'-5 'exonuclease. Peran dari Gen tersebut apa?
Jawab:
Peran dari Gen 43 DNA polymerase ini adalah mengkatalisis sintesis DNA dan menghapus primer. Ketika DNA T4 polimerase menggunakan DNA beruntai tunggal sebagai template, laju kemajuan tidak merata. enzim bergerak cepat melalui daerah untai tunggal, tetapi hasil jauh lebih lambat melalui daerah yang memiliki basis dipasangkan intrastrand struktur sekunder. Protein aksesori membantu polimerase DNA dalam melewati rintangan tersebut dan mempertahankan kecepatan
DAFTAR RUJUKAN
Snustad, D. P. dan Simmons, M. J. 2012. Principles of Genetics Sixth Edition. New York: John Wiley & Sons, Inc.
Densmore, L. 2007. Modes of Replication 2. (Online),
(http://www.faculty.biol.ttu.edu/densmore/MolBio/Mol%20Bio%2007/Molecular %20lect%2011.07.pdf), diakses 17 February 2016.