Peningkatan Pertumbuhan dan Kualitas Buah Cabai Merah
Besar dengan Hormon Tumbuh Asal Bakteri Endofit
(Increasing the Growth and Quality of Red Chili with Growth Hormone from
Endophytic Bacteria)
Rafika Yuniawati1*, Siti Fatimah2, Reni Indrayanti2, Ifa Manzila1, Tri P. Priyatno1, dan Dwi N. Susilowati1 1Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Jl. Tentara Pelajar 3A, Bogor 16111 Indonesia
Telp. (0251) 8622833; Faks. (0251) 8622833; *E-mail: [email protected]; [email protected]
2Program Studi Biologi, FMIPA, Universitas Negeri Jakarta, Gd. Hasjim Asj’arie, Jl. Rawamangun Muka Raya, Jakarta Timur 13220 Indonesia
Diajukan: 16 April 2019; Direvisi: 6 September 2019; Diterima: 6 September 2019
ABSTRACT
Red chili is a very important horticultural commodity in Indonesia having low productivity and quality. Cultivation method needs to be improved including the use of exogenous growth hormones. The purpose of this study was to determine (1) the optimum concentration of IAA and GA growing hormones from isolate B6.2 in stimulating plant growth and improving the quality of large red chili fruit; (2) molecular identity of the B6.2 bacterial isolate. The growth hormone content of B6.2 isolates using HPLC obtained 0.49 ppm IAA and 64.53 ppm GA. The growth hormone potential test on the growth and quality of chili was carried out with a concentration of 1, 3, 5 ml/l, while water and synthetic hormones was used as negative and positive control, respectively. The experimental design used was a Complete Random Design with the foliar spray application to the plant canopy three times during the growth period. The results showed the best concentration in increasing plant height, fruit weight, shooth wet, and dry weight compared to controls at the age of 76 days after planting (dap) was a concentration of 5 ml/l, with the values of 71.7±0.9 cm, 94.7±0.3 g, 11.5±0.43 g, and 1.4±0.09 g, respectively. The molecular identification showed that B6.2 isolate was classified as Bacillus vallismortis with 100% homology. The growth hormone from isolate B6.2 has the potential to increase growth and production of red chili plants.
Keywords: Bacillus vallismortis, Capsicum annuum, bioactive compounds.
ABSTRAK
Cabai merah merupakan komoditas pertanian yang sangat penting di Indonesia yang mengalami penurunan produktivitas dan kualitas. Perbaikan teknik budi daya sangat diperlukan di antaranya dengan mengaplikasikan hormon tumbuh eksogen. Tujuan penelitian ini ialah (1) mengetahui konsentrasi optimum hormon tumbuh IAA dan GA asal bakteri endofit isolat B6.2 dalam memacu pertumbuhan tanaman dan meningkatkan kualitas buah cabai merah besar; (2) menentukan identitas spesies bakteri isolat B6.2 secara molekuler. Kandungan hormon tumbuh asal isolat B6.2 diperoleh sebesar 0,49 ppm IAA dan 64,53 ppm GA (1:131) dengan menggunakan HPLC. Uji potensi hormon tumbuh terhadap pertumbuhan dan kualitas buah cabai dilakukan dengan konsentrasi 1, 3, 5 ml/l, dengan menggunakan air dan hormon sintetik hormonik berturut-turut sebagai kontrol negatif dan positif. Percobaan ini menggunakan Rancangan Acak Lengkap dengan aplikasi penyemprotan ke tajuk tanaman sebanyak tiga kali selama periode pertumbuhan. Hasil penelitian menunjukkan konsentrasi terbaik dalam meningkatkan tinggi tanaman, bobot buah, dan bobot basah dan kering tajuk dibanding dengan kontrol pada umur 76 hari setelah tanam (hst) yaitu 5 ml/l, dengan nilai berturut-turut 71,7±0,9 cm, 94,7±0,3 g, 11,5±0,43 g, dan 1,4±0,09 g. Identifikasi secara molekuler menunjukkan isolat B6.2 tergolong Bacillus vallismortis dengan tingkat homologi 100%. Hormon tumbuh asal isolat B6.2 berpotensi meningkatkan pertumbuhan dan produksi tanaman cabai merah besar.
Kata kunci:Bacillus vallismortis, Capsicum annum, senyawa bioaktif.
PENDAHULUAN
Cabai merah besar (Capsicum annuum L.) me-rupakan komoditas hortikultura penting di Indonesia. Kebutuhan buah cabai nasional terus meningkat se-iring dengan semakin bervariasinya jenis menu ma-kanan dan produk-produk industri dan pengobatan yang memanfaatkan cabai (Syukur 2007).
Berdasarkan data Badan Pusat Statistik, pro-duksi cabai merah nasional tiap tahunnya meningkat dalam tiga tahun terakhir sebesar 1.045.200 ton pada tahun 2015 menjadi 1.206.750 ton pada tahun 2018 (Badan Pusat Statistik 2019). Produksi cabai merah besar nasional sebetulnya telah mencukupi kebutuh-an nasional tahunkebutuh-an, namun fluktuasi produksi terjadi sepanjang tahun menyebabkan terjadi lonjakan har-ga yang berimbas pada inflasi. Potensi produktivitas cabai merah besar nasional terus meningkat, namun areal panennya semakin menurun selama tiga tahun terakhir, dari 128.734 hektar pada tahun 2014 menjadi 123.404 hektar pada tahun 2016 (Badan Pusat Statistik 2016). Dengan demikian, teknik budi daya yang benar dan menguntungkan perlu diperhatikan sehingga da-pat meningkatkan pertumbuhan dan kualitas tanam-an cabai merah besar, di tanam-antartanam-anya dengtanam-an memtanam-an- meman-faatkan zat pemacu tumbuh (ZPT) eksogen.
Penggunaan pemacu tumbuh tanaman pada konsentrasi yang optimal mampu mengatur pertum-buhan, parameter panen, dan kualitas buah. Tanam-an mampu memproduksi pemacu tumbuh secara alami, namun penambahan ZPT eksogen dapat membantu mempercepat pertumbuhan dan perkem-bangan tanaman (Sukmadi 2012). ZPT eksogen dapat diperoleh dari air kelapa muda (Ernawati et al. 2017), ekstrak bawang merah (Darojat 2014), dan mikroor-ganisme dari kelompok bakteri endofit (Agustian et al. 2010).
Bakteri endofit merupakan bakteri yang hidup dan berasosiasi dengan jaringan tanaman tanpa memberikan kerugian pada tanaman tersebut (Bacon dan White 2000). Beberapa kelebihan bakteri endofit antara lain mikroorganisme ini banyak terdapat di dalam tanah atau jaringan tanaman sehat, produksi massal lebih mudah dan lebih cepat daripada mikro-organisme lain seperti kapang. Bakteri endofit dapat menguntungkan tanaman inangnya, di antaranya menstimulasi pertumbuhan tanaman, memfiksasi nitrogen, dan meningkatkan sistem pertahanan ta-naman terhadap gangguan penyakit. Selain itu, bakteri endofit mampu memproduksi senyawa antibakteri, enzim, asam salisilat, etilena, dan senya-wa metabolit sekunder yang berperan dalam meng-induksi ketahanan tanaman (Taghavi et al. 2005).
Be-berapa strain bakteri endofit yang mampu menghasil-kan pemacu tumbuh tanaman berupa indole-3-acetic acid (IAA), di antaranya Azotobacter chroococum, A. vinelandii, Pseudomonas putida, P. aeruginosa, dan Klebsiella pneumoniae (Rubio et al. 2000).
Gen 16S ribosomal RNA (16S rRNA) merupakan marka molekuler yang banyak digunakan untuk identifikasi dan analisis keragaman genetik mikroba (Srinivasan et al. 2015) karena memiliki daerah yang conserved dan hypervariable. Daerah yang conserved mencerminkan hubungan filogenetik antarspesies dan daerah yang bervariasi mencerminkan perbeda-an perbeda-antarspesies (Mitreva 2017). Penelitiperbeda-an keragamperbeda-an genetik berdasarkan gen 16S rRNA berhasil meng-identifikasi spesies rizobakteri penghasil IAA, seperti Bacillus subtilis, B. amyloliquefaciens, B. velezensis, dan Brevundimonas olei (Lestari et al. 2017), Azospirillum brasilense dan A. zeae (Ayyaz et al. 2016).
Beberapa hasil penelitian menunjukkan bahwa penggunaan bakteri endofit secara in vitro dapat me-ningkatkan pertumbuhan beberapa jenis tanaman, seperti jagung (Istiqomah dan Trijoko 2014), padi (Munif et al. 2012), dan lada (Kartikawati dan Gusnaini 2018). Hasil penelitian Munif et al. (2015) menunjukkan bahwa penggunaan bakteri endofit yang berasal dari tanaman perkebunan mampu me-ningkatkan pertumbuhan tanaman tomat hingga 60%. Beberapa strain Bacillus sp. juga dapat membantu meningkatkan pertumbuhan tanaman (Wahyudi et al. 2011). Dari hasil penelitian inokulasi bakteri endofit, P. diminuta L. dan B. substilis (C) mampu meningkat-kan pertumbuhan bibit kopi arabika (Asyiah et al. 2015), bibit kakao (Puspita et al. 2018), bibit kelapa sawit (Puspita et al. 2013), dan jagung (Saylendra dan Firnia 2013). Penggunaan Bacillus sp. dengan limbah sawit mampu memacu pertumbuhan panjang malai, meningkatkan berat 100 butir gabah, persentase gabah bernas, berat gabah kering per tanaman, dan bobot kering pada tanaman padi sawah (Tinendung et al. 2014).
Penelitian yang telah dilakukan oleh Susilowati et al. (2017) melaporkan mikroba endofit menghasil-kan hormon bagi tanaman, terutama mikroba yang menunjukkan pengaruh merangsang pertumbuhan (growth promoting microorganism). Pada koleksi kultur mikroba BB Biogen terdapat koleksi bakteri endofit B6.2 dari hasil isolasi tanaman cabai merah besar varietas Landung. Isolat ini tidak patogenik terhadap manusia dan dapat menghambat Colletotrichum capsici dengan persentase pengham-batan sebesar 60%. Isolat bakteri endofit B6.2 juga mampu menghasilkan ZPT berupa IAA dengan
konsentrasi sebesar 41,22 ppm (Fitriyah 2015). Tujuan penelitian ini ialah (1) mengetahui konsentrasi optimum hormon tumbuh IAA dan GA asal bakteri endofit isolat B6.2 dalam memacu pertumbuhan tanaman dan meningkatkan kualitas buah cabai merah besar; (2) menentukan identitas spesies bakteri isolat B6.2 secara molekuler.
BAHAN DAN METODE Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Februari– Juli 2018 di Laboratorium Mikrobiologi dan Rumah Kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB Biogen).
Penyiapan Tanaman Uji
Benih cabai merah varietas Tanjung II yang di-peroleh dari UPBS Balai Penelitian Tanaman Sayuran (Balitsa), Lembang disemai pada polytray 105 lubang (20 cm 10 cm 5 cm). Bibit dipindah tanam setelah berumur 30 hari ke polybag (30 cm 45 cm) berisi 4 kg media tanam yang berupa campuran tanah dan pupuk kandang kotoran sapi (4:1). Polybag kemudian diletakkan secara teratur berjarak 75 cm (antarbaris tanaman) 50 cm (dalam baris tanaman). Pemupuk-an dilakukPemupuk-an dengPemupuk-an pupuk urea, TSP, dPemupuk-an KCl masing-masing sebanyak 7 g untuk setiap jenis pupuk per polybag yang diberikan sebelum tanam.
Perbanyakan Kultur Bakteri Endofit Isolat B6.2 Perbanyakan kultur cair bakteri endofit isolat B6.2 mengikuti metode Tien et al. (1979). Sebanyak 1 ose isolat murni bakteri dari media agar miring NA di-inokulasikan ke dalam tabung ulir berisi 10 ml media NB, kemudian diinkubasi di dalam incubator shaker dengan kecepatan 120 rpm pada suhu ruang selama 12 jam. Selanjutnya, sebanyak 10 ml kultur starter diinokulasikan ke dalam labu Erlenmeyer 1.000 ml yang berisi 500 ml media NB yang telah ditambahkan triptofan sebagai prekursor pembentukan senyawa bioaktif IAA. Kultur 500 ml tersebut selanjutnya diinku-basi dalam incubator shaker dengan kecepatan 120 rpm pada suhu kamar selama 7 hari.
Ekstraksi dan Kuantifikasi Hormon IAA dan GA Asal Bakteri Endofit Isolat B6.2
Menggunakan HPLC
Pembuatan ekstrak senyawa bioaktif IAA meng-ikuti metode Tien et al. (1979). Kultur cair bakteri di-sentrifugasi dengan kecepatan 6.000 rpm pada 4ºC selama 10 menit. Supernatan yang diperoleh diatur
pH-nya menjadi 2,8 dengan penambahan HCl 1 N, lalu ditambahkan dietil eter dengan perbandingan 1:1 dan diinkubasi di ruang gelap selama 4 jam dilanjut-kan dengan inkubasi di dalam refrigerator 4ºC selama 12 jam.
Filtrasi dilakukan dengan menggunakan labu pe-misah untuk memperoleh fase dietil eter. Fase dietil eter kemudian dievaporasi hingga kering dengan rotary evaporator hingga tersisa ekstrak ±1 ml. Ekstrak yang diperoleh dilarutkan dalam 10 ml metanol.
Pembuatan ekstrak senyawa bioaktif GA meng-ikuti metode yang telah dilakukan oleh Tien et al. (1979). Kultur cair bakteri disentrifugasi dengan ke-cepatan 10.000 rpm pada 4ºC selama 10 menit. Supernatan yang diperoleh ditetapkan pH-nya men-jadi 2,0 dengan menambahkan HCl 5 N, selanjutnya ditambahkan etil asetat dengan perbandingan 1:1.
Filtrasi dilakukan dengan menggunakan labu pemisah untuk memperoleh fase etil asetat. Fase etil asetat kemudian dievaporasi hingga kering dengan rotary evaporator hingga tersisa ekstrak sebanyak ±1 ml. Ekstrak dilarutkan dengan menambahkan 10 ml air steril yang mengandung Tween® 80 (0,05 %).
Pengukuran konsentrasi senyawa bioaktif asal bakteri dilakukan dengan HPLC mengikuti metode Mehnaz dan Lazarovits (2007). Sampel hasil ekstraksi disaring dengan kertas saring Millipore® (Merck, Jerman), kemudian dianalisis dengan HPLC Agilent 1260 Infinity (Thermo Scientific, AS) dengan kolom Poroshell 120 SB-C18 (Agilent Technologies, AS). Sampel yang diinjeksikan sebanyak 10 µl dengan fase gerak metanol:akuades dengan asam asetat 1% (30:70:1 [v/v]), flow rate 1,5 ml/menit, dan panjang gelombang 254 nm. Hasil pengukuran kuantitatif senyawa bioaktif (IAA dan GA) dihitung berdasarkan perbandingan luas area grafik senyawa bioaktif dengan IAA dan GA standar (Sigma-Aldrich, Jerman). Pengukuran konsentrasi IAA dan GA ditunjukkan dengan waktu retensi kromatogram sampel relatif sama dengan retensi kromatogram IAA dan GA standar. Konsentrasi senyawa bioaktif asal bakteri di-hitung dengan menggunakan variabel luas area IAA dan GA standar dan luas area sampel.
Uji Efikasi Hormon IAA dan GA
Perlakuan yang diberikan berupa penyemprotan 5 ml/l, 3 ml/l, dan 1 ml/l campuran senyawa bioaktif GA dan IAA dengan perbandingan 1:2 (Susilowati et al. 2017) pada tanaman sebanyak tiga kali, yaitu pada umur 45, 60, 75 hst, kontrol negatif berupa air dan kontrol positif berupa ZPT sintetik (Hormonik®)
dengan konsentrasi yang direkomendasikan (2 ml/l). Penelitian ini disusun dengan menggunakan Ran-cangan Acak Lengkap (RAL) dengan sembilan ulang-an. Parameter pengamatan berdasarkan Prajnanta (2001), yaitu (1) pengukuran tinggi tanaman dilaku-kan pada umur 30, 46, 61, 76 hst diukur dari pangkal batang sampai ke ujung titik tumbuh tertinggi; tinggi tanaman kumulatif dari waktu ke waktu selama pe-nelitian diwakili oleh the area under high progress curve (AUHPC) (Campbell dan Madden 1990):
AUHPC =
dengan n = jumlah pengamatan, Y = tinggi tanaman (cm), t = waktu pengamatan (hari); (2) jumlah buah panen per tanaman dengan kriteria buah berwarna merah; (3) bobot buah per tanaman; (4) panjang buah dalam cm diukur dari bagian pangkal hingga ujung buah; (5) bobot basah akar dan tajuk per tanaman; (6) bobot kering akar dan tajuk setelah dimasukkan ke dalam oven 105ºC selama 2 hari. Parameter kualitas tanaman cabai menurut Badan Standardisasi Nasional dalam SNI No. 01-4480:1998 meliputi keseragaman ukuran (panjang buah dan garis tengah pangkal). Kualitas buah grade I, II, dan III mempunyai panjang buah dan garis tengah pangkal buah masing-masing sebesar 12,0–14,0 cm dan 1,5– 1,7 cm, 9,0–11,0 cm dan 1,3–1,5 cm, <9,0 cm dan <1,3 cm (Badan Standardisasi Nasional 1998).
Data-data hasil pengamatan selanjutnya dianali-sis dengan one way ANOVA pada derajat kepercaya-an 95% dkepercaya-an dilkepercaya-anjutkkepercaya-an dengkepercaya-an uji Dunckepercaya-an pada α = 0,05 dengan program SPSS 16.0.
Identifikasi Isolat Bakteri Endofit B6.2 secara Molekuler
Isolat bakteri endofit B6.2 diidentifikasi secara molekuler dengan primer 16S rRNA mengikuti pro-sedur Lane (1991). Produk PCR gen 16S rRNA isolat bakteri endofit tersebut selanjutnya disekuensing di Macrogen Korea, Korea Selatan. Hasil sekuen dianali-sis dengan program BioEdit dan sebelumnya dilaku-kan pengeditan basa-basa nukleotida dengan prog-ram ChromasPro. Hasil sekuensing kemudian di-analisis homologinya dengan BLASTn berdasarkan basis data 16S rRNA pada situs National Center for Biotechnology Information (NCBI 2019). Konstruksi filogeni antara isolat dan sekuen strain Bacillus lain-nya menggunakan metode Maximum Likelihood dengan bootstrap 1.000 pada program MEGA6 (Tamura et al. 2012).
HASIL DAN PEMBAHASAN Konsentrasi Hormon Tumbuh
Rasio senyawa bioaktif IAA:GA asal isolat B6.2 dan hormon sintetik yang terukur dengan meng-gunakan HPLC berturut-turut sebesar 1:131 dan 1:2. (Tabel 1). Perbedaan rasio senyawa bioaktif ini kemungkinan terjadi karena kondisi kultur dan media yang digunakan untuk isolat B6.2 belum optimal se-hingga diperlukan penelitian lanjutan untuk menemu-kan kondisi kultur dan media yang cocok pada isolat bakteri genus Bacillus untuk menghasilkan senyawa bioaktif yang mampu meningkatkan pertumbuhan dan produksi buah cabai. Hasil penelitian Tien et al. (1979) menunjukkan bahwa kondisi kultur dan media berpengaruh terhadap produksi IAA yang dihasilkan oleh A. brasilense. Selain itu, diperlukan penelitian tentang penambahan bahan suspensi pada senyawa bioaktif hormon tumbuh yang dihasilkan oleh isolat B6.2 sehingga stabilitas hormon terjaga.
Pertumbuhan dan Daya Hasil Tanaman Potensi bakteri endofit untuk meningkatkan per-tumbuhan berbagai jenis tanaman, baik secara in vitro maupun di rumah kaca, telah banyak dilakukan, seperti penelitian bakteri endofit yang diisolasi dari tanaman jahe merah terhadap pertumbuhan benih lada (Kartikawati dan Gusnaini 2018), pertumbuhan tanaman jagung (Istiqomah dan Trijoko 2014), dan pertumbuhan tanaman sambiloto (Gusmaini et al. 2013). Namun, penggunaan ekstrak fitohormon asal bakteri endofit yang diaplikasikan secara langsung pada tanaman di lapangan belum pernah dilaporkan. Hasil penelitian ini merupakan laporan pertama kali penggunaan ekstrak fitohormon asal isolat bakteri endofit B6.2 yang diaplikasikaan secara langsung pada tanaman cabai di lapangan.
Hasil pengamatan menunjukkan bahwa tinggi tanaman cenderung meningkat pada semua umur pengamatan untuk kedua jenis hormon dibanding dengan kontrol negatif (Tabel 2). Perlakuan terbaik ditunjukkan oleh konsentrasi penyemprotan hormon sebesar 5 ml/l. Hal ini juga terlihat dari hasil per-hitungan AUHPC, yaitu konsentrasi 5 ml/l berpotensi memberikan pengaruh terhadap tinggi tanaman cabai (Tabel 2).
Tabel 1. Hasil pengukuran konsentrasi ekstrak IAA dan GA asal isolat B6.2 menggunakan HPLC.
Sampel Senyawa bioaktif (ppm)
IAA GA
Ekstrak isolat B6.2 0,49±0,14 64,53±0,63 Hormon sintetik 1,26±0,12 2,60±0,19
Penyemprotan senyawa bioaktif asal isolat B6.2 dan ZPT sintetik tidak memberikan pengaruh nyata terhadap jumlah buah total, kualitas buah I, dan kualitas buah III, namun berpengaruh nyata terhadap bobot buah cabai. Pemberian hormon tumbuh dengan konsentrasi 5 ml/l memberikan pengaruh nyata terhadap jumlah buah kualitas II dan bobot buah dibanding dengan perlakuan lainnya (Tabel 3).
Semakin banyak jumlah buah, semakin me-ningkat bobot buah (Tabel 3). Penelitian Salli et al. (2016) menunjukkan bahwa jumlah buah tomat yang dihasilkan memengaruhi berat buah per petak. Hal ini berkaitan dengan fotosintat yang ditranslokasikan ke bagian generatif berupa buah. Apabila fotosintat yang ditranslokasikan banyak, jumlah buah semakin banyak dan berat buah per petak pun meningkat.
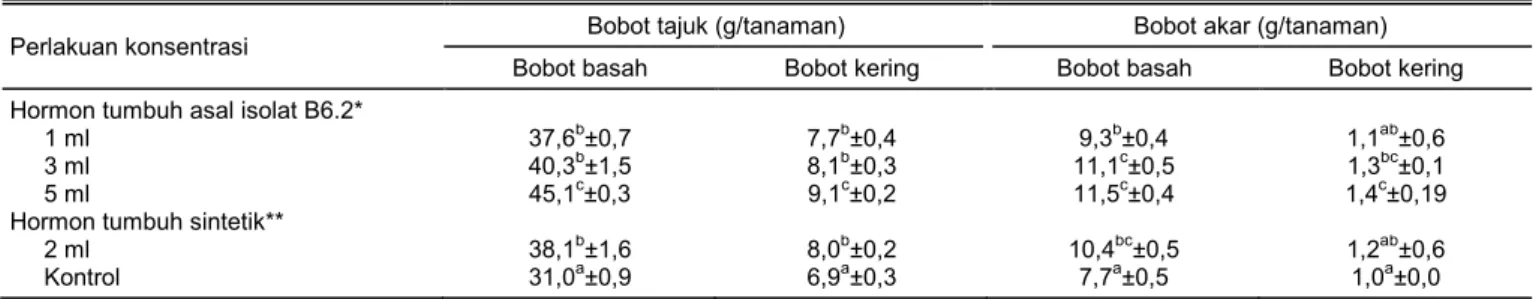
Hasil pengamatan bobot tajuk dan bobot akar menunjukkan bahwa penyemprotan senyawa bio-aktif pada konsentrasi 5 ml/l memberikan pengaruh nyata terhadap bobot basah dan bobot kering dibanding dengan perlakuan hormon tumbuh sintetik (Tabel 4). Adanya peningkatan bobot basah dan bobot kering tajuk menunjukkan bahwa kandungan bahan organik banyak terserap oleh tanaman. Hal ini menunjukkan bahwa proses fotosintesis tanaman tersebut berjalan dengan baik sehingga memengaruhi proses pertumbuhan, baik vegetatif maupun generatif tanaman. Bobot basah tajuk menunjukkan aktivitas metabolisme selama masa pertumbuhan tanaman.
Pertumbuhan tanaman dapat berjalan dengan baik jika faktor genetik dan lingkungan tanaman berada pada kondisi optimal. Faktor lingkungan yang dimak-sud ialah tanah dan iklim (Hayati 2010). Peningkatan bobot basah tajuk merupakan dampak dari proses metabolisme yang normal dengan ketersediaaan air yang cukup. Hasil dari proses tersebut dibutuhkan untuk pertumbuhan dan perkembangan tanaman (Kuswandi dan Sugiyarto 2015). Bobot kering tajuk menunjukkan pola tanaman menyimpan hasil foto-sintesis (Ariyanti et al. 2017). Bobot kering tajuk me-nunjukkan penumpukan senyawa organik yang telah disintesis tanaman dari senyawa anorganik (Kastono et al. 2005). Kemampuan laju sel-sel untuk mem-belah, membesar, dan memanjang akan memenga-ruhi berat kering tanaman (Fathonah dan Sugiyarto 2009).
Hasil penelitian ini menunjukkan bahwa penyemprotan senyawa bioaktif asal isolat B6.2 pada konsentrasi 5 ml/l memberikan pengaruh nyata ter-hadap pertumbuhan dan kualitas buah cabai. Penyemprotan senyawa bioaktif dengan konsentrasi 1 ml/l dan 3 ml/l memberikan pengaruh yang sama dengan pemberian hormon sintetik yang dianjurkan yaitu 2 ml/l. Namun, rasio kandungan senyawa bioaktif IAA dan GA berbeda. Rasio kandungan senyawa bioaktif asal isolat B6.2 dan hormon tumbuh sintetik berturut-turut sebesar 1:131 dan 1:2.
Tabel 2. Pengaruh konsentrasi hormon tumbuh IAA dan GA asal isolat B6.2 terhadap tinggi tanaman cabai merah besar.
Perlakuan konsentrasi Tinggi tanaman (cm) Tinggi kumulatif
(cm)
30 hst 46 hst 61 hst 76 hst
Hormon tumbuh isolat B6.2*
1 ml 29,6±0,3 35,3b±0,2 55,1b±1,2 68,0b±0,5 2.120,5 3 ml 30,8±0,5 37,7c±1,0 56,9bc±0,5 70,7bc±1,1 2.214,5 5 ml 29,8±0,3 39,7d±0,7 58,2c±0,6 71,7c±0,9 2.264,5 Hormon sintetik** 2 ml 30,5±0,4 37,5c±0,6 56,8bc±0,6 69,1bc±0,9 2.195,5 Kontrol air 30,8±0,4 31,1a±0,5 51,8a±0,8 60,0a±0,5 1.955,5 *Kandungan bahan aktif IAA 0,49 ppm + GA 64,53 ppm. **Kandungan bahan aktif IAA 1,12 ppm + GA 2,41 ppm (Hormonik®).
Angka pada kolom yang sama diikuti oleh huruf yang sama menunjukkan tidak berbeda nyata berdasarkan uji Duncan pada taraf 5%.
Tabel 3. Pengaruh perlakuan hormon tumbuh IAA dan GA asal isolat B6.2 terhadap kualitas buah cabai merah besar.
Perlakuan Konsentrasi Jumlah buah Jumlah buah total Bobot buah (g) Kualitas I Kualitas II Kualitas III
Hormon tumbuh asal isolat B6.2*
1 ml 2,0b±0,2 8,0b±0,2 0,6b±0,0 10,9b±0,4 76,7b±0,3
3 ml 2,4bc±0,0 9,3cd±0,2 0,5a±0,0 1,2c±0,4 92,1c±0,4
5 ml 2,6c±0,0 9,8d±0,0 0,9c±0,0 12,9c±0,5 94,7d±0,3
Hormon tumbuh sintetik**
2 ml 2,3bc±0,2 8,9c±0,1 0,9d±0,0 12,1c±0,3 90,8c±0,4
kontrol 1,0a±0,2 5,8a±0,2 1,0e±0,0 7,8a±0,4 59,4a±0,4 *Kandungan bahan aktif IAA 0,49 ppm + GA 64,53 ppm. **Kandungan bahan aktif IAA 1,12 ppm + GA 2,41 ppm (Hormonik®).
Identifikasi Molekuler Isolat Bakteri Endofit B6.2 dengan 16S rRNA
Analisis homologi dan filogeni isolat bakteri endofit B6.2 berdasarkan 16S rRNA menunjukkan isolat B6.2 membentuk satu kelompok dalam genus Bacillus dan relatif dekat dengan B. vallismortis (Gambar 2). Hasil BLASTn juga menunjukkan isolat B6.2 memiliki homologi 100% dengan B. vallismortis (nomor aksesi LT838175.1). Gen 16S rRNA memiliki banyak posisi yang berevolusi secara bebas sehingga hubungan filogenetik sekelompok mikroba termasuk bakteri dapat diduga (Nobandegani et al. 2015).
KESIMPULAN
Analisis senyawa bioaktif hormon tumbuh asal isolat B6.2 menunjukkan bahwa kandungan campur-an IAA 0,49 ppm dcampur-an GA 64,53 ppm (dengcampur-an per-bandingan 1:31). Penyemprotan tanaman cabai merah besar dengan hormon tumbuh asal isolat B6.2 pada konsentrasi 5 ml/l meningkatkan tinggi tanam-an, bobot buah, dan bobot basah dan kering tajuk. Analisis molekuler menggunakan sekuen 16S rRNA menunjukkan bahwa isolat bakteri B6.2 diidentifikasi sebagai B. vallismortis dengan tingkat homologi 100%. Bakteri endofit isolat B6.2 berpotensi untuk di-kembangkan sebagai fitohormon pada tanaman cabai.
UCAPAN TERIMA KASIH
Penelitian ini didanai melalui DIPA Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian TA 2017–2018. Tim penulis mengucapkan terima kasih kepada Ir. Yadi Suryadi, M.Sc. dan Dr. Karden Mulya, M.Sc. yang telah memberikan bimbingan dalam penulisan naskah. Ucapan terima kasih disampaikan juga kepada teknisi Siti Aminah, Jajang, dan Pamuji yang telah membantu pelaksanaan penelitian.
KONTRIBUTOR PENULISAN
RYW: kontributor utama, perencana penelitian, analisis data. SF: kontributor anggota, perencana pe-nelitian. RI: kontributor anggota, perencana peneliti-an. IM: kontributor anggota, perencana penelitipeneliti-an. TPP: kontributor anggota, perencana penelitian. DNS: kontributor utama, penanggung jawab penelitian, analisis data.
DAFTAR PUSTAKA
Agustian, N., Maira, L., & Emalinda, O. (2010) Rhizobakteria penghasil fitohormon IAA pada rizosfer tumbuhan semak karamunting, titonia, dan tanaman pangan. Jurnal Solum, 7 (1), 49–60.
Ariyanti, Suherman, C., Anjasari, I.R.D., & Sartika, D. (2017) Respons pertumbuhan bibit nilam Aceh (Pogostemon cablin Benth.) klon Sidikalang pada
Tabel 4. Pengaruh penyemprotan IAA dan GA asal isolat B6.2 terhadap bobot tajuk dan bobot akar tanaman cabai merah besar. Perlakuan konsentrasi Bobot tajuk (g/tanaman) Bobot akar (g/tanaman)
Bobot basah Bobot kering Bobot basah Bobot kering Hormon tumbuh asal isolat B6.2*
1 ml 37,6b±0,7 7,7b±0,4 9,3b±0,4 1,1ab±0,6
3 ml 40,3b±1,5 8,1b±0,3 11,1c±0,5 1,3bc±0,1
5 ml 45,1c±0,3 9,1c±0,2 11,5c±0,4 1,4c±0,19
Hormon tumbuh sintetik**
2 ml 38,1b±1,6 8,0b±0,2 10,4bc±0,5 1,2ab±0,6
Kontrol 31,0a±0,9 6,9a±0,3 7,7a±0,5 1,0a±0,0
*Kandungan bahan aktif IAA 0,49 ppm + GA 64,53 ppm. **Kandungan bahan aktif IAA 1,12 ppm + GA 2,41 ppm (Hormonik®). Angka pada kolom yang sama diikuti oleh huruf yang sama menunjukkan tidak berbeda nyata berdasarkan uji Duncan pada taraf 5%.
Gambar 2. Dendrogram perbandingan isolat bakteri endofit terpilih dengan strain bakteri dari spesies Bacillus berdasarkan sekuen gen 16S rRNA pada basis data NCBI menggunakan metode Maximum Likelihood dengan boostrap 1.000 (Tamura et al. 2012).
0,005 100 NR074540 B. cereus NR041248 B. anthracis MK881067 Bacillus sp. MK880428 Bacillus sp. Sampel B6.2 LT838175 Bacillus vallismortis
media tanam subsoil dengan pemberian pati beras dan pupuk hayati. Jurnal Kultivasi, 16 (3), 394–401. Asyiah, I.N., Wiryadiputra, S., Fauzi, I. & Harni, R. (2015).
Populasi Pratylenchus coffeae (Z.) dan pertumbuhan bibit kopi arabika akibat inokulasi Pseudomonas diminuta L. dan Bacillus subtilis (C). Jurnal Pelita Perkebunan, 31 (1), 30–40.
Ayyaz, K., Zaheer, A., Rasul, G. & Mirza, M.S. (2016) Isolation and identification by 16S rRNA sequence analysis of plant growth-promoting azospirilla from the rhizosphere of wheat. Brazilian Journal of Microbiology, 47 (3), 542–550.
Bacon, C.W. & White, J.F. (2000) Microbial endophytes. New York, Marcel Dekker.
Badan Pusat Statistik (2016) Luas panen produksi dan produktivitas cabai. [Online] Tersedia pada: https://www.bps.go.id html [Diakses 31 Agustus 2018]. Badan Pusat Statistik (2019) Luas panen produksi dan
produktivitas cabai. [Online] Tersedia pada: https://www.bps.go.id html [Diakses 29 Agustus 2019]. Badan Standardisasi Nasional (1998) Syarat kualitas
tanaman cabai. SNI 01-4480-1998. Jakarta, Badan Standarisasi Nasional.
Campbell, C.L. & Madden, L.V. (1990) Introduction to plant disease epidemiology. New York, John Wiley and Sons.
Darojat, M.K. (2014) Pengaruh konsentrasi dan lama perendaman ekstrak bawang merah (Allium cepa L.) terhadap viabilitas benih kakao (Theobroma cacao L.).
Skripsi. Universitas Islam Negeri Maulana Malik Ibrahim.
Ernawati., Rahardjo, P. & Suroso, B. (2017) Respon benih cabai merah (Capsicum annuum L.) kadaluarsa pada lama perendaman air kelapa muda terhadap viabilitas, vigor dan pertumbuhan bibit. Agritrop, 15 (1), 71–83. Fathonah, D. & Sugiyarto (2009) Effect of IAA and GA3
toward the growing and saponin content of purwaceng (Pimpinella alpina). Bioscience, 1 (1), 17–22.
Fitriyah, L.A. (2015) Respon ketahanan biokimia 10 genotip cabai merah terhadap infeksi Colletotrichum sp. melalui induksi mikrob endofit. Skripsi. Institut Pertanian Bogor.
Gusmaini, Aziz, S.A., Munif, A., Sopandi, D. & Bermawi, N. (2013) Potensi bakteri endofit dalam upaya meningkatkan pertumbuhan, produksi, dan kandungan andrografolid pada tanaman sambiloto. Jurnal Penelitian Tanaman Industri, 19 (4), 167–177.
Hayati, E. (2010) Pengaruh pupuk organik dan anorganik terhadap kandungan logam berat dalam tanah dan jaringan tanaman selada. Jurnal Floratek, (5), 113– 123.
Istiqomah, D. & Trijoko (2014) Keefektifan bakteri endofit dalam meningkatkan pertumbuhan tanaman jagung secarain vitro. [Online] Seminar Nasional Dies Natalis ke-68 Fakultas Pertanian UGM. Yogyakarta, 13
September. Tersedia pada: https://www.
researchgate.net/publication/316188920_Keefektifan_ Bakteri_Endofit_dalam_Meningkatkan_Pertumbuhan_ Tanaman_Jagung_secara_In_Vitro [Diakses 29 Agustus 2019].
Kartikawati & Gusnaini (2018) Potensi bakteri endofit yang diisolasi dari tanaman jahe merah untuk memacu pertumbuhan benih lada. Buletin Penelitian Tanaman Rempah dan Obat, 29 (1), 37–46.
Kastono, D., Sawitri, H. & Siswandono (2005) Pengaruh nomor ruas setek dan dosis pupuk urea terhadap pertumbuhan dan hasil kumis kucing. Jurnal Ilmu Pertanian, 12 (1), 56–64.
Kuswandi, P.C. & Sugiyarto (2015) Aplikasi mikoriza pada media tanam dua varietas tomat untuk peningkatan produktivitas tanaman sayur pada kondisi cekaman kekeringan. Jurnal Sains Dasar, 4(1), 17–22.
Lane, D.J. (1991) 16S/23S rRNA sequencing. In: Stackebrandt, M. & Goodfellow, E. (eds.) Nucleic acid techniques in bacterial systematics. London, John Wiley and Sons, pp. 115–148.
Lestari, P., Susilowati, D.N., Samudra, I.M., Priyatno, T.P., Nugroho, K., Nurarfa, W., Setyawati, I. & Suryadi, Y. (2017) Keragaman genetik rizobakteri penghasil asam indol asetat berdasarkan 16S rRNA dan amplified ribosomal DNA restriction analysis. Jurnal AgroBiogen, 13 (1), 25–34.
Mehnaz, S. & Lazarovits, G. (2007) Inoculation effects of
Pseudomonas putida, Gluconacetobacter azotocaptans, and Azospirillum lipoferumon corn plant growth under greenhouse conditions. Microbial Ecology, 51, 326–335.
Mitreva, M. (2017) Infectious diseases: The microbiome in infectious diseases. In: Cohen, S.M., Powderly, J. &. Opal, W.G. (eds.) Infectious diseases. Fourth edition. China, Elsevier Limited, pp. 68–74.e2.
Munif, A., Wiyono, S. & Suwarno (2012) Isolasi bakteri endofit asal padi gogo dan potensinya sebagai agen biokontrol dan pemacu pertumbuhan. Jurnal Fitopatologi, 8(3), 57–64.
Munif, A., Wibowo, A.R. & Herliyana, E.N. (2015) Bakteri endofit dari tanaman kehutanan sebagai pemacu pertumbuhan tanaman tomat dan agen pengendali
Meloidogyne sp. Jurnal Fitopatologi, 11 (6), 179–189. NCBI (2019) Standard nucleotide BLAST. [Online] Available
from: https://blast.ncbi.nlm.nih.gov/Blast.cgi [Accessed 14 May 2019].
Nobandegani, M.B.J., Saud, H.M. & Yun, W.M. (2015) Phylogenetic relationship of phosphate solubilizing bacteria according to 16S rRNA genes. BioMed Research International, 5, 1–5.
Prajnanta (2001) Agribisnis cabai hibrida. Jakarta, Penebar Swadaya.
Puspita, F., Zul, D. & Khoiri, A. (2013) Potensi Bacillus sp. asal rizosfer Giam Siak Kecil Bukit Baru sebagai rhizobacteria pemacu pertumbuhan dan anti fungi
pada pembibitan kelapa sawit. [Online] Seminar Nasional Peranan Teknologi dan Kelembagaan Per-tanian dalam Mewujudkan Pembangunan PerPer-tanian yang Tangguh dan Berkelanjutan. Pekanbaru, Riau, November. Tersedia pada: https://repository.unri.ac.id/ xmlui/bitstream/handle/123456789/7405/1%20Fifi%20 Puspita.pdf?sequence=1&isAllowed=y [Diakses 29 Agustus 2019].
Puspita, F., Saputra, S.I. & Merini, J. (2018) Uji beberapa konsentrasi bakteri Bacillus sp. endofit untuk meningkatkan pertumbuhan bibit kakao (Theobroma cacao L.). Jurnal Agronomi Indonesia, 46 (3), 322– 327.
Rubio, M.G.T., Plata, S.A.V., Castillo, J.B. & Nieto, P.M. (2000) Isolation of Enterobacteria, Azotobacter sp. and
Pseudomonas sp., producers of indole-3-acetic acid and siderophores, from Colombian rice rhizosphere.
Revista Latinoamericana de Microbiologia, 42, 171– 176.
Salli, M.K., Ismael, Y. & Lewar, Y. (2016) Kajian pemang-kasan tunas apikal dan pemupukan KNO3 terhadap
hasil tanaman tomat. PARTNER-Buletin Pertanian Terapan, 12 (1), 215–227.
Saylendra, A. & Firnia, D. (2013) Bacillus sp. dan
Pseudomonas sp. asal endofit akar jagung (Zea mays
L.) yang berpotensi sebagai pemacu pertumbuhan tanaman. Jurnal Ilmu Pertanian dan Perikanan, 2 (1), 19–27.
Srinivasan, R., Karaoz, U., Volegova, M., MacKichan, J., Kato-Maeda, M., Miller, S., Nadarajan, R., Brodie, E.L. & Lynch, S.V. (2015) Use of 16S rRNA gene for identification of a broad range of clinically relevant bacterial pathogens. PLoS ONE. [Online] 10 (2), 1–22. Available from: https://doi.org/10.1371/journal.pone. 0117617 [Accessed 24 May 2019].
Sukmadi, R.B. (2012) Aktivitas fitohormon indole-3-acetic acid (IAA) dari beberapa isolat bakteri rizosfer dan endofit. Jurnal Sains dan Teknologi Indonesia, 14 (3), 221–227.
Susilowati, D.N., Mulya, K., Samudera, M.I., Priyatno, T.P., Suryadi, Y., Setyowati, M., Akhadiya, A., Sanjaya, R.A., Thontowi, A., Anggraini, S., Widyastuti, I., Pamuji, A., Kosasih, J., Aminah, S. & Achmad, A. (2017) Bioprospeksi mikroba penghasil senyawa bioaktif untuk peningkatan vigor dan ketahanan tanaman cabai terhadap antraknosa dan mikroba penghasil bioetanol. Laporan Hasil Penelitian 2017. Bogor, BB Biogen.
Syukur, M. (2007) Inheritance of resistance to anthracnose caused by Colletotrichum acutatum in chili pepper (Capsicum spp.). Buletin Agronomi, 35 (2), 112–117. Taghavi, S., Barac, T., Greenberg, B., Borremans, B.,
Vangronsveld, J. & Lelie, D.V.D. (2005) Horizontal gene transfer to endogenous endophytic bacteria from poplar improves phytoremediation of toluene. Applied and Environmental Microbiology, 71, 8500–8505. Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar,
S. (2012) MEGA6: Molecular evolutionary genetics analysis version 6.0. Molecular and Biology Evolution, 30 (12), 2725–2729.
Tien, T.M., Gasking, M.H. & Hubbel, D.H. (1979) Plant growth substances produced by A. brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Applied and Environmental Microbiology, 37, 1016–1024.
Tinendung, R., Puspita, F. & Yoseva, S. (2014) Uji formulasi
Bacillus sp. sebagai pemacu pertumbuhan tanaman padi sawah (Oryza sativa L.). Jurnal Online Mahasiswa Faperta Univ Riau. [Online] 1 (2), 1–5. Tersedia pada: https://jom.unri.ac.id/index.php/JOMFAPERTA/article/v iew/3024/2929 [Diakses 29 Agustus 2019].
Wahyudi, A.T., Astuti, R.P., Widyawati, A., Meryandini, A. & Nawangsih, A.A. (2011) Characterization of Bacillus
sp. strains isolated from rhizosphere of soybean plants for their use as potential plant growth for promoting rhizobacteria. Journal of Microbiology and Antimicrobials, 3 (2), 34–40.