fnaialah
llrniab
rssN,
o85+
-
0728
AGR.IPLf]S
Azhqr Bafadat: pEMBIAYAAITI DEFISIT DAI{ KEBERLAI{JUTAI{ FISIGL
NIuTdlanT
K.
z PENGARUH UNGI{.JNGAI{ BISNIS EKSTERNAL DAI\ INTERNAL TERHADAP KINEFLIAUSAI{A KECIL (Kasr-rs Usaha Kecil Sepatu Kulit di Propinsi Jawa Barat)
Ambo AKo: GRMING ADAPTABIUTY OF BEEF CATTLE ON THE DWARF NAPIERGM (Penn isetum
purpureum Schumach) PASTURE
AbdT
:
EFESIENSI PEMAI{FAATAN FAKTOR PRODUKSI USAHATANI PADI LADAFIG PETANITRANSMGRAI{ DI I{ECAIVL{TAN TIKEP IGBUPATEN MUNA
Asussatim,
SahtoGtntrns
danLo
Ode
Soboruddln : PEWILAYAFIAN KOMODITAS PERTAI{IANBERDASARKAI\ ZONA AGROEKOLOGI DI KECAI\4{TAN POLEAI{G SULAWESI TENGGARA
Humsoh, Darnas Dans
dan
Nlarthen B.NI.Malole:
PERAII PAIGI\ ALAMI DALAM PENULARAI{ White Spot SyndromeVirusPADA BENUR UDAI.IG WINDU (Penaeus monodon Fabr.) SEBUAI-{ K,{llAI\fAWAL
H.
Gusti R. Ssdi mantqra: INDUKSI IGLUS DAI\ ORGAI\OGENESIS JER{JK KEPROK SIOMPU PADAMEDIUM MS DENGAN KOMBINASI
AUI$IN
DAI{ SNOKININLs
Ode
Safuan, RoedhyPoerwsnto,
AnasD.
Susllq,
Soblr,dsn
BylcsonSltumorang:
MINUS-ONE TEST KESUBURAI{ TANAFI INCEPTISOL, ULTISOL, DAI{ AI{DISOL LINTUK TANAIVIIAN NENAS
Ls
lvluhurls,
Dtdy Sopandte, Latifah Koslm Dsrusmon : BEBERAPA PEUBAH BIOKIMIA TERKAITRESPIRASI PADA KEDELAI (Glycine mox
L.
Merrill) TOLERAIIDAI\
PEKA INTENSITAS CAHAYARENDAFI
La Ode
Afa : STUDI MATRICONDITIO/VING PADA BENIH KACAI{G TANAFI (Arachis hypogaeo L.) Suoib,WoerJono Nlangoendtdlolo, Nllrzoltnorn, PD.N., donArl lndrlanto
: POPULASI MIKROSPORAUNIN{.JKLEAT BERDASARIGI\ LETAKNYA PADA MALAI TIGA KLONTEBU (SaccgaTum spp.) SEBAGAI
NORUqSI
AWAL BAGI PEMULIAAI\ HAPLOID SECARA IN VITROLa
Rtonda,Lo Ode Arlef,
DJukrana Wahsb, Thamrln danSuto
: I(A.llAI'tr RESPON KONSUMENTERHADAP SIRUP METE PRODUKSI UNIT USAI-IA JASA
DAI\
INDUSTRI FAKUNAS PERTAI\IAI{UNIVERSITAS HALUOLEO.
Soedtmsn : ESSENTIAL FEATURE AI\D OPERATION OF SAI{CHOKU (DIRECT TRAI\SACTION) IN
JAPAI{ S CONSUMER COOPERATIVES
GAK Suturiutt,
Wtdodo, Sudarsonodsn S
flyos
: EFEKTIVITAS AGENS BIOKONTROL UNTUKMENINGKATKAN PERTUMBUHAN
DAN
HASIL CABAI SERTA MENGHNDALIKAI{ PENYAKITDAFTAR
ISI
Halamon
PEMBIAYAAN DEFISI'T DAN KEBERLANJUTAN FISKAL
Azhar
Balodal
I-7
l,tiNGAlttJlt t,INGKUNcAN BtsNts IiKs't'tiRNAt, t)AN tN',t't,:RNAt, 't'ERllADAt' KINERJA TJSAHA KECIL (Kasus Usaha Kecil Sepatu Kulit rli Propinsi Jawa llarat)
Murtljani K.
GRAZING ADAPTABILITY OF BEEF CATTLE ON 'I'I{E DWARF NAPIERGRA
(Pennisetu m pu rpu reu m Schumach) PASTURE Ambo Ako
EFISIENSI PE]VIANFAATAN FAKTOR PRODUKSI USAIIATANI PADI LADANG
PETANI'I'IIANSM IG RAN DI KECAMATAN TI K EP KA I}t J PATEN i\I I INi\
,4bdi
PEWILAYAHAN KOMODITAS PERTANIAN
BERDASAIIKAN ZONAAGROBKOLOGI DI KECAMA'TAN POLEANG SI.]I,AW[,SI'I'F],NGGAIIA
Agussalim, Sohta Ginting dan La Ode
Soburuddin
2g - 36PERAN PAKAN ALAMT DALAM PENUI,ARAN White Spot Syndrome
ltrzs
pADABENTJR [IDANG 1VINDU (Penaeus monodon Fabr.) SEBt.iAtt K,\.ttAN ,\\\,At,
Hamsoh, Dsrnas Dana dan Msrthen B.M. Malole 37 -43
INDUKSI KALUS DAN OIIGANOGENESIS .lERtlK KIPROK
Slor\tpt]
p.A,DAIVIEDIUM MS DENGAN KOMI]INASI ATIKSIN DAN SITOKININ
IL Gusti R Sodimantara 44-49
N{INUS.ONE'I'EST KESI,JI}I.JRAN TANAII INCEPTISOI,, I.JL'rISOI., DAN ANI)ISOI,
TINTI.]K TANAIVIAN NENAS
La Ode Safuu, Roeilhy Poerwanto, Anas D, Susila, Sobir, tlan Rykson
Situnorang.,..
50 - 5{t BEBERAPA PEUBAH BIOKIMIA TEIIKAIT RESPIRASI Pr\DA KED!.r.,\r (Gr.ycine maxL. l\lerrill) TOLERAN DAN PtiKA IN'l'ltNStl'AS CAilA\,,\ RFtNt)Alt
I.a Muhuria, DitlySopondie, Lutitoh Kosim Darusmun
...._...
59 _70sruDl MATRICQNDITIQNING PADA llENttt KACANG 1'/\NAtI (Aruc'his hl,pogaeaL.) La ode
Afa
7r
-.,g PoPIILASI NttKRosPoRA UNINUKt,EA'|" BERDASARKAN l,u'I',\KNyA pADAMALAI
rlGA
KLON TEBU (sacclarum spp.) s[BAGAt tNt-oRl\{Asl AWAL BAGIPEMULIAAN HAPLOID SECARA IN VITRO
8-t4
r5-20
)l _)1
Suaib, ll/oerjono Mongoendidjojo, Mirzawan, P.D.N., dan Ari Intlrionto...
KAJIAN ROSPON KONSTIMIiN I'ERIIADAP SIRTIP ME'I'E PRODIIKSI TINIT TISAIIA JASA DAN INDUSTRI FAKIJLTAS PEIITANIAN TJNIVERSITAS IIALTIOLEO
La Rionda, La Ode Ariel, Djukrana llahob, Thunrin dan Suto
ESSENTIAL FEATURES
AND
OPERATIONOF
SANCHOKU (DIRECTTRANSACTTON) tN JApAN'S CONSUt\tER COOpERATTVES Saedinan
EFEKTIVITAS AGENS BIOKONTROL IjNTTJK IVTENINGKATKAN PERTTIIIIBTJIIAN
DAN HASIL
CABAI
SERTA MENGENDAI,IKAN PF]NYAKIT ANI-RAKNOSA DIRUMAI{ KACA
GAK Sutariati, llidodo, gudarsono ilan S
llyas
103 _ Ill
80-88
POPULASI MIKROSPORA UNINUKLEAT BERDASARKAN
LETAKNYA
PADAMALAI
TIGA
KLON TEBU (Sacchorum spp.) SEBAGAI INFORMASIAWAL
BAGI PEMULIAANHAPLOID SECARA
IN
VITROOteh: Suaibt, ll/oerjono Mangoendidjojot, Mirzowan, P.D.N., dan Ari Indriantoa.
ABSTRACT
An experiment to study the sugarcane panicles containing more than 50 7o of uninucleate microspore development based on the two morphological characteristics of the three sugarcane clones namely
:
52OC2,52OC4 and POJ3025 was conducted
in
Tissue Culture Laboratory at Biology Faculty, Gadjah MadaUniversity, Yogyakarta, since March until May 2005. Morphological characters of both kinds of panicles i.e. unsheated- and sheated-flowers from sheat flag leafwere observed. Mean, percentage, and standard deviation
from the mean values of the three different stages of microspore development e.g early-uninucleate and late-uninucleate developments, and pollen grains were statistically used in this calculation. All data percentages were analyzed by variance analysis through General Linier Model Procedure, and comparison between means based on microspore position in the three different parts of the panicles was calculated by Least Square
Difference method. Comparisons between the two different panicles characteristics in accordance with the
proportion of the three difference microspore development, however, were analyzed by T-student procedure.
All calculation were done by using SAS program of computer statistics package. Result of the research shows
that:
(l)
the unsheated panicles were contained more than 90 %o of uninucleate microspore development; (2) the sheated panicles tend to be in high proportion of binucleate microspore development and pollen grains,and (3) the more away of spikclets or anthers positioned in the panicle or subpanicle, the more number or percentage of uninucleate microspores development were tend to be gradually decreased.
Key wor : panicle ntorpholog', micro,spore, uninucleate, pollen grains, haploid breeding
PENDAHULUAN
Pemuliaan tanaman secara
in
vitrodiilhami oleh tslakeslee dan koleganya yang menemukan tanaman haploid pada tanaman
Datura
stramonium padaawal
abad
XX(Blakeslee et
al.,
1922). Sampai pada tahun 1983, telah berhasil diperoleh tanaman haploid dan haploid ganda melalui kultur antera danmikrospora secara
in
vitro
pada 247 spesies dari 88 genera dan 34famili
(Maheswari etal.,
1983). Bahkan, tahun 2004 telah dilepaslebih dari 60 kultivar unggul berbagai spesies
hasil kultur
antera dan mikrospora (Cistueet
al.,
2004).
Keberhasilan mendapatkantanaman haploid dan atau haploid ganda" baik melalui kultur antera (anther culture) maupun
melalui
kultur
mikrospora
(microsporeculture),
sangat ditentukanoleh
beberapafaktor.
Faktor-faktor dimaksud adalah:(l)
genotipe tanarnan donor.
(2)
kondisi
per-tumbuhan tanaman donor,
(3)
tahapper-kembangan mikrospora,
(4)
praperlaku-an,dan (5) medium dan kondisi kultur (Kush
&
Virmani,
1996; Palmerdan Keller,
1997; Bhojwani&
Bhatnagar, 1999).Tahap perkembangan mikrospora
se-bagai salah satu faktor yang menentukan arah
diferensiasi mikrospora akan menghasilkan embrio dan atau kalus. Terbentuknya embrio akan menghasilkan tanaman hijau dan leng-kap, sedangkan terbentuknya kalus cenderung
akan - menghasilkan tanaman
bulai
dalamfrekuensi yang tinggi.
Ini
berarti diperlukan informasi yang tepat mengenaiciri
morfologimalai yang mengandung mikrospora dalam frekuensi yang
tinggi
pada tahap uninukleatatau berinti satu. Pada tanaman Barley (Kasha
et a\,,2001) dan Wheat (Liu et a\.,2002) akan
diperoleh
embriogenik
mikrospora
danr)
StafPengajar pada Fakultas Pertanian Universitus tlaltroleo, Kendari
2)
Guru Bcsar llmu Pcmuliaan Tanaman pada Fakuitas PertaniarVSekolah Pascasarjana Universitas Gadjah Mada, Yogyakarta
r) Direktur Pusat Pcnclitian dan Perkebunan Gula lndonesia ( P3Gl), Pasuruan Jawa timar
n)
tanaman haploid ganda dalam .iumlah yang banyak apabila mikrospora dikulturkan pada
tahap
uninukleat tengah (mid-uninucleate)hingga
uninukleatakhir
(late-uninucleate). Sementaraitu,
produksi tanaman haploid ganda (doubled haploid) yang sangat tinggi pada tanaman Barley dapat dicapai apabila rnikrospora dikulturkan ketika berada padatahap uninukleat awal hingga uninukleat akhir
(Li &
Devaux, 2003). Pada tanaman Brassicanapus (Cordewener et a1.,1996), untuk meng-hasilkan embrio
yang
banyak, mikrospora harus berada pada tahapakhir
mikrosporaberinti satu
hingga pada tahap
awal mikrosporaberinti
dua
(early-binucleate).Pada tanaman
tebu, tahap
perkembanganmikrospora melalui kultur
antera
mernberi-kan hasil yang lebih baik ketika mikrospora berada pada tahap sesaat setelah stadium selinduk tepung sari yaitu stadium dua sel (diad)
hingga stadium empat sel (tetrad) (Chen et al.,
1979). Sementara iru, Fitch dan Moore (19g3)
mengemukakan bahwa tahap akhir mikrospora
berinti satu hingga
berinti dua
merupakantahap
yang paling
produktif
padaembriogenesis mikro-spora tanaman tebu liar, Saccharum sponlaneum
L.,
melalui kulturantera secara in vitro.
Tahap-tahap perkembangan
micro-spora
di
atas mempunyai hubungan yangsignifikan
denganmorfologi
dan
panjangwaktu
setelahinisiasi
bungaatau
malai.Namun demikian, penggunaan renrang umur setelah inisiasi bunga, kurang diaplikasikan karena sulitnya menen-tukan saat mula-mula terjadinya inisiasi bunga atau malai. Akarr
tetapi, penggunaan kriteria rnorfologi bunga
berupa ukuran panjang bunga atau malai, atau
ukuran pemunculan bunga atau pemunculan malai di atas daun bendera merupakan kriteria
yang umum dipakai pada banyak tanaman
gramineae. Pada tanaman rumput makanan
ternak tahunan, Ryegrass (Lolium perenne L.), pemun-culan seperempat bagian malai di atas
daun
bendera
menghasilkan
populasimikrospora terbanyak pada fase uninucleate (Suaib
et
al.,
1997). Sementaraitu,
untuk tanaman padi ladang beberapa varitas lokal asal Kendari menunjukkan balrwa malai yang8l
muncul di atas gulungan kelopak dau bendera setinggi kurang
dari
3,0
cm
menghasilkan mikrospora pada fase mikrospora berinti satudalam jumlah yang lebih banyak (> 60 %) dari
total
mikrospora dan tepung sari matang di dalam antera (Suaib, 2000). Indrianto (2003)menyatakan bahwa pada tanaman Wheat,
rnorfologi
pertumbuhanmalai yang
me-ngandung mikrospora
yang
'sesuai untuk kultrrrin
vitro adalah ketika masihdi
dalarnbungkusan kelopak daun bendera atau belum ada bulir yang muncul
di
atas daun bendera.Khusus pada tanaman tebu
liar
Saccharumspontaneum L., dilaporkan bahwa mikospora yang sesuai bagi kultur antera didapat ketika malai masih berada di dalam bungkusan daun
bendera (Fitch
&
Moore, 1983; Moore et al..r e89).
Belum ada informasi mengenai ciri
morfologi malai tanaman tebu
klon
hibrida yang rnengandung populasi mikrospora padatahap yang sesuai
bagi kultur
mikrospora. Oleh karena itu, tulisan ini membahas ciri-cirimorfologi malai
tiga klon
hibrida tanamantebu yang mengandung mikrospora lebih dari
50 % pada tahap uninucleate.
METODE PENELITIAN
Penanaman Tanaman Donor
Potongan batang (bagal, seilsz) tebu klon:
(l)
52OC2, (2) 52QC4, dan (3) pOJ3025sepanjang dua buku yang digunakan dalam penelitian
ini,
ditanam pada4
Juli
2004 diRumah Kawat Fakultas Pertanian Universitas
Cadjah Mada, Kampus
Bulak
Sumur,Y\)g)akarta.
Bagal
ditanam dengan posisihorizontal
di
dalam juring pada kedalaman*
25 crn. Panjang juring adalah 5 m dan jarak
antara
juring
adalahI
m,
sehingga luaskeseluruhdn penanaman
tebu
sumbermikrospora adalah
25
m2. Setelah tumbuh, tanaman dipelihara hingga berbunga dalambentuk
pemberianpupuk
N, p,
dan
K, berturut-turut berupa Urea, Sp36, dan KCI dengan dosis berturut-turut 200, 100, dan 100kg
per hektar atau berturut-turut 0,10; 0,05,dan 0,05 kg per 5 m panjang juring. Aplikasi
82
ketiga pupuk
di
atas seluruhnya dilakukanpada
umur
empat minggu
setelah tanam (MSr).Selain
pemupukan,juga
dilakukan pengendaliangulma
dan
pengaturanpe-nyiraman, serta penggemburan tanah di sekitar perakaran tanaman. Tanaman mulai berbunga
pada umur 37,38 dan 39 MST berturut-turut
bagi
klon
52OC2, 52OC4,dan
POJ3025.Sementara
itu,
isolasi antera dan mikrospora dimulai pada 39 MST.Pemanenan Malai
Tanaman tebu yang telah berbunga,
baik malai masih terbungkus di dalarn kelopak
dar"rn bendera maupun yang telah rnuncul di
atas daun bender4 dipotong pada 2-3 ruas di
bawah pangkal
malai.
Malai
tersebutdimasukkan ke dalam tabung gelas yang berisi
air
kran, lalu dibungkus dengan kertas koranyang
lembap
dan
segera
dibawa
dandimasukkan ke dalam lemari pendingin pada
suhu 5oC. Ciri morfologi malai yang diamati, yakni berdasarkan:
(l)
malai masih terbungkusdi dalam kelopak daun bendera, dan (2) malai telah muncul di atas daun bendera. Setiap ciri
morfologi pertumbuhan
malai
diamati tigacabang
malai dan
masing-masing cabangmalai diamati tiga bulir, sedangkan tiap bulir mengandung
tiga
antera
sebagai sumbermikrospora. Setiap
unit
pengamatan diamatiminimal
300
mikrospora,yakni
masing-rnasing satu malai yang belum muncul bagi
klon
52OC2, 52OC4, POJ3025, satu malaiyang telah membuka bagi klon 52OC4, dua
malai
yang
telah
membukabagi
klon POJ3025, dan tiga malai yang telah membukabagi
klon
52OC2. Setiap perlakuan yangberulangan dilakukan pada waktu dan dengan
malai
yang
berbeda.Dengan
demikian, seluruhnyadiamati
sembilanmalai
dalampengamatan ini.
Isolasi
dan
Pemeriksaan
Tahap
Per-kembangan Mikrospora
Mikrospora diisolasi dengan
cara
:antera dikeluarkan
dari
dalam lemma danpalea bulir dengan sepasang pinset, kemudian diletakkan di atas deckglass yang sebelumnya telah ditetesi dengan larutan
3
0% sukrosa.Dengan sepasang
ujung
pinset, mikrospora dikeluarkandari
dalam antera dengan caramembelah antera hingga mikrospora keluar
dari
dalam antera. Kotoran berupa sisa-sisajaringan dinding
antera, kemudian ditutup dengan coverslip.
Popu-lasi mikrospora diatas deckgloss tersebut siap diamati di bawah
mikroskop biasa.
Penentuan
tahap
perkembanganmikrospora di atas menggunakan patokan ciri
perkem-bangan mikrospora
pada
tanamangandum
(Lu
&
Kuo,
1984), pada tanamanBarley (Kasha et a\.,2001), dan pada tanaman
padi (Zhang et a|.,2005). Adapun tahap-tahap
perkembangaq
mikrospora
yang
diamatiadalah:
(a)
uninukleat awal,(b)
uninukleatakhir,
dan(c)
tepungsari.
Ciri
morfologi ketiga tahap perkembangan mikrospora, antaralain: uninukleat awal, inti terletak pada bagian
tepi dekat dinding mikrospora dan dekat porus
tumbuh (germ pore) yang membagi dua dari
seperempat
bagian lingkaran
mikrospora; uninttkleat akhir, inti terletak pada bagian tepi dekatdinding
mikrosporadan
berlawanandengan porus tumbuh atau mem-bagi dua
bagian lingkaran mikraspora, dan tepung sari, mikrospora yang telah mengalami pembelahan
mitosis pertama, inti berjumlah dua atau lebih
dan
telah
mengandungbutir-butir
pati
di dalam sitoplasma. Setiapunit
pengamatandiamati minimal 300 individu mikrospora. Variabel Penelitian
Variabel penelitian adalah persentase
tahap-tahap perkembangan mikrospora dan
dihitung dengan formula:
Persen Uninukleat Awal (o/o UA; =
I
uua
tffi-;Znx
roo%dengan
IVUa =
jumlah
mikrosporauninukleat awal;
IMUp:
jumlah mikosporauninukleat akhir;
ITS
= jumlah tepung sariPerhitungan
tahap
perkembanganmikrospora
lainnya
yakni
persentaseuninukleat akhir, dan tepung sari, mengikuti
cara yang
sama dengancara
perhitunganuninukleat awal di atas.
Analisis Data
Data yang
diperoleh
kemudiandihitung
nilai
rerata,
persentase, dansimpangan baku-nya.
Data
persentase di-analisis secara anova melalui prosedur modellinier
umum (PROC QLM, General Linear Model Procedure), sedangkan korelasi antarapanjang pemunculan malai terhadap persen-tase tahap-tahap perkembangan mikrospora
dianalisis
dengan
PROC CORR.
Penr-bandingan rerata respon kedua morfblogi
malai yang
digunakan pada penelitian ini dilakLrkan denganuji+
(Student's T-te,tt)sesuai prosedur
Steel
dan Torrie
( I 98 I )dengan PROC TTEST. Semua perhitungan di
atas menggunakan paket statistika SAS (SAS lnstitute lnq., 1989-l 996).
83
HASIL DAN PEMBAHASAN Paling sedikit, telah diarnati 218.700
butir
rnikrospora dan tepungsari
dari 729antera,243 bulir,
Sl
bagian cabang malai,27 cabang malai, dan sernbilan malai pada duaklon
tebu Saccharumffieinarum
dan satuklon hibrida kompleks dalam menentukan
ciri-ciri
rnorfologi
pertumbuhanmalai
yangmengandung mikrospora berinti satu lebih dari
50
%.
Sernbilan malai bahan penganlatanberasal dari dua klon Saccharum officinarum rnasing-rrrasing klorr 52OC2 dan 52OC4, dan satu
klon
hibrida komleksyakni
POJ3025,sedangkan
.27
cabangmalai
berasal darirnasing-masing
tiga
bagian,yakni
:
u.jr-rng,tengah
dan
pangkal malai. Masing-rnasing cabang malai nrerupakan sumbertiga
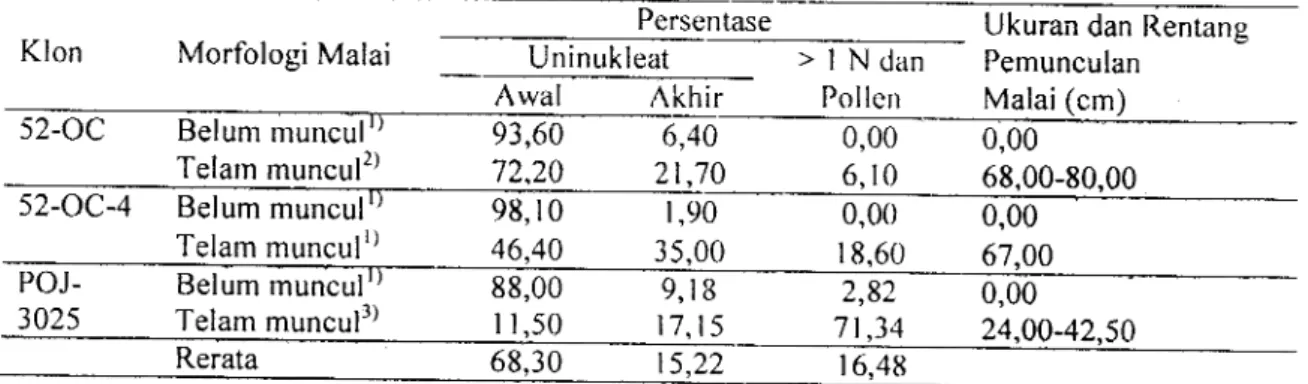
bulir dimana setiap bulir rnengandung tiga antera.Tabel
l.
Morfblogi dan rerata pcrsentase nrikrospora rncnurut tahap pcrke rnbangannya, serta ukuraupemunculan malai dua klon tebu Saccharurn olJicinurum (L.) dan satu klon tebu hibriiJa.
Persentase Ukuran dan Rentang
Klon MorfologiMalai Uninukleat
>lNdan
PemunculanAwal
Akhir
Pollen
Malai (cm)52-OC Belum rnuncul
0,00
0,00Telam muncul2)
52-OC-4
Belum munculr)-
9&10
98,r0 t-9t.9093,60 72.20 6,40 21.70 6, l0 68,00-80,00 Telarn munculr) 46,40 35,00 0,00 18,60 0,00 67,00 POJ-3025 Belum muncul 88,00 2,82 0,00 Telam muncul3) 16,49 Keterangan : ') Satu malai, 2)
Tiga malai, dan r) Dua malai; > lN = bi-, tri-, multi-nukleat dan tepung sari
Tabel
I
menunjukkan bahwa semua malai yang belum memunculkan bunganya,tidak
mengandung mikrospora yang berinti lebih dari satu(>lN)
atau tepung sari Qtoilen).Sedangkan
malai yang telah
muncul bunganya, seluruhnyatelah
mengandungketiga
macam
tahap
perkembanganmikrospora dengan persentase yang
berbeda-beda. Adanya perbedaan persentase
masing-masing tahap perkembangan mikrospora di-sebabkan
oleh
adanya perbedaan:(l)
klon yang diamati,(2)
panjang malai seluruhnya,(3)
ukuran
pemunculanbunga,
dan
(4) proporsi bagian bunga yang belum dan telahmuncul. Pengamatan keempat
ciri
morfologi malai di atas disajikan pada Tabel 2.84
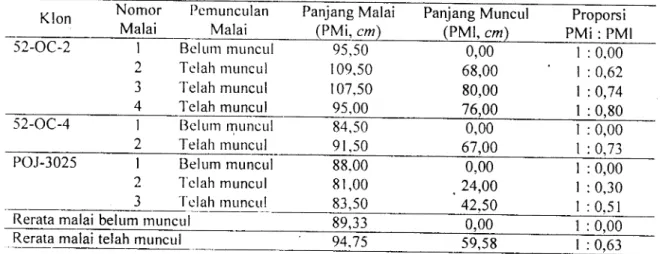
Tabel
2.
Ciri
morfologi malai berdasarkan pemunculan rnalai, panjang malai seluruhnya (cm), panjang pemunculan malai (crn), dan proporsi panjang malai seluruhnya dengan panjang malai yang telah muncul dua klon tebu Sacchorum oficinarum (L.)iun
rutu klon tebuhibrida kompleks
Klon Nomor
Malai
Pcmunculan
Malai PML cm
Panjang Malai Panjang Muncul
52-OC-2 Belum muncul
Telah muncul Telah muncul Telah muncul I 2 J 4 95,50 r09,s0 107,50 95,00 0,00 68,00 80,00 76,00
I
: 0.00I
: 0,62I
:0,74I
: 0,8052-OC-4 Belum muncul
Telah muncul I 2 84,50 91,50 0,00 61.00
I
: 0,00I
: 0,73 POJ-3025 Belum munculTelah muncul 'T'clah rnuncLr!
I
: 0,00I
: 0,30I
: 0.51 I 2 J 88.00 8 r,00 83,50 0,00 24,00 42.50Rerata malai belum rnuncul 89.33 0,00
1:0.
: 0,00 Rerata malai telah muncul 94.t5 59,59I
: 0,63'fabel
2
menr.rnjukkan bahwanralai yang belum memunculkan bunganya
mem-punyai panjang rata-rata 89,J3 crn, sedangkan
malai yang
telah
memunculkan bunganyamempunyai panjang rata-rata 94,75
cm.
lni berarti bahwa rerata panjang malai tanamantebu
obyek pengamatan adalah 92,04 cm.Tabel
3
juga
menunjukkan bahwa apabila bunga telah muncul sepanjang63
o/o atau 59,58 cm, perkembangan mikrospora satu intimenjadi
mikrosporalebih satu
inti
akanmencapai sebesar 24,71 yo (Tabel
l).
Dengan demikian, pada tanamantebu
masih akandiperoleh mikrospora
berinti
saru mskipun bunganya telah muncul lebih dari separuhnyaasalkan belum terjadi antesis yang ditandai dengan perubahan warna antera dari berwarna
hijau
muda
menjadi berwarnaungu
ataucokelat muda, atau rambut sutera (silk) yang terdapat pada pangkal bulir belum merekair.
Apabila panjang pemunculan malai
dikorelasil<an dengan persentase tahap per-kembangan
mikrospora,
nampak
bahwahampir selnua variabel berkorelasi negatif kecuali antara variabel panjang pemunculan rnalai dengan tahap mikrospora uninukleat
akhir yang bernilai
positif.
Demikian pula,signifikansi
tidak
berarti
antara
panjangpemunculan malai dengan ketiga tahap
per-kembangan mikrospora.
Hanya
persentasetahap uninukleat awal dan persen-tase tahap
tepung sari yang berkorelasi sangat erat dan
signifikan meskipun bernilai negatiq
sedang-kan
uninukleatawal
dan
uninukleat akhir berkorelasi secara tidak signifikan (Tabel 3).Dengan
demikian, apabila
rnalai
telah memunculkan bunganya maka peluang malai akan mengandung mikrospora berada pada tahap berinti lebih dari satu hingga pada tahaptepung sari menjadi semakin besar. Semakin
panjang pemunculan bunga akan semakin
tinggi
persentase mikrospora berada padatahap
lebih dari
satuinti
khususnya tahaptepung sari sehingga jumlah mikrospora yang berpeluang untuk memasuki
jalur
sporofitikakan semakin berkurang.
'_
Tabel 3. Korelasi antara
rnikrospora tiga
85
panjang pemunculan malai dcngan persentase tiga tahap perkem-bangan
klon tebu.
Uninukleat awal Uninukleat akhir Tepung sari
Pemunculan malai - 0,17009 ts 0,6617 0,54508 ts 0,t291 - 0,04413 ts 0,9102 Uninukleat awal - 0,25280 ts 0,51r6 - 0,92197 ss 0,0004 Uninukleat akhir - 0,141 60 ts 0,7 r 63
Keterangan
:
Nilai-nilai yang dicetak biasa adalah koefisien korelasi; Nilai-nilai yang dicetak miringadalah peluang > lRl terhadap Ho:Rho:0(o = 0,05) ts = riclak signifikan,
ss = s-angat signifikan
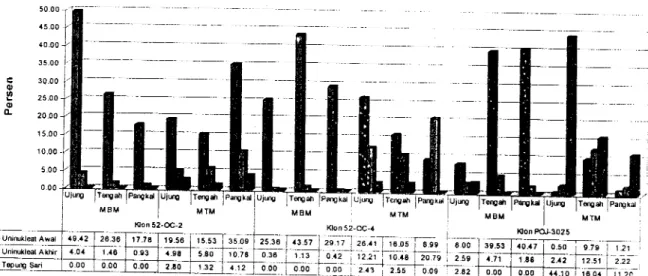
Gambar 1 menunjukkan bahwa malai
yang
belum memunculkan bunganya padaketiga
klon
yang
diamati
mengandungmikrospora pada tahap
berinti
satu
padafrekuensi yang tinggi (> 80 %).
0t o o .L 50.00 45.00 10.00 35.00 30.00 25.0 0 20.00 15.00 10.00 s.00 0.00 M8M Tsrgdr MTM 15.53 tEo UJW|C lldpah MBM
Kon 52'@-2 Klon 52-CC-,4 loon Po.l3025 O Uninukl@l Awal
E Uninrkl*t AkHr
Gambarl' Reratapersentasemikrosporauninukleatawal,uninukleatakhirdantepungsari
menurutklondan morfologi malai belum muncul (MBM) dan
malai relah muncul (MTM),-serta letak pada ujung, tengah dan pangkal malai.
mikrospop berada pada pada tahap berinti
satu
apabila
malai
belum
memuncul_kanbunganya. Akan tetapi bagi malai yang telah nrernunculkan bunganya, persentase nrikro_
spora yang berada pada tahap berinti satu
menunjukkan penurunan
proporsi
ataujumlahnya mengikuri letak pacla malai yakni semakin mengarah ke ujung malai, semakin
besar penu-runannya. Artinya, perkembangan
Di
pihak lain, malai yang telah me_munculkan bunganya menunjukkan bahwa
persentase mikrospora berinti lebih dari
satu
atau
mikrosporapada tahap tepung
sarimenjadi semakin meningkat seiring dengan peningkatan
ukuran
pemunculan- bunga.Dilihat dari letak mikrospora
di
dalam*uLi,
baik
pada bagian ujung dan bagian tengahmaupun
pada
pangkal
malai,
seluruh86
mikrospora meningkat ke perkembangan lebih lanjut (tepung sari) dimulai dari bagian ujung malai. Hasil uji+ perbedaan antara malai yang belum dengan malai yang telah memunculkan
bunganya menunjukkan bahwa rerata
per-sentase
mikrospora uninukleat
awal, uninukleat akhir, dan tepung sari pada keduabentuk
morfologi
malai
menunjukkanperbedaan yang signi-fikan (Tabel 4).
Tabel 4. Perbedaan rerata persentase tiga tahap
perkembangan mikrospora antara
malai yang
belurr
dan yang teiahmemunculkan bunganya
Morfologi
Uninukleat
Uninuklcatterdiri dari cabang utama (rachis) dan
anak-anak
cabang(rachilla) yang
merupakantempat tcrbentuknya susunan cabang pertama,
kedua dan ketiga. Pada bagian bawah atau pangkal
malai
berukuran besar kemudian ukurannya semakin kecil sampai pada ujungmalai yang hanya
terdiri dari
satu cabang.Cabang utama
(main
axes)atau
susunanpertama (first order) terbentuk secara lateral
anak cabang (lateral axes) atau susunan kedua
(second order) yang lebih panjang, dan dari anak cabang-anak cabang
ini
terbentuk anak-anak cabang atau susunan ketiga (third order).Bulir (spikelets) yang terdiri dari bulir bagian
pangkal (sessile.florers) dan bulir di atas bulir pangkal Qtedicellate
/lorets)
terbentuk padaanak cabang lateral dan anak-anak cabang. Hasil pengamatan dalam penelitian ini
menunjr.rkkan bahwa kernatangan tepung sari
terjadi mulai
dari
ujung malai
utama danujung cabang malai pada cabang bagian ujung malai utama hingga pada cabang malai paling
bawah
dari
malai
utama.Hal
ini
karena persentase terbanyak mikrospora yang sudahmemasuki tahap perkembangan lebih dari satu inti atau tepung sari adalah pada sessile florets
dan ujung malai. Pada pengamatan
ini.
tidak ditemukan mikrosporayang
masih berada pada tahapdiad
dan
tetrad
sebagaimanadilaporkan
oleh
Raghavan(l9Sg)
padatanaman padi, Summers
e/ al.
(1992) padatanaman tomat, da Silva-Lauxen et al. (2003) pada tanaman kedelai, dan lndrianto
et
al. (2004) pada tanaman cabai besar. Ketiadaan mikrosporatahap
diad
dan tetrad
padapenelitian
ini
disebabkanoleh
umur malaiyang
mulai
memasuki tahap perkembanganIebih lanjut yang
ditandai
oleh
semakinpanjangnya ukuran
malai dan
munculnyabunga di atas gulungan daun bendera sehingga
semakin banyak
mikrosporayang
telalr memasuki perkembangan lanjut.Berdasarkan ukuran panjang antera
pada
tanamanpadi
(Oryza sativa
L.),Raghavan (1988) melaporkan bahwa semakin
panjang
ukuran antera,
semakin
tinggipersentase mikrospora yang berada pada tahap
perkembagan
lebih lanjut.
Sementara itu,semakin
lanjut
pertumbuhanmalai
akanMalai
awal
akhirTepung sari Belum membuka Telah menrbuka 93,22 a 47,79 b 5,84 p 23.30 q 0,94 x 28,75 y
Keterangan
:
Nilai-nilai yang diikuti huruf yangberbeda pada kolom yang sama
berarti berbeda signifikarr menurut
u-ii T-Student
Demikian pula, perbedaan antar klon terhadap rerata persentase tiga tahap
perkem-bangan mikrospora melalui Lr.ii -f-Fishcr (LSD,
Leust
Square
DifJbrencc)
menunjukkan adanya perbedaan yang signifikandi
antara ketiga klon yang diamati (Tabel 5).Tabel 5. Perbedaan rerata persentase tiga tahap
perkembangan ntikrospora antara
tiga klon tebu.
Klon
Uninukleat
Uninukleat TepurrgAwal
akhir
saris20c2
82,90 a 14,05 k 3,05 o52OC4
72,25a
18,45k
9,30 p_loJ!9{__wsa t3,t7k
37,08qKeterangan
:
Nilai-nilai yang diikuti huruf yangberbeda pada kolom yang sama
berarti berbeda signifikan menurut
uji T-Student
Menurut Moore ( 1987) bunga
tana-man tebu berbentuk malai Qtanicle) mernbuka
setelah muncu dari bungkusan daun benclera,
senrakin panjang
pula
ukuran
altteranya sehingga pada ukuran bulir sepanjang 7.000-9.142
pm,
antera akan berukuran 1.125-1.750 pm, dan umumnya, mikrospora telah
berada pada tahap uninukleat dan sebagian
telah memasuki tahap binukleat.
Meskipun beragam ukuran panjang kuncup pada
tiga
kultivar
tanaman toltlat, masing-masing: A.Craig, L-680A, dan Licato, ukuran panjang antera ketiganya relatif samayakni
2,8-3,5 rnm,
semuakultivar
hanyamengandung
mikrospora
berinti
satu(Summers et
al.,
1992). Demikian pula pada tiga kultivar kedelai berturut,turut '. Decada,IAS5, dan RSZ dilaporkan bahwa bunga yang
berukuran 1,5-2,9
mm
akan
mengandung mikospora mulai pada tahap sebelum diad hingga tetrad, sedangkan bunga yang telahberukuran 3,0*33,5
mm
akan mengandungmikrospora pada tahap uninukleat awal hingga uninukleat
akhir (da
Silva-Lauxener
al.,2003).
Pada tanarnan cabai besar, Indrianto et
al.
(2004) menguraikan bahwa mikrospora pada tahap tetrad hanya dijumpai pada kr-rncupbunga yang berukuran hingga 0,3-{,5 cm dan
anteranya berwarna mulai
dari hijau
rnuda,sebagian
hijau dan
sebagianungu,
akanmengan-dung rnikrospora uninukleat. Setelah bungan berukuran 0,6
-
0,8 cm dan anteranyaberwarna
ungu
muda, bunga hanya akanmengandung mikrospora binukleat. Dengan
demikian, tahap perkembangan mikrospora di
dalam antera sangat ditentukan oleh ukuran
malai, bunga dan antera, sehingga penggunaan
ciri
morfologimalai atau
ukuran kuncup merupakan cara yang tepat dalarn menentukantahap perkembangan mikrospora yang sesuai
bagi pelaksa-naan pemuliaan haploid melalui kultur mikrospora secara in vitro.
KESIMPULAN DAN SARAN Berdasarkan pengamatan
dua
ciri rnorfblogi pertumbuhan malai terhadap jumlah mikospora pada tahap satuinti
lebih dari 50Yo, dapat disimpulkan sebagai berikut: (a) pada
malai bunga yang masih terbungkus di dalam
81
kelopak daun bendera terdapat mikrospora umlrmnya pada tahap uninukleat;
(b)
rnalai yang telah memunculkan bunganyadi
atas gulungandaun
bendera cenderung akanmengandung mikrospora yang sebagian besar pada tahap binukleat atau tepung sari, dan; (c) senrakin ke ujung letak bulir atau antera pada
malai atau cabang malai, jumlah mikrospora yang berada pada tahap uninukleat semakin berkurang.
lsolasi mikrospora paling tepat di-lakukan pada saat bunga belum muncul di atas
gulungan kelopak
daun
bendera karenapersentase mikrospora berinti satu mencapai
maksi-malnya
ketika
bungamasih
beradadalam gulungan lielopak daun bendera.
DAFTAR PUSTAKA
Bhojwani, S.S. and S.P. Bhatnagar, 1999, .'The
embriology
of
angiosperms", pp. 30g-321. 4'h revised and enlarged' edition. Vikas Publishing House pVT. LTD.Blakeslee, A., J. I3elling, M.E. Farnham and A.D.
Berger, 1922.
*A
haploid mutant in thejinrson weed Dqlura
slramoniunt,'.Science. SS: 646-647.
Chen, 2.H., C. Qian, M. Qin, C. Wang, C. Suo, F.
Chen and Z. Dheng, 1979, ,,'fhe inducrion
of
pollen plantsof
sugarcane". .,lnau.Rep. Inst. Genet. Acad..tir., pp.9l_93. Cistue, L., M.P. Valles, B.Echavarri, M. Sans, and
A.M.
Castiilo, 20Q4. ..production ofbarley doubled haploids by anther and
microspore culture", pp.l-17. Dalam :
Mujib, A., M.J. Cho, S. predieri, and S.
Barnerjee (eds.). ln Vitro Application in
Crop Improvement. Science publishers
lnc., Enfield, USA.
Cordewener, J.H.G., J.B.lU. Custers, H.J.M. Dons
and
M.M.
Van
Lookeren Campagne,1996, "Molecular and biochemical events
during
the
inductionof
microsporeembryo-genesis", pp.
lll-124.
Dalatn ..Jain, S.M.,
S.K.
Sopory,and
R.E.Veilleux (Eds.).
In
Vitro
HaploidProduction
in
Higher plants.Vol.
l, Fundamental Aspcctsand
Methods.88
Kluwer
Acadenric Publishers. TheNetherlands.
da Silva-Lauxen, M., E. Kaltchuk-Santos, C.y. Hu,
S.M. Callegari-Jacques and M.H. Boda-nese-Zanettini,
2003.
..Association between floral bud size and developnrental stage in soybean microspores" . BrassilianArchives
of
Biolog_v&
Technology,46()z 8-14.
Fitch, M.M. and P.H. Moore, 1983, .,Haploid production
from
antherculture
ofSaccharum
spontaneumL".
Z.Pflanzenphysiol, 109: 197 -206.
lndrianto,
A., 2003,
"Cyological
andultrastructural features
of
initiation ofwheat
microspore
embryogenesis,'.Biologi,3(2): 65-79.
lndrianto,
A.,
E.
Semiarti dan Surifah, 2004."Produksi galur murni melalui induksi
embrio-genik mikrospora cabai merah dengan stres". Zuriat, l 5(2): 1 33- 1 39.
Kasha, K.J., E. Sirnion, R. Oro, e.A. yao,'l'.C. llu and A.R. Carlson,200 l,..An improved
ir
yilro techniquefor
isolated nricrospore culture of barley". Eupltytica, I2Al. 379-3 85.Kush, G.S. and S.S. Virmani. t996, ..Haploids in
plant breeding", pp.12-17. Dalam
:
Jain, S.M., S.K. Sopory, and R.E. Villeux (eds.)In
Vitro Haploid.Productionin
Higher Plants. Vcll.l. Fundamental Aspects andMethods. Kluwer Academic publishers,
Dord-recht, Boston,
London.
. .Li,
H.
and P. Devaux, 2003, ..High frequencyregeneration
of
barley doubled haploid plants from isolated microspore culture,'.Plant Science, l64z 379-386.
Liu, W., M.Y. Zheng, E.A. Polle, and C.F. Konzak,
2002, "Highly efficient doubled-haploid
production
in
wheat (Triticun aesliwrntL.)
via
induced
m icrosporeembryogenesis". Crop Science, 42:
6t6-692.
Lu,
W. L.
and C.S.Kuo,
t984. ..Cltologicalobservation
of
microsporogenesis andpollen developntent
in
wheatin
vivo',.Acta Botunico Sinica, 26-. 26-33.
Maheswari, S.C., A. Rashid and A.K. Tyagi, 1983.
"Anther pollen culture for production of
haploids
and
their
utility".
IAPTC Newsletter, 4l:2-7.Moore, P.H. 1987. "Anatomy and morphology",
pp.85-142. Dalam
:
Heins, D.J. (ed,). Sugar-caneImprovement
ThroughBreeding. Developments jn Crop Science I
l.
Elsevier, Amsterdam,New
york,Oxford, Tokyo.
Moore, P.H., C. Nagai and M.M. Fitch, 19g9,
"Production and evaluation of sugarcane
haploids". Proc.
Int.
Soc. Sugar Cane Technology, 20: 599-607 .Palmer, C.E. and W.A. Keller, lgg7, ..pollen
embryos",
pp.
392-422.Dalam
:Shrivanna, K.R. and V.K. Sawney (eds.).
Pollen Biotechnology for Crop production
and Improvement. Cambridge University
Press. U.K.
SAS Institute Inc., 1989-1996, ..SAS/STAT User's
Cuide Release 6.12". SAS lnstitutc. lnc.. Cary, N.C.
Steel, R.C.D. and J.H. Torrie, 1981, ..principles and procedures
of
statistics". 2nd ed.McOraw-Hill Editions, Singapore etc. 633pp.
Suaib, S. Madsen, A. Olesen dan S.B. Andersen, 1997, "Seleksi tanaman rumput makanan ternak tahunan Ryegrass (Lolium perenne
L.)
yang tanggap terhadap perlakuan prakultur antera". Zuriat, 8(2): 90-93.Suaib,2000, "Determinasi enam kultivar lokal padi
ladang asal Kendari yang mengandung
tepung
sari
berinti satu (uninucleate) untuk pemuliaan in vitro". Zuriat, l0(2):2l-27.
Summers, W,L., J. Jaramillo and T. Bailey, 1992.
"Microspore developmental stage and anther length influence in the induction of
tdmato anther callus,'. Hortscience, 27(7): 838-E40.
Zhang,
2., Y.
Lu,X.
Liu
and J. Feng, 2005, "Nuclear and cell migration during pollendeve-lopment in rice (Oryza sativa L.),,.
Sex Plont Reproduction,