Production and Utilization Technology
for Sustainable Development of Eaglewood (Gaharu)
in Indonesia
ITTO PD425/06 Rev. 1 (I)
MINISTRY OF FORESTRY OF INDONESIAIN COOPERATION WITH
INTERNATIONAL TROPICAL TIMBER ORGANIZATION IT OT
FRAGRANT WOOD GAHARU:
W H E N T H E
W I L D C A N
NO LONGER
P R O V I D E
Production and Utilization Technology
for Sustainable Development of Eaglewood (Gaharu)
in Indonesia
ITTO PD425/06 Rev. 1
(I)
MINISTRY OF FORESTRY OF INDONESIA IN COOPERATION WITH
INTERNATIONAL TROPICAL TIMBER ORGANIZATION
FRAGRANT WOOD GAHARU:
W H E N T H E
W I L D C A N
NO LONGER
P R O V I D E
Batu No. 5 Bogor, Indonesia; e-mail : [email protected] The place and date the report
was issued
: Bogor, July 1, 2011.
Disclaimer : Copyright @ 2011
This Proceeding is a part of Program ITTO PD425/06 Rev. 1 (I) : Production and Utilization Technology for Sustainable Development of Gaharu (Gaharu) in Indonesia
Published by : Indonesia’s Work Programme for 2011 ITTO PD425/06 Rev.1 (I) R&D Centre for Forest Conservation and Rehabilitation Jalan Gunung Batu No. 5 Bogor, Indonesia
Phone :62-251-8633234 Fax :62-251-8638111 E-mail : [email protected] ISBN : 978-979-3145-88-4
Cover by : Bintoro
Project number : PD425/06 Rev. 1 (I)
Host Government : Indonesia Name of the Executing
Agency
: Forestry Research and Development Agency (FORDA)
Project Coordinator Dr. Ir. Maman Turjaman, DEA Starting date of the project : May 1, 2008
The importance of gaharu (eaglewood or aloewood or jingkoh or oudh) for many users has long been recognized. Gaharu is also considered the world’s most valuable incence with even higher price for high quality gaharu. Due to its multiuses, demand for gaharu products continues to increase significantly and may cause rapid depletion of gaharu trees in the wilds. Natural habitat of gaharu suffers from uncontrolled exploitation, and as the consequences, some important gaharu-producing trees under a serious degradation. People who live surrounding the forests are the ones who are affected directly from the rapid depletion of gaharu because their livelihood depends on the forest.
On the other hand, forests are now receiving more and more attention from international society because profound appreciation on the function of forest has increased over the years. Forests are no longer seen as a place for timber production only, but also for many non-timber forest products. More importantly, forest is seen a lot as as environmental service provider nowadays. Indonesia is the world’s third largest area of tropical forest, being endowed with nearly 90 million hectares under forest cover. With today’s international focus on climate change, the forests have become the assets that contribute significantly to the country’s income.
It is thus timely to promote sustainable production of gaharu as an important strategy for conserving natural gaharu tree species, thus the forest habitats, and concurrently fulfilling the demand for gaharu products from cultivation. This project of ITTO PD425/06 Rev.1 (I): “ Production and Utilization Technology for Sustainable Development of Eaglewood (Gaharu) in Indonesia” is targeting to achieve the goals. Technology for accelerating gaharu production is intensively studied and several gaharu cultivation plots have been established in several locations in Indonesia. It is our aim to alleviate poverty of particularly forest community by providing simple technology for gaharu production as source of income and to stimulate cultivation of gaharu plants as their valuable backbone commodity.
The objective of this book is to give thorough information concerned with gaharu, and summarize the findings of the state-of-the-art research on gaharu. A key message of this book is to stimulate an understanding that the future of gaharu relies solely on sustainable production of gaharu and habitat conservation, and that technology intervention plays a major role in the process.
Adi Susmianto
Head, R & D Centre for Forest
Conservation and Rehabilitation
PREFACE ...iii
LIST OF CONTENTS ... v
LIST OF TABLE ...vii
LIST OF FIGURE ... ix
1. INTRODUCTION...
1
2. KNOWING SPECIES THAT PRODUCE GAHARU ...
3
3. SILVICULTURE OF GAHARU PLANT ...
15
3.1 Propagation using seeds, cuttings and tissue culture ...
15
3.2 Inoculation of beneficial microbial to promoted plant growth ...
16
3.3 Pest and Disease ...
16
3.4 Soil characteristics ...
19
4. GAHARU BIOINDUCTION TECHNOLOGY ...
23
4.1 Deliberate tree wounding using mechanical tools ...
23
4.2 Deliberate tree drilling and chemical injection ...
23
4.3 Deliberate tree drilling and inoculation of fungal inoculum ...
24
5. CHEMICAL PROPERTIES OF GAHARU ...
31
6. GAHARU PRODUCTS AND TRADING ...
39
7. CONSERVATION AND SUSTAINABLE PRODUCTION OF GAHARU ...
49
8. EXIT STRATEGY ...
51
8.1 The Role of Institution ...
51
8.2 Master Plans ...
53
9. CONCLUDING REMARKS ...
57
REFERENCES ...59
LIST OF TABLE
Table 1. Scientific names, synonyms, common names of Aquilaria and Gyrinops
and distribution ...3
Table 2. Pests and Diseases of gaharu plants in several locations in Indonesia ...16
Table 3. Soil physical characteristics of three gaharu plantation sites in Java Island, Indonesia ...20
Table 4. Soil chemical characteristics of three gaharu plantation sites in Java Island, Indonesia ...20
Table 5. Fungi isolated from infected tree identified based on their morphological characteristics and experiments related with the fungal inoculation ...24
Table 6. Experimental plot of gaharu trees induction by deliberate tree drilling and Fusarium spp. injection in several locations across Indonesia ...26
Table 7. Molecular identification of 36 strains of gaharu-inducing fungi collected from 17 provinces in Indonesia ...29
Table 8. Phenol compounds present in the induced gaharu products ...34
Table 9. Classification of gaharu quality in Samarinda (East Kalimantan) ...41
Table 10. Criteria and classification of gaharu quality ...42
Table 11. Selling price of gaharu in markets of Samarinda, in East Kalimantan ...42
Table 12. Classification of gaharu quality according to Indonesian National Standard (SNI) ...43
Table 13. Development of quota and realization related to the eagle wood export from Indonesia ...45
Table 14. Several institutions/stakeholder who will carry out the exit strategy following the ITTO’s PD 425/06 Rev. 1 (I) project ...52
LIST OF FIGURE
Figure 1. Seeds of gaharu. (1-2): Aquilaria malaccensis; (3-4): Gyrinops versteegii. ...5
Figure 2. Gaharu trees. ...6
Figure 3. Flowering phenology of Aquilaria spp. in natural forest, plantation and Botanical Garden (a) A. malaccensis and A. microcarpa in West Kalimantan; (b) A. microcarpa in East Kalimantan; (c) A. malaccensis, A. microcarpa,
and A. beccariana in Botanical Garden; (d) A. crassna, A. malaccensis
and A. microcarpa in plantation, Bogor; (e) A. filarial in plantation, Bogor; (f) A. hirta in plantation, Bogor (Source: Suhartono and Newton, 2001). ...7
Figure 4. (1): Aquilaria malaccensis La,. 1. twig, 2 flower, 3. longitudinal section of flower, 4. fruit, 5. longitudinal section of fruit. (Source: Plant Resources of South East Asia 19) and (2): Gyrinops ladermannii. a- branchlet habit; b- flower bud (left), opened flower (right); c- seed dorsal view (left), ventral view (right); d- dehisced fruit emerging from lateral slit of floral tube with one seed hanging out on funicle; Herbarium specimen Zich 315, CANB Accession Number 531408. Botanical illustration : Sharyn Wragg. (Zich and Compton 2001 in Dunn et al., 2003). ...12
Figure 5. Silviculture of gaharu.(1-2): Propagation by cuttings with KOFFCO system; (3): Effect of inoculation with beneficial microbes for promoting growth; (4): Gaharu plantation. ...18
Figure 6. Pests and disease of gaharu trees. (1): Leaf eater Heortia vitessoides;
(2): Stem Borer; (3): Colony of ants, the predator of H. vitessoides: (4):
Hearth-rot fungi. ...19
Figure 7. An illustration of induction procedure for stimulation of gaharu formation. (1): Tree drilling to make about 5 mm diameter hole with 25 cm space in between holes; (2-3): one ml of liquid inoculum is injected with a syringe; (4): one month after inoculation, the efficiency of induction is observed by peeling a tree bark to observe the disease symptom (Source: http://www. trubus-online.co.id/mod/publisher/media/465.jpg. ...27
Figure 8. Technology of fungal induction to stimulate gaharu production.
(1): Bamboo stagger for climbing tree for inoculation; (2): Drilling the tree and making holes for inoculation; (3): Fusarium sp., gaharu-inducing fungi grown on agar plate; (4): Wood coloration on stem tissue, an
indication of resin production. ...28
Figure 9. Harvesting procedure of gaharu product from deliberate tree injection with fungal inoculant. (1): Initial symptom of gaharu formation on stem;
chromones, (2): C19H18O5, (3): C1H14O4, (4):
6-hydroxy-2-[2-(4-hydroxyphenyl)ethyl]chromone, (5): dihydroxyl derivative of 2-(2-pheny-lethyl)-chromone, (6): C1H14O3, and (7): C18H16O4 (Source: Konishi
et al., 2002). ...32
Figure 11. Chromatogram revealing constituents in 6-month old induced gaharu ...33
Figure 12. (4) Figure 1. Chemical content of gaharu originated from natural process and artificial induction. (1): Natural gaharu of West Kalimantan origin; (2): Gaharu from deliberate inoculation with isolate from West Kalimantan; (3): Natural gaharu of Gorontalo origin; (4): Gaharu from deliberate inoculation with isolate from Gorontalo (Source: Santoso et al., unpublished data). ...36
Figure 13. Structures of compound from gaharu oil ...37
Figure 14. Major constituent genkwanin 5-O-b-primeveroside compound that
caused mild laxative effect in mice (Source: Hara et al., 2008). ...38
Figure 15. Gaharu products. ...46
Figure 16. Other gaharu products ...47
Figure 17. Gaharu classes of quality: (a) Tanggung; (b) Kacangan ; (c) Teri and
(d) Kemedangan. ...47
Figure 18. Several pieces of gaharu still in logs which are ready for export. ...48
Figure 19. Flow-scheme regarding the exit strategy of gaharu development that
INTRODUCTION
Gaharu (agarwood, eaglewood, aloeswood, jinkoh, or oudh) has long been appreciated for its multipurpose uses, range from incense for religious and traditional ceremonies, perfume, medicine and ornamental functions in many countries. The occurrence of this-so-called the wood of the gods has been strongly surrounded by myths and history. Gaharu use is mentioned in the Old Testament as ‘aloe’ or ‘ahaloth’ in Psalm 45:8. Gaharu is the only tree in the Eastern myth that has been descended to Man from Eden garden (Duke, 2008). In Egypt and Japan, gaharu was used to embalm dead bodies. In India and Cambodia, it is used for traditional and religious ceremony.
The resin compound of gaharu is highly commercial. Resin impregnated in the heartwood a number of gaharu-producing species is due to fungal infection. Two mostly known genera are Aquilaria and Gyrinops that are native to Southeast Asia with Indonesia, Malaysia, Vietnam, Cambodia, Thailand, Laos and Papua New Guinea being the main producing countries, and Singapore being the central trade country (Persoon, 2007).
Natural forests have been the main resource for gaharu collection for many years. However, gaharu hunters usually cut down the whole trees to find the resin and this practice has diminished gaharu population in the wilds and consequently has led gaharu-producing tree species under a threat of extinction. Major harvesting of gaharu was recorded between the 1980s and early 1990s in East Kalimantan caused by high demand for gaharu and was due to diminishing supply from countries like Vietnam and Cambodia (Barden et al.,2001 in Gunn et al., 2003). This excessive hunting activity has caused significant reduction of wild gaharu stocks within a short period of time. Similar activity also occurred in Papua after gaharu hunters landed in 1996 that has led to an ending of gaharu harvesting from its natural habitat (Persoon, 2007). 1998 was the first officially recorded year for gaharu discovery and harvesting in Yapsiei, May River and Ama villages in West Sepik, Papua New Guinea (Gunn et al., 2003). Harvesting of gaharu in these countries involve professional and traditional collectors. Professional collectors sponsored by Chinese and bogus trades were sometimes dropped by helicopters to hunt for gaharu (World Wide Fund for Nature, 1999).
In November 1994, Aquilaria malaccensis Lamk. was initially listed in CITES (the Convention on the International Trade in Endangered Species of Wild Flora and Fauna), Appendix II to prevent this species from extinction. However, continual excessive gaharu exploitations have then put two genera Aquilaria and Gyrinops in CITES, Appendix II, only ten years later. CoP13 Prop.49 (TRAFFIC, 2004) listed 24 species of the genus
Aquilaria and seven species of the genus of Gyrinops. CITES regulates the permitted quota for gaharu export in order to sustain gaharu existence, and yet, Indonesia, in particular, has not been able to meet the quota because it has become more difficult to collect gaharu from its natural habitat because they are dissapearing.
KNOWING SPECIES
THAT PRODUCE GAHARU
Aquilaria and Gyrinops are the two most important gaharu-producing genera, within the family of Thymelaeaceae (Order: Myrtales and Class: Magnoliopsida). There are slight differences in reports on the number of species within each genus. TRAFFIC-CITES-CoP13 Prop.49 (2004) recorded 24 species belong to the genus Aquilaria and 7 species belong to the genus of Gyrinops (Table 1). On the other hand Ding Hou (1960
in Gunn et al., 2004), reported there are 12 species belonging to the genus Aquilaria
and 8 species belonging to the genus Gyrinops.
These trees naturally occur in at least 12 countries: Bangladesh, Butan, Cambodia, Indonesia, Lao PRD, Malaysia, Myanmar, Philippines, Thailand, Vietnam and Papua New Guinea (Barden et al. in Gunn et al., 2004).
Compton (2002) reported three gaharu-producing species originated from Papua Island. Two species, A. filaria and Gyrinops versteegii are found in Papua (formerly Irian Jaya) and the only species, G. ledermannii is from Papua New Guinea (PNG, TRAFFIC PC12 Doc.8.3, 2002). This PNG gaharu species was firstly found and harvested in approximately 1998 for its resinous wood in the Yapsiei, May River, and Ama Villages in West Sepik (Gunn et al., 2004). The first reported taxonomic work determined the species as Gyrinops ledermannii. Later report by Gunn et al. (2004) however suggested five of the eight species Gyrinops, have been found in New Guinea island: G. ledermannii, G. caudate, G. podocarpus, G. salicifolia and G. verstegii. Except G. walla found in Sri Langka, the other seven are distributed east of the Wallace Line which is a transitional biogeographical zone that marks Asia zone to the west and Australian zone to the east.
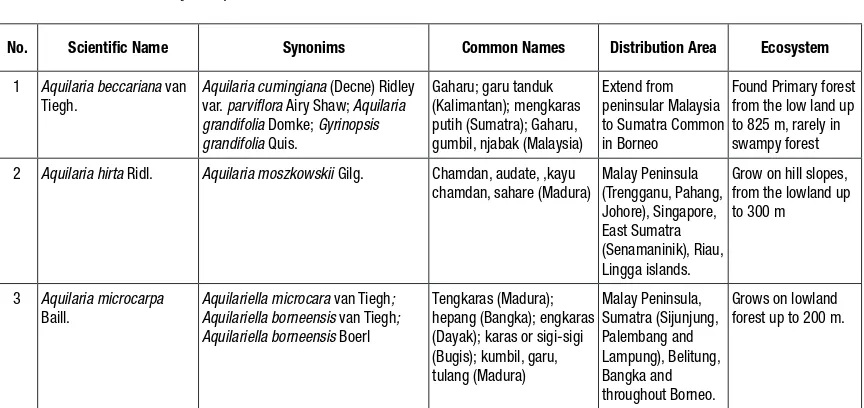
Table 1.
Scientific names, synonyms, common names of Aquilaria and Gyrinops and distributionNo. Scientific Name Synonims Common Names Distribution Area Ecosystem
1 Aquilaria beccariana van Tiegh.
Aquilaria cumingiana (Decne) Ridley var. parviflora Airy Shaw; Aquilaria grandifolia Domke; Gyrinopsis grandifolia Quis.
2 Aquilaria hirta Ridl. Aquilaria moszkowskii Gilg. Chamdan, audate, ,kayu chamdan, sahare (Madura)
No. Scientific Name Synonims Common Names Distribution Area Ecosystem
4 Aquilaria cumingiana
(Decne) Ridl.
Gyrinopsis cumingiana Decne; Decaisnella cumingiana O.K.; Gyrinopsis cumingiana var. Pubescens Elm.; Gyrinopsis decemcostata Hall.f.; Gyrinopsis pubifolia Quis.
5 Aquilaria audate (Oken) Merr.
Aquilaria filaria (Oken) Merr., Gyrinopsis brachyantha Merr.,
Cortex filarius Rumph., Pittosporum ferrugineum var. filarium DC.,
Pittosporum filarium Oken,
Aquilaria tomentosa Gilg, Gyrinopsis bracyantha Merr.. Gyrinopsis acuminate Merr. A. _audate_e Quis.J.
Agé (Sorong), bòkuin
Gyrinopsis brachyantha Merr. - Luzon: Cagayan Province
Gyrinopsis urdanetensis Elmer Mangod, makolan (Mbo) Mindanao: Mt. Urdaneta
Gyrinopsis citrinaecarpa Elmer Agododan (Mbo) Mindanao on moist compact soil of forested ridges, about 1300 m.
9 Aquilaria apiculata Elmer - - Mindanao: Bukidnon
province
in dry and mossy forest at 1100-1800 m.
10 Aquilaria parvifolia (Quis.) Ding Hou
- - Luzon on forested slopes at 1000 m.
11 Aquilaria rostrata Ridl. - - Malay Peninsula
(Pahang, Gunung Tahan).
12 Aquilaria crassna Pierre ex Lecomte
15 Aquilaria subintegra Ding Hou
- - Thailand
16 Aquilaria grandiflora Bth. - - China.
17 Aquilaria secundana D.C. - - Moluccas
18 Aquilaria moszkowskii
Gilg
- - Sumatra
19 Aquilaria tomentosa Gilg - - New Guinea
20 Aqularia bailonii Pierre ex Lecomte
- - Cambodia
21 Aquilaria sinensis Merr. - - China.
22 Aquilaria apiculata Merr. - - Philippines
No. Scientific Name Synonims Common Names Distribution Area Ecosystem
25 Gyrinops versteegii (Gilg) Domke
Gyrinops wala (non Gaertn.) Koord.;
Branchythalamus versteegii Gilg;
Buru and Halmahera in rain-forest
27 Gyrinops decipiens Ding Hou
29 Gyrinops salicifolia Ridl. - - Western New
Guinea (Utakwa, Nabire)
in fringing rain-forest, 300 m
30 Gyrinops _audate (Gilg) Domke
Aquilaria podicarpus Hall.f.; Gyrinops ladermanii (non Donke) Merr & Perry
Figure 2.
Gaharu trees.Gaharu-producing species is characterized by small flower similar to that of jasmine and the fruit is bitter (Figure 1). The fruit does not fall when it is ripe, but it tossed the seeds to the surrounding ground (Prosea, 1999). Gaharu tree was firstly introduced by Arabic in Aceh, Sumatra. From here, the tree spread to the southern part of Sumatra, Kalimantan and Papua. In 1991, gaharu tree was found in Kayan Mentarang National Park, East Kalimantan.
Gaharu is commonly found in primary and secondary forest at 850m above sea level (asl) with 1,500 – 6,500 mm/year with 14-28ºC (Prosea, 1999). Gunn et al. (2004) reported the range of gaharu occurrence is about latitude 27°N to 10°N30’S and longitude 75°E to 149°E.
The most important genus, Aquilaria is typical of understorey species (Figure 2).
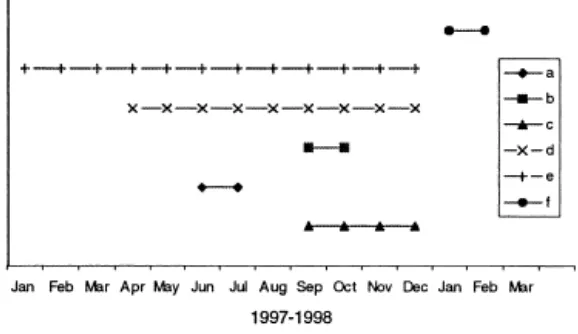
Aquilaria malaccensis, A. beccariana, A. crasna, A. filarial, A. hirta, A. microcarpa are of six Aquilaria species native to Indonesia (Ding Hou, 1960 in Suhartono and Newton, 2001). These six species are primarily found in lowland and upland Sumatra, Kalimantan, Maluku and Papua (Suhartono and Newton, 2001).
Generally, flowers grew on terminal branches among the foliage, but a few flowers of
of flowering to produce seed takes about three months in plantation area (Suhartono and Newton, 2001). Further Suhartono and Newton (2001) noted that smaller trees of
A. malaccensis and A. microcarpa produced more seeds than larger and older ones. Seeds of Aquilaria had no dormancy (recalcitrant) and will germinate soon after seed maturation.
Figure 3.
Flowering phenology of Aquilaria spp. in natural forest, plantation and Botanical Garden (a) A. malaccensis and A. microcarpa in West Kalimantan; (b) A. microcarpa in East Kalimantan; (c) A. malaccensis, A. microcarpa, and A. beccariana in Botanical Garden; (d) A. crassna, A. malaccensis and A. microcarpa in plantation, Bogor; (e) A. filarial in plantation, Bogor; (f) A. hirta in plantation, Bogor (Source: Suhartono and Newton, 2001).Upon ripening, fruits of Aquilaria spp. split loculicidally in half from the apex, and the seeds are hanging on thin threads for about one hour before the threads break and the seeds are scattered (CIFOR, 1996 and Ding Hou, 1960 in Suhartono and Newton, 2001).
Below is specific description of species belonging to mostly the genera Aquilaria
and Gyrinops as well as other less important gaharu-producing species.
1. Aquilaria malaccensis Lam
a. Distribution and Habitat
Aquilaria malaccensis has been found in India, Burma, Malaysia, Phillipines and Indonesia. In Indonesia, it is found mainly in Sumatra (Sibolangit, Bangka, Jambi, Riau and South Sumatra), Kalimantan, Sulawesi, Moluccas and Papua.
Aquilaria malaccensis has been found in primary and secondary forests, mainly in lowland and on hillsides at altitude of 200-750m asl. It grows well in sandy soils and areas having Koeppen climate type A-B with temperatures of 14-32ºC and annual rainfall of 2,000-4,000 mm.
b. Botanical description
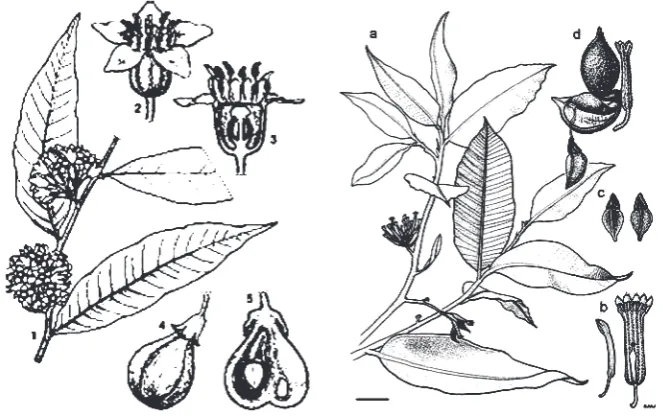
Aquilaria malaccensis reaches up to 20 – 40 m tall and 60 cm in diameter. Young bark is light brown with fine hairs, older bark is smooth and whitish in color. Without resin, wood is white, light and soft, however resinous wood is hard, dark and heavy. Leaves are characterized by alternate, elliptic or lanceolate, 3-3.5 cm wide and 6-8 cm long with 12-16 pairs of veins. Inflorescens a terminal or axillary umber. Flowers hermaphroditic, up to 5 mm long, fragrant and yellowish green or white (Figure 4).
Fruit is green in color, egg-shaped capsule, leathery exocarp with fine hairs, 4 cm long and 2.5 cm wide. There are two seeds per fruit. Seed is blackish brown incolor, ovoid, and densely covered with red-brown hair. There are approximately 1,500 seeds per kg. At the age of 5-6 years, the tree starts flowering and fruiting, and medium size tree is reported to produce about 1.5 kg of seed during good seed years. Seasons for flowering and fruiting are dry season. In Sumatra, flowering and fruiting season is twice a year. The seasons are July-August and March-April for flowering which have mature fruits in November-December and July-August, respectively. Mature fruits have blackish brown colour and they should be collected directly from the tree (Suhartono and Newton, 2001).
2. Aquilaria crassna
This species is the most widely known gaharu species from Indochina (Adelina, 2004). The Vietnamese government has enlisted A. crassna as endangered (Burfield, 2003).
Aquilaria crassna is found widely in Thailand (Nobuchi and Siripatanadilok, 1991). Flowering season in plantation area, 2 km north of Bogor occurs April to December (Suhartono and Newton, 2001). In general, seeds germinate 9 – 15 days after sowing and this species had the highest germination success (92%) compared with the other 5 species (Suhartono and Newton, 2001).
3. Aquilaria microcarpa
4. Aquilaria beccariana
Flowers are green to yellowish in color. Flowering season in Botanical Garden occurs between September and December. In general, seeds germinate 9 – 15 days after (Suhartono and Newton, 2001).
5. Aquilaria filaria
Flowers are white to yellowish green in color. Flowering season in plantation area, 2 km north of Bogor occurs all year around. In general, seeds germinate 9 – 15 days after sowing and this species had the lowest germination rate (53%) compared with the other 5 species (Suhartono and Newton, 2001).
6. Aquilaria hirta
Flowers are white in color. Flowering season in plantation area, 2 km north of Bogor occurs during rainy season in January. Seeds start to germinate 9 day after sowing (Suhartono and Newton, 2001).
7. Gyrinops versteegii (Gilg.) Domke
a. Distribution and Habitat
Gyrinops versteegii (Gilg.) is found in eastern part of Indonesia: Sumbawa, called
Seke, in East Alor, Lombok, Flores, Sumba, and Papua (Mulyaningsih and Yamada, 2007).
In Sumbawa, G. versteegii grows on the hillside (400-800m above sea level) ranging from Tartar village (Doro Tambiung Mountain) in West Sumbawa to Lambu village (Doro Saboke Mountain) in East Sumbawa. These plants grow in secondary and primary forests with high humidity, with Ficus sp., Eugenia spp., Garcinia sp.,
Calophyllum spp., Maranthes corymbosa Bl., Sterculia foetida L., Schleichera oleosa (Lour.) Oken., etc. (Mulyaningsih and Yamada, 2007).
In Sumba, this plant grows on brown, thin humus soil found in primary forest in Riwuta and on the foot of Meja Mountain, West Sumba. In Alor, this plant was planted in field with Erytrina sp., and the soil around the seedlings was mulched with rice straw and sprinkled well with water.
b. Botanical description
In Sumbawa and Alor Island, this species grows as shrub, 1-4m height, 1-10 cm diameter, while in Flores Island, it grows as trees.
In Flores the the tree is found to have 10-17½ m height, 25-30 cm diameter. Young branchletspubescent, bark gray. Leaves chartaceous to subcoriaceous, pubescent, on the nerves and veins beneath, glabrescent or glabrous, dull and light green in beneath, shinning and dark green above, elliptic-oblong, ovate-oblong, or obovate-ovate-oblong, 8-15 by 1½-5 cm; base cuneate, apex up to 2 cm narrow- 366 acuminate; nerves and veins similar, numerous, slightly oblique and parallel; petiole short, 3-5 mm, pubescent. Fruits yellow or orange, slightly obovoid or ellipsoid, 2-2¾ by 1-1½ cm, shortly acuminate to the apex, attenuate to the base. Seed ovoid, Plano convex, 9 by 6 mm, with a caruncle-like appendage at the base, ± 2 mm thick.
In West Sumba, shrub or tree of this species is found, 2 m up to 25 m height, diameter 3 cm up to 40 cm. Young branchlets pubescent, bark gray. Leaves
chartaceous to subcoriaceous, pubescent, on the nerves and veins beneath, glabrescent or glabrous, dull and light green in beneath, shinning and dark green above, elliptic-oblong, ovate-oblong, or obovate-oblong, 8-15 by 1½-5 cm; base cuneate, apex up to 2 cm narrow-acuminate; nerves and veins similar, numerous, slightly oblique and parallel; petiole short, 3-5 mm, pubescent.
8. Gyrinops decipiens Ding Hou
a. Distribution and habitat
This plant was found in primary forest with thin humus on the side and the top of the Ganda Dewatan mountain in Buttu Ada and Salusampe, Salubaka and Tampakura villages, Mamuju, the Tapusaang mountain in Karama village, Mamasa, and the Kapusaan mountain and the Tunggumanu mountain in Karosa in West Celebes. Gyrinops decipiens was also found in Kulawi, Tuwulu village, the Ulu Karosa river, Tembok Jerman and Lengke mountains around the Towuti Lake in Central Celebes; and in mountains in North Luwu in South Celebes (Mulyaningsih and Yamada, 2007).
b. Botanical description
Shrub to tree, 2 m up to 17 m height, diameter 3 cm up to 30cm. Branchlet fissure shallow to deep, light to dark gray, glabrous to pubescent. Leaves chartaceous to subcoriaceus, above glabrous, below pubescent scattered on the vena, shining on both surfaces, elliptic-oblong, lanceolate 7.5-23.5x 2.6-6.8 cm; base acute-acuminate, base caudate (0.52.0 cm long). Vena parallel, 23-39 pairs, elevation visible on below and obscure on above. Inflorescent axilar or terminal on the short branchlet in the axilar, umbel, consisting of 1-6 flowers, pedicle 2-5 mm, pubescent, brachtea opposite on the base of pedicelus, a crescent, rounded and thick on the apex of pedicle; pedicelus 1-3 mm. Flower like club, calyx tube pubescent outside, 4-6x2 mm long, calyx lobe 1,5x1 m. Fruit ovoid-oblong, 1-1.5x0.8-1.3 cm, color orange when fruit mature, two locus, 1-2 seeds each fruit. Stipes 7 mm, emerges on the base to mid of calyx lobe. Seed planocovex, 6x(5-7)mm, caruncle 5 mm. Flowering and Fruiting season on July-August.
Mulyaningsih and Yamada (2007) divided this plant into two varieties, i.e., gaharu beringin and gaharu cabut. Gaharu beringin or Gyrinops decipiens var. microphylla
Mulyaningsih & Yamada var. nov. Character: tree, fissured bark deep. Leaves
in specific wood. Since 2003, in some villages such as Dara, Maepi and Lere, North Luwu (South Celebes) and Tampalopo and Tampakura villages, Mamasa (West Celebes) cultivated this species. Gaharu cabut or Gyrinops decipiens var. macrophylla
Mulyaningsih & Yamada var. nov. Character: Shrub, fissured bark shallow. Leaves
chartaceous, more weigh, elliptic-oblong, 14.5-23.5x6.0-6.8 cm, vena 36-39 pairs,
gaharu forming in whole wood tissue of plant.
9. Gyrinops salicifolia Ridl.
a. Habitat and distribution
Gyrinops salicifolia grew in Dosay village, Sentani, Papua. This plant was cultivated by a person as decoration plant, because of its good canopy and leaves. Gaharu from Sentani and Jayapura was taken from this hill (Mulyaningsih and Yamada, 2007).
b. Botanical description
Slender shrub, up to 2 m. Branchlet light brown, pubescent. Leaves sparsely pubescent on the midrib and sometimes on the nerves and veins beneath, lanceolate to linear-lanceolate, 1½-10 by 1/
5-1 cm; base cuneate apex acuminate
and pointed; nerves and veins similar and equally strong, slightly visible beneath, obscure above; petiole ⅓-½ mm.
10. Gyrinops ladermannii Domke
a. Habitat and distribution
Gyrinops ladermannii resembles G. salicifolia, but it grows as shrub of small trees with 7-10 m high and 13-15 cm in diameter. The leave is broader, the angle of leave and ranchlet is larger (Figure 4). Branchlet has darker color, and the wood is harder. This plant grows in secondary forest with Callophylum sp. on lime soil with thin humus. It was found on a hill in Maribau village, 50-200 m above sea level, Sentani, Jayapura, Papua (Mulyaningsih and Yamada, 2007).
b. Botanical description
Leave subcoriaceous, obovate-oblong to lanceolate, 6½-12 kali (1½) 2½-5 cm; pubescent scattered on vein and midrib beneath and glabrous above. Base acute-cuneate, apex acuminate to caudate, nerves spread, visible, dense, curve, ascending face to tip. Inflorescentia pseudo lateral or terminal, sessile, consisted of 2-3 flowers, pedicellus thin, 3-5 mm. Calyx tube cylinders, 13 mm long, diameter 1½ mm. Calyx lobe ovate, 1½-2 kali ½ mm, outside acute, pubescent, inside obtuse, tomentose. Petaloid square, 3/
5x ½ mm, obtuse, villous. Stamen sessile,
oblong, 1-1¼ x 1/
5 mm. Fruitpyriform 1¾ x ⅓ cm (included stipe 3 mm and apex
up to 4 mm acuminate or caudate), pannose, irregular, wrinkled to transversal.
Figure 4.
(1): Aquilaria malaccensis La,. 1. twig, 2 flower, 3. longitudinal section of flower, 4. fruit, 5. longitudinal section of fruit. (Source: Plant Resources of South East Asia 19) and (2): Gyrinops ladermannii. a- branchlet habit; b- flower bud (left), opened flower (right); c- seed dorsal view (left), ventral view (right); d- dehisced fruit emerging from lateral slit of floral tube with one seed hanging out on funicle; Herbarium specimen Zich 315, CANB Accession Number 531408. Botanical illustration : Sharyn Wragg. (Zich and Compton 2001 in Dunn et al., 2003).11. Gyrinops caudate (Gilg.) Domke
a. Habitat and distribution
Gyrinops caudate was found in Agat, Mappi and Boven Digul and Merauke, West Papua. It grows in primary forest in the swamp, 5-20 m above the sea, among the sago plant, the most common soil type is sandy clay over clay. Cultivation was found in Aboge and Ecy village, District Assue, Mappi (Mulyaningsih and Yamada, 2007).
b. Botanical description
Shrub or tree up to 17 m by 36 cm. Branchlets grayish, whitish pubescent and glabrescent. Leaves chartaceous, glabrous, dull beneath and shining above, elliptic-oblong, ovate-oblong, rarely lanceolate, 6-13 by 1½-4 cm; base cuneate; apex up to 1½ cm, acuminate; nerves and veins scarcely distinguishable, numerous, parallel, visible beneath, obscure above; petiole ± 3 mm. Inflorescences axillary or on the terminal of short branches, 12-18 flowers, peduncle 2-8 mm. Flowers
c. 5 mm pedicelled 5-7 mm, floral tube copular, 3-4 mm long, calyx lobes oblong, 1 mm long, petaloid appendages transverse oblong, c. ½ mm long; stamen subsessile, slightly longer than the appendages. Ovary ovoid, densely pillose; style very short; stigma capitate. Fruit protunding from the flower, rhombicus-oblong, contricted from the base to apex, pubescent, 21/
5 cm included 5 mm stipes,
12. Less important gaharu species
a. Excoecaria agallocha L. (Euphorbiaceae)
Exocoecaria agallocha is called gaharu buaya (crocodile gaharu) because its gaharu production smells like fired wood and less fragrance. The smoke can irritate eyes (Mulyaningsih and Yamada, 2007). The price is usually low and the product is used for mixing material of joystick (hio) or incense. This plant can be found along beach and composes mangroves in Somau Island.
b. Wikstroemia androsaemifolia Decne i. Distribution and habitat
This species is found in primary forest in Meja Mountain. Gaharu from
Wikstroemia androsaemifolia, known as male gaharu or red gaharu is less preferable because of its spicy smell and the smoke irritates eyes (Mulyaningsih and Yamada, 2007).
ii. Botanical description
Shrub, young branchlets slightly flattened at the nodes, densely appressed pubescent, glabrescent. Branches terete, reddish brown, glabrous; axillary buds densely covered with golden-coloured hairs. Leaves papery, glabrous, rarely sparsely hairy on the lower surface and especially on the nerves and veins of young leaves, in dry state light-greenish, light brown or greenish-brown to brownish and shining on the upper surface, pale greenish, light-yellowish-green or light-brown and dull on the under surface, elliptic or ovate-oblong, 1¾-5½(8-) by ¾-2½(-4) cm; base acute, apex acute to narrow-acute, very rarely obtuse; nerves 8-11 pairs, elevated below and slightly depressed above, obliquely spreading towards the margin and the curved upward; veins almost as distinct as the nerves, loosely reticulate beneath, obscure above; petiole ± 2mm.
c. Phaleria capitata Jack
Phaleria capitata grows in Kapusaan Puncak and Tiga Puluh Puncak Mountains, Mamasa, West Celebes and in some villages around the Towuti Lake in Central Celebes.
This species is also called gaharu buaya and less preferable because the fragrance is not good. The gaharu is used as a mixture in the making of joystick and for craft (Mulyaningsih and Yamada, 2007).
d. Phaleria microcarpa
Phaleria microcarpa was found in Papua with local name ‘gaharu puk-puk’. It produced smoky, bitter and unpleasant fragrant when burnt. It does not have commercial value. The bark was used to create bilums (traditional woven carrying bags) in the upper Sepik, Papua (Mulyaningsih and Yamada, 2007).
e. Phaleria nisdai Kanehira i. Habitat and distribution
glabercent on the both surfaces, dull beneath, nerves 5-9 pairs, elevation, decendent, veins reticulate, beneath distinct, obscure above. Inflorescences terminal and or at a long axils of branchlets, umbelliform, 10-12 flowers, one peduncle each nodes, peduncles 6-8 mm, small bracts at the base, involucral bracts 2, 2½ by ½ mm opposite, oblong, persistent or caudocous when anthesis. Floral tube c. 1½ cm, Floral tube gradually enlarged towards the top, glabrous outside. In side, calyx lobes oblong or ovale 4 by 2 mm glabercent on the both surfaces, papillate, appendages petaloid, stamens, styles exerted up to 6 mm, ovary. Fruit obovate slightly compressed and constricted from the base to apex, usually 2 fruits opposite each peduncle, yellow when ripe, 3-4½ by 5-6½ cm, two locus usually with 2 seeds. Seeds
SILVICULTURE OF GAHARU PLANT
Although gaharu has been known since ancient times, intensified research on a broader aspects of gaharu has just begun in the last ten years. One of these aspects is silviculture of gaharu. Research conducted by R & D Centre for Forest Conservation and Rehabilitation, FORDA of the Ministry of Forestry found that propagation of gaharu plants is not difficult. Gaharu can be propagated from seeds and cuttings. However, cultivation of gaharu plantation in a large scale has a high risk of pest and disease attack. This chapter provides silvicultural practice for gaharu plant.
3.1 Propagation using seeds, cuttings and tissue
culture
Seeds of gaharu are collected while they are still hanging on their mother trees because gaharu fruits crack and throw their seeds before they fall to the ground. Seedlings, on the other hand can be collected from underneath their mother trees. To date, propagation from seeds is still much cheaper than cuttings because seeds stocks are still abundant in the field and can be harvested every year from mother trees in the field. Gaharu seeds, however, is recalcitrant and its germination is affected by storage periods and conditions. A study by Subiakto et al. (2009) revealed that germination percentage of non-stored seeds (direct seeding) was 82% and down to 42% after being stored for 8 weeks at room temperature. Germination was further declining to 24% if the seeds were stored in refrigerator at 4°C for 8 weeks.
Study on vegetative propagation by using cuttings was done by adopting by KOFFCO (Komatsu-Forda Fog Cooling) System (Figure 5). Using this system, Subiakto et al. (2009) found that the best growth media and watering interval for gaharu shoots cuttings were mixtures of coconut dust and paddy husk of 1 : 1 ratio, and twice a week of watering, respectively. These treatments showed the highest rooting percentage, i.e. 69%.
3.2 Inoculation of beneficial microbial to promoted
plant growth
In 2002, Tamuli and Boruah observed the association of arbuscular mycrorrhizal fungi (AMF) with gaharu tree in Jorhat district of the Brahmaputra Valley. They isolated different AMF but found that Glomus is the dominant genus. Among the Glomus spp.,
Glomus fasciculatum is the most dominant followed by Glomus aggregatum. Further studies revealed that AMF and plant growth promoting bacteria (PGPR) promoted early growth of gaharu seedlings. Inoculation of AMF, Glomus clarum and Gigaspora decipiens, increased growth of 180 days old A. filaria, under greenhouse conditions (Figure 5). Growth acceleration was indicated by percentage of AM colonization up to 93%, plant growth (height, diameter and biomass dry weight), survival rate, and nitrogen (70-153%) and phosphorus (135-360%) concentrations (Turjaman et al., 2006). Two PGPR, Burkholderia sp. CK28 (IAA-producing bacteria), and Chromobacterium sp. CK8 (mycorrhization helper bacteria) accelerated height growth of Aquilaria sp. in in the nursery. Percentage of height increase over non-inoculated control seedlings ranges from 12.2 to 38.7%, five months after inoculation. Effect of inoculation was no longer observed after seedlings were transplanted to the fields, probably due to interaction with soil microflora (Sitepu et al., 2009).
3.3 Pest and Disease
Gaharu plants cultivated in a large scale is prone to pest and disease attack. Some important pests and diseases have been found attacking several locations of gaharu plantations in Indonesia with various level of attack (Table 2; Figure 6).
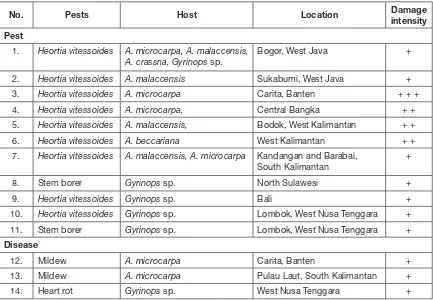
Table 2.
Pests and Diseases of gaharu plants in several locations in IndonesiaNo. Pests Host Location Damage intensity Pest
1. Heortia vitessoides A. microcarpa, A. malaccensis, A. crassna, Gyrinops sp.
Bogor, West Java +
2. Heortia vitessoides A. malaccensis Sukabumi, West Java + 3. Heortia vitessoides A. microcarpa Carita, Banten + + + 4. Heortia vitessoides A. microcarpa, Central Bangka + + 5. Heortia vitessoides A. malaccensis, Bodok, West Kalimantan + + 6. Heortia vitessoides A. beccariana West Kalimantan + + 7. Heortia vitessoides A. malaccensis, A. microcarpa Kandangan and Barabai,
South Kalimantan
+
8. Stem borer Gyrinops sp. North Sulawesi + 9. Heortia vitessoides Gyrinops sp. Bali + 10. Heortia vitessoides Gyrinops sp. Lombok, West Nusa Tenggara + 11. Stem borer Gyrinops sp. Lombok, West Nusa Tenggara +
Disease
12. Mildew A. microcarpa Carita, Banten + 13. Mildew A. microcarpa Pulau Laut, South Kalimantan + 14. Heart rot Gyrinops sp. West Nusa Tenggara +
The most important pest found to date is identified as Heortia vitessoides Moore (Odontiinae, Crambidae) (Irianto et al., 2010). The pest (previously Tyspana vitessoides),
has caused severe damage to many gaharu plantations in Indonesia in the past three years, i.e. in Forest Area with Specific Purposes (Kawasan Hutan dengan Tujuan Khusus, KHDTK) Carita, Banten Province; Bodok, Sanggau, West Kalimantan Province; Kandangan, Barabai, South Kalimantan Province; Malino, East Kalimantan Province; Bali Province; and Lombok, West Nusa Tenggara Province; and South Sumatra (Erdy Santoso, Pers. Comm.). In 2008, the percentage of pest attack in gaharu plantation in KHDTK, Carita, reached 100%, many of the trees defoliated, and about 20 trees died because of recurring attack on newly emerging leaves. This pest has also been reported to have distributed in Fiji, Hongkong, Thailand, and North Queensland (Austtralia) (Herbison-Evams and Crossley, WWW page).
The caterpillars of this species are pale green with a broad knobbly black line along each side. Their head is brown. The caterpillars live in a group in a shelter made by joining a number of leaves together with silk. The caterpillars drop on silk threads if disturbed. When mature, the caterpillars descend and pupate in the soil. The adults have a striking pattern on the forewings of black on pale yellow. The hindwings are white with a broad black margin. The moths have a yellow and black banded abdomen. The wingspan is about 3 cms. The eggs are yellowish-green, and are flattened. They are laid in an overlapping cluster, like tiles on a roof (Herbison-Evams and Crossley, WWW page).
A study by Irianto et al. (2010) investigated pest management strategy to control the population of H. vitessoides. This study found that application of a mixture of systemic and contact insecticides with addition of leaf fertilizer and plant sticker effectively controlled high population of the pest. A biological control approach was also studied by using ants, predator of the pest, that build a nest on gaharu trees and keep the plant protected from the pest. This experiment is still underway but seems encouraging.
Figure 6.
Pests and disease of gaharu trees. (1): Leaf eater Heortia vitessoides; (2): Stem Borer; (3): Colony of ants, the predator of H. vitessoides: (4): Hearth-rot fungi.3.4
Soil characteristics
Table 3.
Soil physical characteristics of three gaharu plantation sites in JavaCarita, Banten Dramaga, Bogor Kampung Tugu, Sukabumi Value Category Value Category Value Category Texture ( %)
0-30 Sand 8.33 8.33 12.78
Silt 25.1 Clay 12.59 Clay 18.73 Clay
Clay 66.57 79.08 68.49
30-60 Sand 8.55 6.33 9.95
Silt 22.1 Clay 11.98 Clay 5.9 Clay
Clay 69.35 81.69 84.15
> 60 Sand 6.01 5.13 11.54
Silt 36.51 Clay 9.09 Clay 26.37 Clay
Clay 57.48 85.78 62.09
(Source: Pratiwi et al., 2009)
Table 4.
Soil chemical characteristics of three gaharu plantation sites in Java Island, IndonesiaChemical cha-racteristics
Dramaga, Bogor Carita, Banten Kampung Tugu, Sukabumi
Chemical cha-racteristics
Dramaga, Bogor Carita, Banten Kampung Tugu, Sukabumi
Horizon 1 (0-30 cm)
Horizon 2 (30-60 cm)
Horizon 3 (> 60 cm)
Horizon 1 (0-30 cm)
Horizon 2 (30-60 cm)
Horizon 3 (> 60 cm)
Horizon 1 (0-30 cm)
Horizon 2 (30-60 cm)
Horizon 3 (> 60 cm)
KCl (me/100 gr)
Al 3.72 (VL) 4.16 (VL) 4.90 (VL) 5.84 (L) 7.36 (L) 6.40 (L) 2.76 (VL) 2.76 (VL) 6.40 (L)
H 0.33 0.36 0.41 0.49 0.53 0.45 0.3 0.3 0.42
0,05 N HCl (ppm)
Fe 2.04 1.8 1.48 1.72 1 1.04 0.52 0.36 0.32
Cu 3.44 2.64 2.4 1.64 1.68 1.52 1.2 1.12 1.44
Zn 5.24 4.88 5.28 3 2.6 2.8 1.4 1.56 1.56
Mn 85.6 88.01 79.2 28.48 17.08 16.4 17 22.12 26.36
GAHARU BIOINDUCTION TECHNOLOGY
There are many beliefs on how gaharu is formed and these views have strongly been wrapped in myth and history. Only in the last few decades that more scientific approaches have been conducted. Scientists with assistance from the locals have tried to understand the mechanism involved in the formation of gaharu and conduct research based on their understanding. Experiments involve laboratory works and setting-up demonstration plots have been established in some countries, including China, Thailand, Indonesia, and Cambodia. Approach in these studies is generally done by wounding trees deliberately with different treatments to induce gaharu formation.
Some extensive researches and their findings are detailed in this chapter.
4.1 Deliberate tree wounding using mechanical tools
Wounding gaharu tree using blade or hammering of nails into the trunks has been used widely in the past, however gaharu yielded from this treatment is generally of inferior quality and cannot meet the desired market demand (Persoon, 2007). It will take many years before high quality gaharu is formed. However, gaharu hunters in Papua New Guinea, for example Imnai village people (Yapsiei) that deliberately wounded gaharu trees in an attempt to stimulate gaharu production reported that they were able to harvest gaharu of B and C grades, three years after this treatment (Gunn et al., 2003). They believe that following this wounding, muddy water enters the tree through the wound is responsible for gaharu production. Pojanagaroon and Kaewrak investigated various mechanical methods in stimulating gaharu formation including making a holes with screws, wounding using chisels, and bark removal with hatchets. The study found that methods of injury had influence in gaharu formation and was determined by seasonal changes, rainy season accelerated gaharu formation faster than dry season. However, the gaharu produced by this method was reported to only yielded pale fragrant and small percentage of essential oil.
4.2 Deliberate tree drilling and chemical injection
Another country, Thailand being a large producers and consumers of gaharu traditionally, conducted similar experiment and establish gaharu plantation. The development of gaharu in this country has been intensive because of the rapid decrease in gaharu supply from the nature. Krissana Panasin company in Chantaburi, Southeast Thailand has established gaharu plantation of several hundred hectares. Similar project has been established in Merauke, Papua, Indonesia by a Catholic Church and in Papua New Guinea, which involves trials of numerous methods to treat the trees.
4.3 Deliberate tree drilling and inoculation of fungal
inoculum
The formation of gaharu is a result of plant defense mechanism towards fungal attacks by producing resinous compounds as secondary metabolite. In their natural habitat, the process of resin accumulation as the result of tree-pathogen battle may take many years and the longer the process takes place the more expensive and highly valuable the resin is. Many scientists have been passionately trying to understand the cascade process of this tree-fungi interaction in producing gaharu. Isolation of various fungi from infected trees have been widely reported (Table 5).
The finding of novel technology to accelerate gaharu formation based on this mechanism continues to sprout and research has become more intensive. In this section, some experiments conducted in Indonesia for gaharu induction are presented. An emphasis is given for induction technology developed by Forestry Research and Development Agency and a thorough information is presented at the later part of this section.
Table 5.
Fungi isolated from infected tree identified based on theirmorphological characteristics and experiments related with the fungal inoculation
No. Fungi Host isolated Activity Author
1. Torula sp. A. agallocha Formation of gaharu Bose (1934 in Burfield, 2005a)
Stopped in 1931 due to inoculum contamination Sagopal (1959 in Burfield, 2005a)
2. Cladosporium sp. Obtained result but, trees were destroyed. Burfield (2005a).
Work stopped and no later work yielded positive results
3. Epicoccum granulatum
Further investigation of gaharu formation by fungus Battcaharrya (1952 in Burfield, 2005a)
Prospecting the deliberate tree infection by fungus Sadopal (1960 in Burfield, 2005a)
Showed conflicting results Varma (1977 in Burfield, 2005a)
4. Pencillium citrinum A. agallocha Ascribed spiral cavitation of the tracheid walls of the wood of A. agallocha to P. parasitica
Hawksworth et al. (1976 in
Burfield, 2005a)
Aspergillus tamari Phialophora parasitica was frequently associated with better quality portions of gaharu
Gibson (1977 in Burfield, 2005a)
Aspergillus spp. Gaharu formation rarely occurred in trees under 25 years old, and formation followed injury to the tree, for example following wind or storm damage
Rahman (1980 in Burfield, 2005a)
Fusarium solani
No. Fungi Host isolated Activity Author
A. agallocha Succeeded in inoculating healthy wooden blocks so that colony growth occurred
Tamuli et al. (2000 in Burfield, 2005a)
Chaetum globosum
Kunze
Colonies were deposited in cultures with the MTCC, Institute of Microbial Technology (CSIR), Chandigarh 8. Fusarium xylaroides Aquilaria spp. from
Java, Kalimantan, Mollucas, Sumatra
Fusarium spp. Have successfully infected gaharu-producing trees in the field and yielded gaharu products
Agustini et al. (2006)
Fusarium tricinctum Colonies were deposited in cultures with the FORDA CC, Laboratory of Forest and Nature Conservation R&D Center, Ministry of Forestry, Indonesia
Isolation of fungi from tropical rain forest in Rayong, Chanthaburi, Trad, Nakhon Ratchasima, Krabi, Trang and Pattarung, Thailand.
Subansenee et al. (1985)
Curvularia lunata Four species: B. theobromae, C. lunata, F. oxysporum,
and Pestalotia sp. were parasitic and others were saprophytic.
A study by a group of researcher in Laboratory of Biotechnology, Agriculture Faculty, Mataram University, Indonesia conducted inoculation of fungi for stimulation of gaharu production. The group found that F. lateritum could be isolated and cultured in potato dextrose agar (PDA) media and that the fungi could form gaharu after 2-month injection to 4-year-old trees. Biology Tropical (BIOTROP) reported the use of stressing agent on a tree prior to fungal injection accelerated the formation of gaharu.
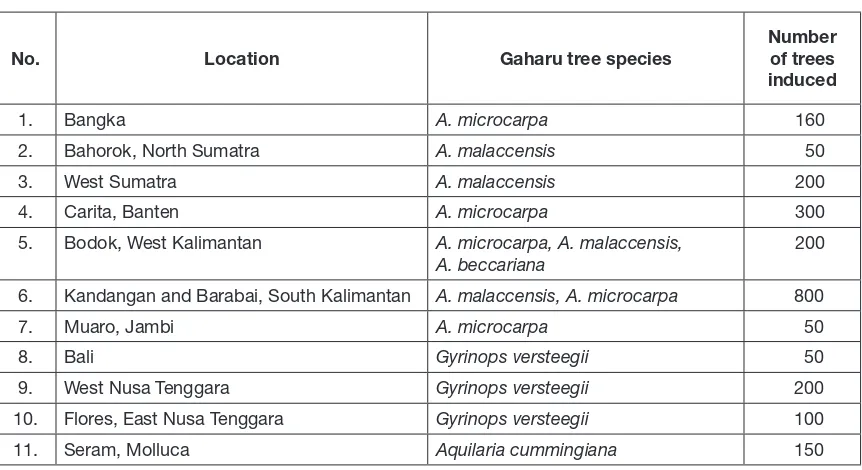
Research and development of artificial induction technology of gaharu in Forestry Research and Development Agency, Ministry of Forestry has started since 1984, but only in the last ten years that the activities have become more intensive (Table 6). The first trial for artificial induction was located in Sebadu Village, West Kalimantan, Indonesia by injecting five fungal-producing gaharu: Botryodiplodia sp., Fusarium oxysporum, F. bulbigenium, F. laseritium, and Phytium sp. on a total of 300 trees of 8-year-old A. microcarpa. Before injection, palm sugar and lubricant were added and mixed with the fungi. This preliminary study showed that the formation of gaharu was improved by the addition of palm sugar and lubricant. The three Fusarium speciesformed larger area of gaharu on the infected trees (Santoso, pers. comm.). The institution also collaborated with Provincial Forestry Institute in Kalimantan by injecting Fusarium sp. into 9-year-old
A. malaccensis grown in Sempaja arboretum. After three months, gaharu formation was indicated by stem cracking and the skin was easily torn. The infected part of stem is fragrant when it was burnt confirming that gaharu was formed.
Table 6.
Experimental plot of gaharu trees induction by deliberate tree drilling and Fusarium spp. injection in several locations across IndonesiaNo. Location Gaharu tree species
Number of trees induced
1. Bangka A. microcarpa 160
2. Bahorok, North Sumatra A. malaccensis 50 3. West Sumatra A. malaccensis 200 4. Carita, Banten A. microcarpa 300 5. Bodok, West Kalimantan A. microcarpa, A. malaccensis,
A. beccariana
200
6. Kandangan and Barabai, South Kalimantan A. malaccensis, A. microcarpa 800
7. Muaro, Jambi A. microcarpa 50
8. Bali Gyrinops versteegii 50
9. West Nusa Tenggara Gyrinops versteegii 200 10. Flores, East Nusa Tenggara Gyrinops versteegii 100 11. Seram, Molluca Aquilaria cummingiana 150
(Source: Santoso et al., unpublished data)
Novel findingsby Santoso et al. have revealed important aspects that determine the successful of gaharu formation by artificial induction, i.e. methods of injection, fungal strain type, and growing media for delivering the fungi. Intensive studies for several years have confirmed efficient gaharu inducing methods (Figure 6, 7, and 8), as follows:
1. Injection hole is of small size of about 3 mm in diameter. The holes will be closed naturally by the plant, not long after inoculants injection. This closing process of the injection hole is important in stimulating the formation of gaharu
3. Type of fungal strain determines the gaharu formed, so screening of efficient strain using few samples in several locations to confirm its efficiency is essential prior to establishing large demonstration plots
4. Spaces in between holes should be wide enough (about 25 cm apart) to prevent from overlapping of vertical disease development from each other’s hole
5. The quality of gaharu formed becomes higher with longer incubation time. the Gaharu product harvested after three years of induction using this method was classified as tanggung a grade higher than kemedangan, while gaharu harvested from shorter incubation period was considered as kemedangan grade A-B.
u
Figure 7.
An illustration of induction procedure for stimulation of gaharuFigure 9.
Harvesting procedure of gaharu product from deliberate tree injection with fungal inoculant. (1): Initial symptom of gaharu formation on stem;(2): Felling of tree; (3-7): Cutting away tissue to obtain the resinous parts; (8): Residue of trees that is used for distillation of oil.In search for gaharu-inducing fungi, thirty six fungi from infected gaharu trees from 17 provinces in Indonesia have been isolated and subsequently identified by means of 28S rRNA partial gene sequencing (Sitepu et al., in preparation for publication). Identification was conducted in the Laboratory of Forest Microbiology, FORDA using FUS1 and FUS2 primers that enabled amplification of up to 460-bp fragment. Most isolates identified were members of Fusarium solani species complex, only one isolate, FORDACC-02375, originated from East Kalimantan showed high similarity to Fusarium oxysporum (Table 5). This study highlighted a rapid molecular identification protocol for gaharu-inducing fungi over the conventional measure.
Table 7.
Molecular identification of 36 strains of gaharu-inducing fungi collected from 17 provinces in IndonesiaNo. Isolate Number Origin (Province) Molecular identification
1 FORDACC506 North Sumatra Fusarium solani
2 FORDACC509 Gorontalo Fusarium solani
7 FORDACC510 Molucca Fusarium solani
8 FORDACC497 Central Kalimantan Fusarium solani
9 FORDACC499 West Kalimantan Fusarium solani
10 FORDACC2372 East Nusa Tenggara Fusarium solani
11 FORDACC504 Riau Fusarium solani
12 FORDACC514 Papua Fusarium solani
13 FORDACC502 West Sumatra Fusarium ambrosium
14 FORDACC515 East Nusa Tenggara Fusarium sp. 15 FORDACC2379 Molucca Fusarium solani
16 FORDACC511 West Nusa Tenggara Fusarium solani
17 FORDACC2370 Bangka Belitung Fusarium solani
18 FORDACC517 Bangka Belitung Fusarium solani
19 FORDACC513 Papua Fusarium solani
20 FORDACC519 West Java Fusarium falciforme
21 FORDACC2375 East Kalimantan Fusarium oxysporum
22 FORDACC520 West Java Fusarium solani f. batatas 23 FORDACC518 Babel Fusarium solani f. batatas 24 FORDACC2371 Babel Fusarium solani
25 FORDACC2377 West Java Fusarium solani
26 FORDACC507 Lampung Fusarium solani f. batatas 27 FORDACC498 Central Kalimantan Fusarium solani
28 FORDACC2369 West Sumatra Fusarium ambrosium
29 FORDACC495 South Kalimantan Fusarium solani
30 FORDACC2373 West Nusa Tenggara Fusarium solani f. batatas 31 FORDACC2374 East Kalimantan Fusarium solani
32 FORDACC508 Bengkulu Fusarium sp. 33 FORDACC505 North Sumatra Fusarium solani
34 FORDACC496 South Kalimantan Fusarium solani f. batatas 35 FORDACC516 Babel Fusarium solani
36 FORDACC2378 West Java Fusarium solani
CHEMICAL PROPERTIES OF GAHARU
Gaharu is a resinous wood and it is the resin that determines the quality of gaharu. Many studies have revealed two major constituents of gaharu, i.e. sesquiterpenes and chromones, as the main source of the fragrant.
About 50 years ago, the chemical content of fragrant gaharu was isolated by Indian chemists from Aquilaria agallocha ROXB and they characterized several sesquiterpenes (Konishi et al., 2002). Twenty years later, Japanese scientists isolated and characterized many sesquiterpenes from two gaharu types, the first, presumably originated from A. malaccensis and the second, “kanankoh” (in Japanese) (Konishi et al., 2002). Another constituent of gaharu was also revealed, i.e. an oxygenated chromone derivative, followed by isolations of two new chromone derivatives in 1989 and 1990 by Chinese scientists.
Sapwood gaharu exemplifies as merely unexuded resin, but rather it is deposited in the wood tissues of trees. This resin deposit renders the wood with loose fibers and white color becoming solidly compact, white in color, and fragrant in smell. This resin belongs to sesquiterpene group, which is easily volatile (Ishihara et al., 1991). Most of the compounds in gaharu are identified as sesquiterpenoid group. One of the fragrant-smelling compounds in gaharu was first identified by Bhattacharyya dan Jain as agarol,
categorized as mono-hydroxy compunds (Prema and Bhattacharyya, 1962).
Research conducted by Nakanishi succeeded in characterizing jinkohol (2β-hydroxy -(+)-prezizane) in gaharu originated from Indonesia, through benzene extraction. This team also found two new sesquiterpene compounds in Aquilaria malaccensis Lamk. from Indonesia, comprising jincoheremol dan jincohol II, called as type B to differentiate it from the type A of A. agallocha Roxb., and isolated alpha-agarofuran and (-)-10- epi-gamma-eudesmol, oxo-agarospirol as the main constituent at gaharu type B (Burfield 2005b). In Burfield (2005b), it was stated that Yoneda managed to identify the main sesquiterpene that existed in gaharu type A (in A. agallocha) and type B (in A. malaccensis). Gaharu type A contained β-agarofuran 0,6%, nor-ketoagarofuran 0,6%, agarospirol 4,7%,
jinkoh-eremol 4,0%, kusunol 2,9%, dihydrokaranone 2,4%, and oxo-agarospirol 5,8%. Meanwhile, in gaharu type B were identified compounds comprising α-agarofuran (-)-10-epi-γ-eudesmol 6,2%, agarospirol 7,2%, jinkohol 5,2%, jinko-eremol 3,7%, kusunol
3,4%, jinkohol II 5,6% dan oxo-agarospirol 3,1%.
Figure 10.
Chemical structures of 6 2-(2-phenylethyl) chromones and an unkown 2-(2-phenylethyl)-chromone (1), flidersiachromone. Compound (1): 2-(2-phenylethyl)-chromone, flidersiachromone; (2-7): 2-(2-phenylethyl)chromones, (2): C19H18O5, (3): C1H14O4, (4): 6-hydroxy-2-[2-(4-hydroxyphenyl)ethyl]chromone, (5): dihydroxyl derivative of 2-(2-phenylethyl)-chromone, (6): C1H14O3, and (7): C18H16O4 (Source: Konishi et al., 2002).Yagura et al. (2005) isolated novel chromone derivatives from gaharu, produced by intentionally wounding A. crassna and A. sinensis. The three compounds were characterized as diepoxy tetrahydrochromones have not been reported from natural gaharu product, i.e. Oxidoagarochromone with molecular formula of A (1) (C17H14O4); B(2): (C18H16O5); C(3): C18H16O6 .
Figure 10.
Chromatogram revealing constituents in 6-month old induced gaharu
Figure 11.
Chromatogram revealing constituents in 6-month old induced gaharuFurther, results of GCMS analysis on the induced gaharu products originated from Dramaga and Carita each comprising 2 sample trees revealed that there were 16 phenol compounds that belong to high group, and 8 phenols as low group (Table 6). Scrutinizing that Table 6, it seems that there has occurred a sequence (series) of secondary metabolite process, such as the evolving/release of iseugenol and veratrol compounds that function as perfumes and medicine, whereby those two compounds are not encountered in regular wood. The veratrol itself is evolved from phenol compounds that undergo hydrolysis into catechol, which further through a sequence of complex mechanisms, i.e. Kreb cycle, is transformed to veratrol. Likewise, eugenol compounds are evolved from guaiacol (main constituent of lignin) through ferulic acid intermediate (Waluyo et al., 2011).
Results of identification on gaharu resin indicated the presence of caryophene compounds that typify the main constituents for eugenol which usually exists in clove leaves. In gaharu resin were also identified cembren compounds (diterpenoid) that comprised a feromon compound effective for termites, a palustrol compound as antitusive, and copaene compounds that can function as essential oil and are rather toxic to be taken orally if the LD is 5000 mg/kg
Recent study revealed distinct chemical compositions and its relative concentration of four gaharu woods from A. microcarpa that was artificially induced by four Fusarium
spp. from four different localities in Indonesia (Novriyanti et al., in press). Analysis using GCMS pyrolysis revealed that Fusarium sp. from Tamiang Layang, Central Kalimantan had the highest confirmed constituent (12.89%), but gaharu induced by Fusarium sp. from Molluca had the highest total concentration of odorant compounds (12.47%).
Further study done by the same group of scientists at FORDA of the Ministry of Forestry, Indonesia compared the chemical contents of gaharu formed by natural process and by artificial induction (Santoso et al., unpublished data). The GCMS analysis showed differences in chemical contents of gaharu harvested from these two processes (Figure 11).
sesquiterpenes and chromone derivates have been isolated from agarwood and some of these have sedative analgesic effects. Phytochemical research has been carried out on the trunk and resin of agarwood, but little is known about the pharmacological effects of agarwood leaves (Kakino et al., 2010).
Table 8.
Phenol compounds present in the induced gaharu productsNo. Compound name
4 1,4-benzenediol, 2-methoxy- (CAS) hydroquinone, 2-methoxy 12 phenol, 2-methoxy-4-(1-propenyl)- (E) (CAS)
(E)-isoeugenol
0.95 0.85 1.12 1.38
13 phenol, 2-methoxy-4-(2-propenyl)- (CAS) eugenol 0.19 14 Phenol, 4-(3-hydroxy-1-propenyl)-2-methoxy-(CAS)
19 benzaldehyde, 4-hydroxy-3-methoxy- (CAS) vanilin 0.70 20 benzeneacetic acid, 4-hydroxy-3-methoxy-(CAS)
homovanillic acid
0.56
21 Capcaisin 0.09
22 Jasmolin II (CAS) cyclopropane carboxylic acid,
0.22
23 octanoic acid (CAS) caprylic acid 0.16 24 1,3-benzenediol, 4-ethyl- (CAS) 4-ethylresorcinol 1.40
Total 40.45 39.89 19.78 22.57
CHEMICAL PROPER
TIES OF GAHAR
d G
ah
aru
:
(3) (4)
Gaharu oil for use in aromatherapy has attracted more and more attention nowadays, especially for psychosomatic disease caused by stress. In the United Stated, aromatherapy is allowed for clinical use in syndromes, such as Attention Deficit Disorder and Attention Deficit Hyperactivity Disorder (Takemoto et al., 2008). During the aromatherapy, volatile compound is inhaled and it is important to know its pharmacological activity. The composition of volatile compounds in gaharu oil varies. Takemoto et al.
(2008) examined two types of gaharu oil, i.e. from a Hong Kong market and originated in Vietnam, by SPME-GCMS to characterize their volatile compounds. SPME-GC analysis revealed the composition in the gas phase, as follows: sample originated from Hong Kong market contained 47.1% benzylacetone, and sample of Vietnam made contained 61.5% a-gurjunene and 24.7% (+)-calarene as the main volatile components. GC analyses revealed volatile mass in a liquid extract, as follows: benzylacetone was accounted for only 0.96%, a-gurjunene for 15.1% and (+)-calarene for 17.3% of the whole oil (Figure 12). Spontaneous vapor administration system applied using 400µl of gaharu oil showed that these two oil types gave similar sedative activity although the main component of each oil was different (Takemoto et al., 2008).
Figure 13.
Structures of compound from gaharu oilWhen used as sedative, gaharu may affect central nervous system. A sesquiterpene characterized as spirovetivane-type sesquiterpene, (4R,5R,7R)-1(10)-spirovetiven -11-ol-2-one at a concentration of 100µg/ml, caused an induction effect on brain-derived neurotrophic factor mRNA expression in vitro neuronal cells of rat (Ueda et al., 2006).
GAHARU PRODUCTS AND TRADING
Today, the range of gaharu products seems endless from raw materials to various by-products. Varieties of gaharu products continue to blossom in particular, in countries where gaharu has been appreciated for many years traditionally and historically. In Japan, Korea and Taiwan, countries with long tradition of gaharu, for instance, solid heartwood of gaharu is crafted to form beautiful sculptures, appreciated as ‘natural art’. In these countries also, gaharu is used make beads and bracelets (Persoon, 2007). The unscented wood of gaharu has been used in Assam for papermaking, while the wood fiber is used for ropemaking. In Taiwan, gaharu is used as wine ingredient in Chu-yeh Ching and Vo ka Py. Japan and Arab, however demand high quality gaharu to be further processed. Most of the resinous woods is processed and formed into chips, oil and powder (Figure 14 and 15).
Gaharu incense and perfumes are used as a skin daub for embalming, and to give fragrant to soap and shampoo. Oil is used in the production of perfume as fixative and other cosmetics. The oil is also used in the production of traditional Chinese and Korean medicine, medicinal wine and various other products. In Arab, the essential oil of gaharu is the most expensive oil which can be ten times more expensive than sandalwood oil. Soap and hand body lotion from gaharu oil have also emerged for beautifying skins. The oil is extracted from the gaharu through distillation. This delicate process determines both the amount and quality of oil produced. With the exception of large solid pieces of gaharu which are traded as individual pieces, most of the wood is ground into very small pieces or powder, which is immersed in water and left to ferment over time. Then the material is transferred to distillation kettles and steamed. After heating, the condensed water and oil are captured in a container where the oil floats on top of the water. The water is removed and the oil is tapped. The price of high quality oil can be as much as US$50,000 to US$80,000 per litre. This process can be repeated once or twice depending on the quality of the water and the costs of the distillation process. The powder which remains after distillation can be used for low grade incense making. It is estimated that for the production of one litre of oil 100 to 150 kilos of gaharu is necessary.
A range of capacity in oil production per tree was reported in Cropwatch (2005). An 80 year old tree could yield 6-9 kg of gaharu oil, although another report in India suggested yield of 2-7 – 3.6 kg oil/tree. A much lower yield of 1 kg oil/tree was also reported.
Most gaharu traders estimate the quality of gaharu organoleptically. Grading quality based on botanical description and its origin is not in place. A practical analysis of its chemical content is also highly difficult for practical implementation at the trading sites. CITES also regulates all parts and derivatives of Aquilaria spp., Gyrinops spp. and