Vol. 4 No. 1. Hal 26-31 ISSN: 2087-7706
POLA DISTRIBUSI FOTOSINTAT PADA FASE VEGETATIF BEBERAPA
VARIETAS KEDELAI PADA TANAH MASAM DI SULAWESI TENGGARA
Distribution Pattern of Photosynthate of Vegatif Fase of Some Soybean
Varieties on Acid Soil of Southeast Sulawesi
SARAWA*), ARSY AYSYAH ANAS, DAN ASRIDAJurusan Agroteknologi Fakultas Pertanian Universitas Halu Oleo, Kendari
ABSTRACT
This study was to determine differences in the distribution of photosynthate of vegetative phase of several varieties of soybean (Glycine max L. Merril) cultivated on acid soil of Southeast Sulawesi. This research was conducted in the sub district Anduonohu, Poasia district and at Agrotechnology Laboratory, Faculty of Agriculture University Halu Oleo, Kendari from July to October 2013. The research design used was randomized block design (RBD) with three replications. Variety treatment consisted of varieties Kaba, Argomulyo, Tanggamus, Grobogan and Gema. The variables observed in this study were root dry weight, stem dry weight, leaf dry weight, stem dry weight, and branch dry weight. Results of statistical analysis of the data showed that the treatment affected the distribution of photosynthate on different plant varieties. Different varieties gave significant effect on root dry weight, stem dry weight, leaf dry weight, branch dry weight.
Keyw or ds : Acid soils, distr ibution of photosynthate, soybean, var ieties
1
PENDAHULUAN
Kedelai (Glycine max L. Mer r il) mer upakan tanaman yang ber fungsi sebagai sumber pr otein nabati dan dapat digunakan sebagai bahan baku agr oindustr i seper ti tempe, tahu, tauco, kecap, susu kedelai dan untuk keper luan industr i pakan ter nak. Kedelai juga memiliki kandungan gizi yang tinggi yang ber per an untuk membentuk sel-sel tubuh dan menjaga kondisi sel-sel ter sebut. Kedelai Indonesia ter golong memiliki kandungan pr otein yang lebih tinggi dar ipada kedelai impor . Kandungan pr otein kedelai dalam neger i 39-40% sedangkan kedelai impor hanya 35–37%. Selain pr otein, biji kedelai juga mengandung lemak 15,5–24,7% dan kar bohidr at 31,7-31,85% (Banaszkiew icz, 2011). Pr otein biji kedelai juga mengandung ber bagai asam amino yang sangat penting bagi kesehatan, seper ti Isoleucine, Leucine, Lycine, Methionine, Cysteine, Phenylalanine, Tyr osine, Thr eonine, Tr yptophan, Valine, Ar ginine,
*)Alamat kor espondensi:
Email : sar aw a60@yahoo.com
Histidine, Alanine, Aspar tic acid, Glutamic acid, Glycine, Pr oline dan Ser ine (Palomoet al.,
2011). Dengan demikian, biji kedelai mempunyai nilai gizi dan nilai kesehatan yang tinggi. Setiap var ietas member ikan ber bagai var iasi genetik yang ber beda baik dar i sifat maupun kemampuan ber pr oduksi. Di Thailand diver sifitas genetik digunakan sebagai per timbangan dalam per baikan genetik tanaman kedelai (Tantasaw at et al., 2011).
Stabilitas hasil dar i suatu tanaman pada setiap daer ah dapat ter jadi akibat per bedaan kondisi lingkungan baik biotik maupun abiotik. Faktor abiotik meliputi jenis tanah, cur ah hujan, suhu dan kelembaban. Faktor abiotik ser ingkali lebih ber pengar uh ter hadap stabilitas hasil dar ipada faktor biotik kar ena faktor abiotik biasanya ter jadi dalam kur ang w aktu yang lebih lama (Her u, 2014). Besar nya akumulasi bahan ker ing pada fase per iode aw al pengisian biji (fase gener atif) mer upakan kar akter istik yang menentukan per bedaan hasil antar genotipe. Pr oses pr oduksi bahan ker ing ber var iasi ter gantung pada genotipe, kondisi lingkungan dan teknik budidaya yang dilakukan. Par a pemulia ser ing melakukan uji
pada ber bagai kondisi lingkungan ter hadap var ietas yang dpr omosikan untuk dilepas (Gur mu et al., 2009). Per baikan dan pengelolaan genetik telah lama dilakukan pada tanaman kedelai dan ter nyata dapat meningkatkan hasil 31 kg ha-1 tahun-1 di Amer ika (Salvagioti et al., 2008, Specht et al.,
1999). Sejak per baikan genetik maka telah dihasilkan var ietas tanaman kedelai yang memiliki kemampuan menyer ap har a yang tinggi dan tahan pada kondisi lingkungan sub optimal (Br ow n, 1979). Di China telah dilepas 600 kultivar , akan tetapi pr oduktivitas r endah masih menjadi pr oblem ser ius (Liu and Her ber t, 2002; Liuet al.,2008).
Kar akter istik tanah di Sulaw esi Tenggar a yang umumnya digunakan untuk budidaya kedelai ter golong tanah-tanah mar ginal dengan tingkat kemasaman tanah yang sangat tinggi. Tanah masam adalah tanah yang dicir ikan dengan nilai pH r ata-r ata kur ang dar i 5,5; kandungan bahan or ganik tanah (BOT) yang r endah; keter sediaan P dan kapasitas tukar kation (KTK) tanah r endah; tingginya kandungan unsur Mn2+ dan aluminium r eaktif (Al3+) yang dapat mer acuni akar tanaman sehingga menghambat pembentukan bintil akar tanaman legum (Hair iahet al.,2005). Ker acunan Al dan Mn, di samping defisiensi kalsium dan dan fosfor pada tanah masam menyebabkan ter hambatnya per tumbuhan r hizobium dan menur unnya kemampuan infeksi r hizobium pada kedelai (Bakker et al., 1999). Kemasaman tanah telah lama dikenal sebagai salah satu faktor penyebab ter jadinya defisiensi N pada tanaman legum kar ena ber kaitan dengan ter hambatnya pr oses fiksasi N2(Wor knehat al.,2013).
Ber dasar kan hal ter sebut di atas, maka penelitian tentang “Pola Distr ibusi Fotosintat Beber apa Var ietas Kedelai (Glycine max L. Mer r il) pada Tanah Masam Sulaw esi Tenggar a” penting untuk dilakukan untuk mengetahui pola distr ibusi fotosintat beber apa var ietas kedelai.
BAHAN DAN METODE
Waktu dan Lokasi. Penelitian ini
dilaksanakan di Kelur ahan Anduonohu Kecamatan Poasia dan Labor ator ium Agr oteknologi Fakultas Per tanian Univer sitas
Halu Oleo Kendar i. Penelitian ini ber langsung dar i bulan Juli sampai bulan Oktober 2013.
Rancangan Penelitian. Penelitian
dilaksanakan dengan menggunakan Rancangan Acak Kelompok (RAK) yang ter dir i dar i 5 per lakuan var ietas, yaitu var ietas Kaba, Ar gomulyo, Tanggamus, Gr obogan, dan Gema. Masing-masing per lakuan diulang 3 kali sehingga diper oleh 15 unit per cobaan. Dalam setiap unit per cobaan ditempatkan 12 pot sehingga dalam penelitian ini ter dapat 180 pot.
Persiapan Media Tanam. Tanah yang
digunakan adalah tanah masam (podzolik mer ah kuning) dar i kebun per cobaan Fakultas Peter nakan yang ter letak di Kelur ahan Kambu. Tanah yang diambil adalah lapisan atas (top soil) sampai kedalaman ± 30 cm. Tanah diber sihkan dar i sisa akar , r anting, daun tanaman dan kotor an lainnya. Sebelum diisi ke dalam pot tanah diker ing anginkan selama 2-3 har i. Setelah ker ing angin dimasukkan ke dalam pot sebanyak 15 kg. setelah itu tanah disir am hingga mencapai kapasitas lapang.
Penanaman dan Pemeliharaan Benih
kedelai ditanam sebanyak 4 biji per pot. Setelah tanaman tumbuh baik, dilakukan penjar angan sehingga menyisakan 2 tanaman per pot yang selanjutnya menjadi tanaman sampel untuk pengamatan. Pemelihar aan tanaman meliputi penyiangan dan penyir aman. Penyiangan mulai dilakukan pada saat tanaman ber umur 2 minggu setelah tanam dan pada saat gulma sudah mulai tumbuh, sedangkan penyir aman dilakukan setiap har i kecuali ter jadi hujan.
Pengamatan. Var iabel pengamatan pola
distr ibusi fotosintat adalah ber at ker ing akar (g), ber at ker ing batang, ber at ker ing daun, ber at ker ing tangkai daun, dan ber at ker ing cabang. Pengambilan sampel dilakukan secar a acak pada umur 45 HST sebanyak 2 tanaman (1 pot). Mula-mula tanaman didestr uktif secar a pelan-pelan dar i pot agar tidak ada akar yang ter tinggal. Tanaman dipotong-potong dengan menggunakan gunting untuk memisahkan bagian akar , batang, daun, tangkai daun dan cabang. Setiap bagian tanaman dimasukkan ke dalam amplop lalu
diovenkan dengan suhu 80oC selama 2 x 24 jam. Setelah bagian tanaman dinyatakan ker ing selanjutnya ditimbang dengan timbangan analitik.
Analisis Data. Data pengamatan dar i tiap
var iabel dianalisis dengan menggunakan sidik r agam (Anova). Apabila sidik r agam menunjukkan pengar uh nyata (F hitung > F tabel), maka dilanjutkan dengan uji lanjut Beda Nyata Jujur (BNJ) pada tar af keper cayaan 95%.
HASIL DAN PEMBAHASAN
Berat Kering Akar. Ber dasar kan analisis
r agam diper oleh bahwa var ietas kedelai ber pengar uh nyata ter hadap ber at ker ing akar . Rata-r ata ber at ker ing akar ter tinggi diper oleh pada var ietas Ar gomulyo (0,46 g) dan ber beda nyata dengan var ietas Kaba, akan tetapi tidak ber beda nyata dengan var ietas lainnya. Ber at ker ing akar ter endah diper oleh pada var ietas Kaba dan ber beda nyata dengan var ietas Ar gomulyo, akan tetapi tidak ber beda nyata dengan var ietas lainnya (Tabel 1 kolom 2).
Berat Kering Batang. Ber dasar kan hasil
analisis sidik r agam diper oleh bahw a var ietas kedelai ber pengar uh nyata ter hadap ber at ker ing batang. Rata-r ata ber at ker ing batang ter tinggi diper oleh pada var ietas Ar gomulyo (0,80 g), sedangkan r ata-r ata ber at ker ing ter endah diper oleh dar i var ietas Kaba (0,42 g) dan ber beda nyata dengan var ietas Kaba, akan tetapi tidak ber beda nyata dengan var ietas lainnya. Sebaliknya ber at ker ing batang ter endah diper oleh pada var ietas Kaba dan ber beda nyata dengan var ietas lainnya, kecuali var ietas Gr obogan dan var ietas Gema (Tabel 1 kolom 3).
Berat Kering Daun. Ber dasar kan analisis
r agam menunjukkan bahw a var ietas kedelai ber pengar uh nyata ter hadap ber at ker ing daun. Rata-r ata ber at ker ing daun ter tinggi ter dapat pada var ietas Gr obogan (1,28 g) akan tetapi hanya ber beda nyata dengan var ietas Gema. Var ietas Gema member ikan ber at ker ing daun ter endah dan ber beda nyata dengan var ietas Gr obogan dan Ar gomulyo, akan tetapi tidak ber beda nyata dengan var ietas Tanggamus, dan var ietas Kaba (Tabel 1 kolom 4)
Berat Kering Tangkai Daun. Ber dasar kan
analisis r agam menunjukkan bahw a var ietas kedelai ber pengar uh nyata ter hadap ber at ker ing tangkai daun. Rata-r ata ber at ker ing tangkai daun ter tinggi ter dapat pada var ietas Ar gomulyo (0,314 g) dan ber beda nyata dengan var ietas lainnya kecuali dengan var ietas Tanggamus tidak ber beda nyata. Var ietas Gr obogan member ikan ber at ker ing tangkai daun ter endah akan tetapi hanya ber beda nyata var ietas Ar gomulyo (Gambar 1 kolom 5).
Berat Kering Cabang. Ber dasar kan
analisis r agam menunjukkan bahw a var ietas kedelai ber pengar uh nyata ter hadap ber at ker ing cabang. Rata-r ata ber at ker ing cabang ter tinggi ter dapat pada var ietas Gema ( 0,21 g) akan tetapi hanya ber beda nyata dengan var ietas Gr obogan. Sebaliknya ber at ker ing ter endah diper oleh pada var ietas Gr obogan dan hanya ber beda nyata dengan var ietas Gema (Gambar 1 kolom 6).
Distribusi Fotosintat, Par tisi fotosintat
(ber at ker ing) pada akar , batang, daun,tangkai daun, dan cabang telah diper oleh dar i ber bagai var ietas seper ti ter lihat pada tabel 1 ber ikut ini.
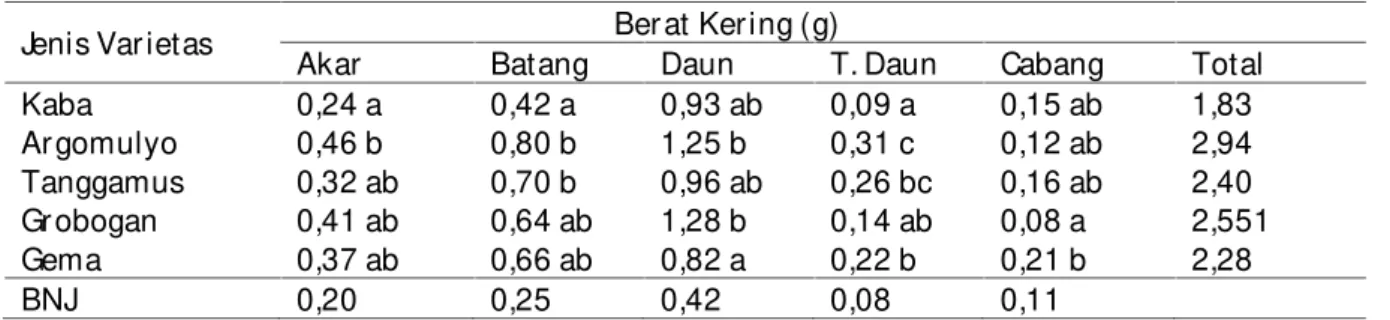
Tabel 1. Ber at ker ing bagian bagian tanaman dar i ber bagai var ietas kedelai
Jenis Var ietas Ber at Ker ing (g)
Akar Batang Daun T. Daun Cabang Total
Kaba 0,24 a 0,42 a 0,93 ab 0,09 a 0,15 ab 1,83 Ar gomulyo 0,46 b 0,80 b 1,25 b 0,31 c 0,12 ab 2,94 Tanggamus 0,32 ab 0,70 b 0,96 ab 0,26 bc 0,16 ab 2,40 Gr obogan 0,41 ab 0,64 ab 1,28 b 0,14 ab 0,08 a 2,551 Gema 0,37 ab 0,66 ab 0,82 a 0,22 b 0,21 b 2,28 BNJ 0,20 0,25 0,42 0,08 0,11
Keter angan. Angka-angka yang diikuti dengan hur uf yang tidak sama pada kolom yang sama ber beda nyata berdasarkan Uji BNJ pada taraf kepercayaan 95%.
Pada tabel ter lihat bahwa semua var ietas member ikan par tisi fotosintat ter tinggi ke daun, disusul ke batang, ke akar , dan ke cabang. Vaietas Ar gomulyo member ikan
par tisi fotosintat ter tinggi ke semua bagian tanaman, kecuali ke cabang par tisinya lebih r endah dibandingkan dengan var ietas Kaba, Gema, dan Tanggamus (Gambar 1)
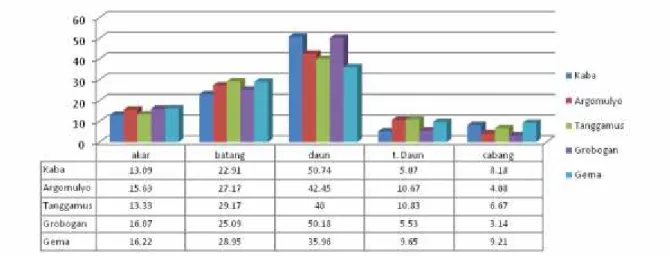
Gambar 1. Per sentase distr ibusi fotosintat beber apa var ietas kedelai pada ber bagai bagian vegetatif tanaman
Var ietas Ar gomulyo member ikan ber at ker ing total ter tinggi (2,94 g) member ikan par tisi fotosintat ke daun 42,45%, batang 27,17%, akar 15,63%, ke tangkai daun 10,67% dan ke cabang 4,08%. Var ietas Gr obogan member ikan ber at ker ing ter tinggi kedua (2,55 g) member ikan par tisi fotosintat ke daun 50,18%, batang 25,09%, akar 16,07%, tangkai daun 5,53% dan ke cabang 3,34%. Var ietas Tanggamus member ikan ber at ker ing total ter tinggi ke 3 (2,4 g) dan member ikan par tisi fotosintat ke daun 40,00%, batang 29,17%, akar 13,33%, tangkai daun 10,83% dan cabang 6,67%. Var ietas Gema member ikan ber at ker ing total ter tinggi ke 4 (2,28 g) dan member ikan par tisi fotosintat kedaun 35,96 %, batang 28,95 %, akar 16,22 %, tangkai daun 9,65 % dan ke cabang 9,21 %. Var ietas yang paling r endah ber at ker ing totalnya adalah var ietas Kaba (1,83 g) dan member ikan distr ibusi fotosintat ke daun 50,74%, batang 22,91%, akar 13,09%, tangkai daun 5,07%, dan ke cabang 8,18%.
Pembahasan
Ber dasar kan hasil analisis var ians ter hadap ber at ker ing akar , ber at ker ing batang, ber at ker ing daun, ber at ker ing tangkai daun, dan ber at ker ing cabang diper oleh bahw a var ietas member ikan pola distr ibusi fotosintat yang ber beda ter hadap var iabel ter sebut. Hal ini
disebabkan kar ena per bedaan sifat dan kar akter masing-masing var ietas menyebabkan pola par tisi fotosintatnya juga ber beda. Pfeiffer dan Har r is (1990) melapor kan bahw a gen sangat efektif mempengar uhi per tumbuhan, dan ber at ker ing tanaman kedelai ter utama pada R1. Ter dapat hubungan yang signifikan antar a hasil dengan gen yang mempengar uhi per tumbuhan vegetatif tanaman (Pfeiffer , 2000). Ber at ker ing tanaman ser ing dijadikan sebagai alat untuk menyeleksi var ietas kedelai yang mampu member ikan pr oduksi yang tinggi (Scot et al.,2005).
Adanya per bedaan ber at ker ing (akumulasi bahan ker ing ke tanaman) dar i kelima var ietas ter hadap var iabel yang diamati, diduga disebabkan oleh adanya per bedaan sifat atau keunggulan dar i masing-masing var ietas sesuai dengan genotipe yang dimiliki oleh setiap var ietas ter sebut. Menur ut Sitompul dan Gur itno (1995), per bedaan susunan genetik tanaman mer upakan salah satu penyebab ker agaman penampilan tanaman. Pr ogr am genetik akan diekspr esikan pada bagian sifat tanaman yang mencakup bentuk dan fungsi tanaman yang menghasilkan ker agaman per tumbuhan tanaman. Ker agaman penampilan tanaman akibat susunan genetik selalu mungkin ter jadi
sekalipun bahan tanaman yang digunakan ber asal dar i jenis yang sama.
Per bedaan yang ber agam dar i masing-masing var ietas ter hadap var iabel yang diamati dapat disebabkan oleh per bedaan genetik dar i var ietas ter sebut dan adaptasinya ter hadap lingkungan tumbuh tanaman. Dimana, setiap var ietas memiliki daya adaptasi yang ber beda-beda untuk dapat tumbuh secar a maksimal. Menur ut Lovelles (1989), suatu fenotipe (penampilan dan car a ber fungsinya) individu mer upakan hasil inter aksi antar a genotip (w ar isan alami) dan lingkungannya. Walaupun sifat khas suatu fenotipe ter tentu tidak dapat selamanya ditentukan oleh per bedaan genotipe atau oleh lingkungannya, ada kemungkinan per bedaan ter sebut disebabkan oleh per bedaan lingkungan atau keduanya. Selain disebabkan oleh faktor genetik, per bedaan distr ibusi fotosintat dar i setiap var ietas yang diujikan juga disebabkan oleh faktor lingkungan (tanah masam). Kondisi lingkungan (tanah masam) menyebabkan tanaman mengalami cekaman. Pada tanah masam, keter sediaan unsur P menjadi kendala utama dalam meningkatkan hasil tanaman. Per tumbuhan tanaman kedelai pada tanah masam mender ita akibat cekaman abiotik dan biotik, seper ti per tumbuhan vegetatif ter hambat sebagai akibat kekur angan har a makr o dan mikr o, ker acunan Al atau Mn, pembentukan nodul ter hambat, tanaman mudah mendapat cekaman keker ingan dan per tumbuhan akar nya ter hambat (Sumar no, 2005).
Per bedaan akumulasi ber at ker ing dar i beber apa var ietas tanaman kedelai yang diujikan juga dapat dilihat dar i per sentasi distr ibusi fotosintat dar i setiap var iabel yang diamati. Dar i pengamatan per sentase distr ibusi fotosintat setiap bagian tanaman kedelai diketahui bahw a akumulasi fotosintat untuk semua var ietas lebih banyak dialir kan ke bagian daun dibanding ke bagian tanaman lainnya. Hal ini menunjukkan bahw a daun (sour ce) masih aktif dan mempengar uhi besar nya kapasitas sink. Sour ce meliputi or gan-or gan yang mampu mempr oduksi fotosintat yang ber lebih selain untuk or gan ter sebut. Contohnya daun dewasa yang mampu ber fotosintesis. Sink meliputi or gan-or gan non-fotosintetik dan gan-or gan yang tidak mampu mempr oduksi fotosintat yang cukup untuk kebutuhan or gan ter sebut. Contohnya,
daun yang belum mampu ber fotosintesis, akar , batang, biji, bunga, buah dan or gan lainnya. Per sentase ber at ker ing daun untuk setiap var ietas yaitu 39,73% (var ietas Kaba), 31,48% (var ietas Ar gomulyo), 33,98% (var ietas Tanggamus), 42,07% (var ietas Gr obogan) dan 27,10% (var ietas Gema). Besar nya distr ibusi fotosintat ke bagian daun dapat disebabkan oleh masih adanya daun-daun muda yang ter bentuk dan ter us ber tambahnya ukur an daun tanaman pada saat tanaman sudah memasuki fase gener atif. Hal ini menyebabkan pembagian fotosintat banyak didistr ibusikan ke bagian daun ter sebut dan menyebabkan kur angnya akumulasi fotosintat yang ter simpan dalam biji. Menur ut Mustamu (2009), ber at atau besar nya ber at daun disebabkan oleh kegiatan fotosintesis yang tetap diper tahankan tinggi oleh tanaman. Selanjutnya, distr ibusi fotosintat juga banyak didistr ibusikan ke bagian akar , batang dan polong.
Ber dasar kan dar i deskr ipsi var ietas kedelai diketahui bahw a semua var ietas yang diujikan memiliki pola per tumbuhan deter minit yaitu saat tanaman ter sebut memasuki fase gener atif, per tumbuhan vegetatifnya diakhir i. Namun, dar i hasil penelitian diketahui bahwa saat tanaman memasuki fase gener atif, fase per tumbuhan vegetatifnya juga masih ter us ber langsung yang ditunjukkan oleh masih banyaknya daun-daun muda yang ter bentuk dan masih ber tambahnya ukur an sel daun tanaman. Kondisi ter sebut menunjukkan r espon tanaman dalam ber adaptasi dengan lingkungan tumbuhnya.
SIMPULAN
Ber dasar kan hasil penelitian dan pembahasan dapat disimpulkan sebagai ber ikut:
1. Ter dapat per bedaan distr ibusi fotosintat tanaman kedelai antar a var ietas Kaba, Ar gomulyo, Tanggamus, Gr obogan dan Gema yang ditanam pada tanah masam. 2. Var ietas Ar gomulyo dan Gema yang
ditanam pada tanah masam memiliki pola distr ibusi fotosintat cender ung lebih ter baik dibandingkan dengan var ietas lainnya.
3. Semua var ietas member ikan par tisi fotosintat ter tinggi ber tur ut-tur ut ke kedaun, batang, akar , tangkai daun, dan cabang.
UCAPAN TERIMA KASIH
Kami menyampaikan ter ima kasih kepada Dikti yang telah member ikan dana penelitian melalui Skim Fundamental Univer sitas Halu Oleo tahun 2013. Hal yang sama juga kami sampaikan kepada tim peneliti, dan selur uh mahasiswa Agr oteknologi yang ter libat membantu dalam penelitian ini.
DAFTAR PUSTAKA
Bakker MR, Ker isit R, Ver bist K, Nys C. 1999. Effect of liming on r hizospher e chemistr y and gr ow th of fine r oots and of shoot of sessile oak. Plant Soil. 217 : 243-255.
Bambar a S, Ndakidemi PA. 2010. The potential r ole of lime and molybdenum on the gr ow th, nitr ogen fixation and assimilation of metabolites in nodulated legum. A Special r efer ence to Phaseolus vulgar is. Afr . J. Biotechnol 8: 2482 - 2489
Br ow n JC. 1979. Genetic impr ovement and nutr ient upatake. In plant 29:289-292
Gur mo F, Mohammed H, Alemaw G. 2009. Genotype X invir onment inter actions and stability of Soybean for gr ain yield and Nutr ition quality. Afr ican Cr op Sci Jour nal 17:87-99.
Hair iah K, Widianto, Supr ayogo D. 2005. Dapatkah pengembangan budidaya tanaman pangan pada tanah masam selar as dengan konsep per tanian sehat? Pr osiding Lokakar ya Pengembangan Kedelai di Lahan Sub-optimal. Puslitbangtan Bogor .
Her u K. 2014. Potensial yield of soybean lines ar e higher than their par ent Indonesia low land popular var iety. Inter . J. Of Biol. 6.2. :49-57 Liu XB, Har ber t SJ. 2002. Fiefteen year s of r esear ch
axamining cultivation of continnous soybean in Nar theast China. Field Cr ops Res. 79: 1-7 Liu XB, Jin J, Wang GH, Her ber t SJ. 2008. Soybean
yield physiology and develovement of hight -yielding pr actices in Nor theast China. Field Cr op Res 105:157-171.
Lovelless AR. 1989. Pr insip-Pr insip Biologi Tumbuhan Untuk Daer ah Tr opik. Ter jemahan Kar taw inata K, Dinimihar ja S, Soetisna U. Gr amedia, Jakar ta.
Mustamu YA. 2009. Seleksi kedelai gener asi F4 ter hadap intensitas cahaya r endah di dua lingkungan [Tesis] . Pasca Sar jana Institut Per tanian Bogor .
Palomo I, Guzman L, Leiva E, Mujica V, Car r asco G, Mor gado N, Gonzalez DR, 2011. Soybean pr oducts consumption in the pr evention of car diovascular diseases. In H.A. El-Shemy (Ed). Soybean and Health. In Tech: Rijeka, Cr oatia. Pfeiffer TW, Har r is LC. 1990. Soybeans yield
delayed planting as affected by alleles incr easing vegetative w eight. Field Cr op. Res. 23:93-101
Pfeiffer TW. 2000. Selecting soybean for adaptation to double cr opping on the basis of full season plant height. Cr op Sci. 40: 387-390.
Salvagiotti F, Cassman KG, Specht JE, Walter DT,. Weiss A, Daker mann A. 2008. Nitr ogen uptake, fixation and r esponsto fer tilizer N in Soybeans. Field Cr ops Res. 105 : 1-18
Scot TR, David W, Wesley WC. 2005. Nitr ogen application yield and ear ly dr y metter accumulation in late-planted soybean. Cr o. Sci. 15(3) : 854-858.
Sitompul SM, Gur itno B. 1995. Analisis Pertumbuhan Tanaman. Gadjah Mada Univer sity Pr ess, Yogyakar ta.
Specht JE, Hume DJ, Kumudinia SV. 1999. Soybean yield potential-A genetic and physiologycal per spective. Cr op. Sci. 39:1560-1570
Sumar no. 2005. Str ategi Pengembangan Kedelai di Lahan Masam. Pr osiding Lokakar ya Pengembangan Kedelai di Lahan Suboptimal. Puslitbangtan Bogor .
Tantasaw at P, Tr ongchuen J, Pr ajongjai T, Jenw eer aw at S, Chaow iset W. 2011. SSR Analysis of soybean (Glicine max (L) Mer r ) genetic r elationship and var iety identification in Thailand. Aust. J. Of Cr op Sci 5.3 : 283-290. Wor kneh B, Tesfu K, Jafer D. 2013. Gr ow th and
nodulation r esponse of soybean (Glicine max(L) Mer r il) to Lime,Bradyrhizobium japonicumand nitr ogen fer tilizer in acid soil at Melko, South Water n Ethopia. Inter J. Of Sci 8.1 : 25 – 31.