95
Efek Stres Anthropogenik Terhadap Struktur Komunitas Makrozoobentik Substrat
Lunak Perairan Laut Dangkal di Teluk Buyat, Teluk Totok dan Selat Likupang
(Semenanjung Minahasa, Sulawesi Utara)
Lawrence J. L. Lumingas, Ruddy D. Moningkey dan Alex D. KambeyFakultas Perikanan dan Ilmu Kelautan, Universitas Sam Ratulangi, Manado
e-mail: [email protected]
Diterima 28 Maret 2011, disetujui untuk dipublikasikan 2 Mei 2011 Abstrak
Ekosistem perairan laut dangkal di Semenanjung Minahasa diduga telah mengalami tekanan akibat berbagai kegiatan pembangunan dari daratan seperti pertambangan, pertanian dan limbah domestik. Penelitian ini bertujuan untuk menganalisis respon makrozoobentos terhadap berbagai tingkat stres anthropogenik seperti 'tailing', tambang tradisional dan kegiatan pertanian terhadap struktur komunitas makrozoobentos substrat lunak di semenanjung Minahasa. Pengambilan sampel (sampling) telah dilaksanakan pada bulan April dan Mei 2010 pada 10 stasiun sampling di zona subtidal di teluk Buyat, teluk Totok, dan selat Likupang. Dalam penelitian ini, telah dikaji variabel komunitas seperti kelimpahan dan keanekaragaman spesies termasuk indeks keanekaragaman spesies Shannon-Wiener (H’), indeks kekayaan spesies (SR), indeks kemerataan spesies (J’), indeks dominasi Berger-Parker (d), kurva ‘K-dominance’ serta ‘assemblage’ (grup) makrozoobentik dengan analisis multivariat baik analisis klasifikasi maupun analisis faktorial korespondens. Dalam penelitian ini telah diperoleh total 543 individu yang termasuk dalam 114 spesies (takson). Struktur komunitas dan ‘assemblage’ makrozoobentik substrat lunak di semenanjung Minahasa ditentukan oleh kondisi substrat dan tingkat stres anthropogenik khususnya sedimentasi, baik berasal dari ‘tailing’ maupun dari kegiatan pertambangan rakyat. Struktur komunitas serta ‘assemblage’ makrozoobentik di perairan dangkal selat Likupang umumnya berada pada kondisi alamiah dengan dominasi spesies sensitif, sedangkan yang berada di teluk Totok khususnya dekat muara Sungai Ratatotok berada pada kondisi buruk (awal suksesi) dengan dominasi spesies oportunis. Berdasarkan karakteristik struktur komunitas serta ‘assemblage’ makrozoobentiknya, ekosistem bentos di teluk Buyat belum mencapai tahap akhir suksesi (ekosistem ekuilibrium) kecuali pada kedalaman yang lebih dangkal dekat muara sungai Buyat.
Kata kunci: Komunitas makrozoobentik, Ekologi bentik, Teluk Buyat, Analisis multivariat. Abstract
The ecosystem of shallow marine waters in Minahasa penninsular has got pressures from various development activities in the land, such as mining, agriculture and or domestic wastes. This study was aimed at analyzing the macrozoobenthos response to various levels of anthropogenic stresses, such as tailing, traditional mining and agricultural activities on the community structure of soft substrate macrozoobenthos, in Minahasa penninsular. Sampling was carried out in April and May, 2010, at 10 sampling sites in the subtidal zone of Buyat bay, Totok bay and Likupang strait. In this study, the community variables, such as species abundance and diversity using species diversity index of Shannon-Wiener (H’), richness index (SR), eveness index (J’), dominance index of Berger-Parker (d), ‘K-dominance’ curve, were measured. Macrobenthos assemblage was analysed using multivariate analysis, both for classification analysis and factorial correspondence analysis. The study recorded a total of 543 individuals of 114 species (taxa). The soft substrate macrozoobenthic community structure and assemblage in Minahasa penninsular were determined by the substrate condition and the level of anthropogenic stresses, especially sedimentation from either tailing or traditional mining activities. Both macrozoobenthos community structure and assemblage in shallow waters of Likupang strait were in natural condition dominated by sensitive species, but those in Totok bay, near Ratatotok downstream, were in bad condition (early succession) dominated by opportunistic species. Based on the characteristics of macrozoobenthic community structure and assemblage, it was found that the benthic ecosystem of Buyat bay has not reached the final phase of succession (ecosystem equilibrium), except the shallow waters near Buyat river mouth.
Keywords: Macrozoobenthic communities, Benthic ecology, Buyat bay, Multivariate analysis. 1. Pendahuluan
Makrozoobentos adalah fauna yang menghuni bagian dasar perairan yang berukuran diameter tubuh lebih besar dari 1 mm atau yang tertahan pada ayakan dengan ukuran lubang 1 mm (Collignon, 1991).
Fauna ini umumnya hidup melata, menetap, menempel, memendam dan meliang di dasar perairan baik substrat lunak maupun substrat keras. Komunitas makrozoobentik laut umumnya terdiri atas empat kelompok utama yakni Mollusca, Annelida (Polychaeta), Crustacea dan Echinodermata
dan kelompok lain yang terdiri atas berbagai filum kecil lainnya seperti Sipunculida, Cnidaria dan Nemertea (Lumingas, 1990).
Komponen makrofauna dalam komunitas bentik sering dikuantifikasi untuk menentukan kesehatan lingkungan karena organisme tersebut relatif bersitat sedenter, mempunyai masa hidup yang panjang dan spesies makrobentos menunjukkan perbedaan toleransi terhadap stress (Dauer, 1984). Gray dkk. (1988) mengemukakan dibanding dengan meiozoobentos dan mikrozoobentos, makrozoobentos lebih banyak dan lebih umum digunakan untuk memantau dan sebagai indikator pencemaran. Hal ini disebabkan karena informasi taksonomik dan biologi makrozoobentos lebih banyak diketahui.
Keterbatasan mobilitas untuk menghindari kondisi yang kurang menguntungkan mengakibatkan bentos sering terekspos pada kontaminan yang terakumulasi dalam sedimen dan konsentrasi oksigen yang rendah dalam perairan bentik, sehingga komunitas bentik dapat menggambarkan kondisi lingkungan lokal (Smith dkk, 2001). Dengan demikian perubahan kondisi lingkungan perairan dapat tergambar atau terekam lewat perubahan struktur komunitas makrozoobentiknya atau berfungsi sebagai 'pita rekaman' perubahan lingkungan di sekitarnya.
Berbagai kajian telah menunjukkan bahwa bentos dapat menanggapi secara teramal berbagai tingkat maupun jenis stres baik alami maupun karena manusia (anthropogenik) (Pearson dan Rosenberg, 1978; Dauer, 1993; Orlando-Bonaca dkk, 2008). Pranovi dkk (2008) bahkan menggunakan data tentang komunitas makrobentik di Laguna Venesia yang tersedia dari tahun 1935 sampai tahun 2004 dan memungkinkan menggambarkan perubahan struktur komunitas selama hampir 70 tahun dengan menunjukkan penurunan yang tajam keanekaragamannya.
Ekosistem perairan laut dangkal (subtidal) di sekitar Semenanjung Minahasa diduga telah mengalami tekanan (stres) akibat berbagai kegiatan pembangunan (dampak anthropogenik) dari daratan seperti pertambangan, industri, pelabuhan, domestik perkotaan serta pertanian. Di teluk Buyat, misalnya, selama kurang lebih 8 tahun (1996-2004) telah dijadikan tempat pembuangan limbah tambang emas (tailing) dengan kapasitas sekitar 2000 ton perhari pada kedalaman hanya sekitar 82 meter (Blackwood dan Edinger, 2007). Tailing yang yang dipompakan lewat pipa dengan debit yang tinggi akan menghasilkan gundukan dan awan bottom dengan kandungan ‘total suspended solid’ (TSS) berkisar 200 000 s/d 600 000 mg/l (Coumans, 2002). Efek fisik pembuangan tailing tersebut diduga telah memberikan dampak yang serius bagi komunitas bentik di perairan tersebut. Penghentian pembuangan tailing ke dalam teluk Buyat sejak September 2004 memungkinkan rekolonisasi komunitas bentik menuju kondisi ekilibrium. Dalam rangka manajemen lingkungan laut yang efektif diperlukan
kajian struktur komunitas makrozoobentik laut dangkal dalam merespon gradien stres anthropogenik tersebut di atas.
Penelitian ini bertujuan untuk menganalisis dampak berbagai tingkat stres anthropogenik, seperti 'tailing', tambang tradisional dan kegiatan pertanian pada struktur komunitas makrozoobentik terutama komposisi dan kelimpahan spesies pada substrat lunak di semenanjung Minahasa. Hasil penelitian ini akan memberikan informasi status lingkungan pada masing-masing lokasi berdasarkan komposisi spesies, struktur komunitas serta ‘assemblage’ makro-zoobentik, dan informasi mengenai rona awal komunitas bentik untuk program monitoring dalam manajemen lingkungan laut maupun pesisir.
2. Bahan dan Metode
Pengambilan sampel (sampling) dalam penelitian ini dilaksanakan pada bulan April dan Mei 2010 pada 10 stasiun sampling di zona subtidal masing-masing 4 stasiun di teluk Buyat, 3 stasiun di teluk Totok, dan 3 stasiun di selat Likupang (Gambar 1). Posisi geografis serta karakteristik stasiun sampling dapat dilihat pada Tabel 1. Pada setiap stasiun diambil 1 satuan sampel sedimen dengan menggunakan grab La Motte berpenampang 30 cm x 20 cm. Sampel yang diambil dimasukkan dalam kantong plastik dan diawetkan dengan formalin 10 %, selanjutnya dibawa ke Laboratorium Hidrobioekologi dan Manajemen Sumberdaya Perairan Fakultas Perikanan dan Ilmu Kelautan Universitas Sam Ratulangi Manado.
Di laboratorium, sampel sedimen dicuci dengan air keran untuk menghilangkan formalin dan garam, disaring dengan ayakan berukuran lubang (mata jaring) 1 mm, diberi pewarna ‘rose bengal’ kemudian dilakukan penyortiran bentos dengan bantuan mikroskop stereo. Bentos yang tersortir dipindahkan ke dalam cawan petri dan diawetkan dengan alkohol 70 % untuk selanjutnya diidentifikasi dengan menggunakan mikroskop stereo (disecting microscope) berdasarkan buku petunjuk yang tersedia seperti antara lain Day (1967), Abbott (1977), Campbell dan Nicholls (1979), Guille dkk (1986), Abbott dan Dance (1990), Kozloff (1990) dan Dance (1993) serta menggunakan sarana internet.
Untuk menganalisis ‘assemblage’ makrozoobentos (pengertian Gray dan Elliott, 2009), data yang dikumpulkan pertama-tama ditabulasi ke dalam tabel kontingensi dua arah (tabel silang) yang terdiri atas beberapa baris (spesies) dan beberapa lajur (stasiun). Data yang telah dimasukkan ke dalam tabel silang tersebut ditransformasikan ke dalam bentuk log (x+1) untuk mengstabilkan varians dan untuk membuat nilai nol berarti (Thouzeau, 1989, Bakus, 2007). Kemudian data tersebut dianalisis dengan menggunakan metode analisis data peubah ganda (multivariate data analysis): dengan pilihan menu Analisis 'Cluster' (Bakus, 2007) dengan metode hubungan (lingkage method) ‘average’ dan pengukuran jarak (distance measure) ‘Euclidean’
untuk analisis normal (stasiun); dan Analisis Faktorial Korespondensi (AFK) untuk memberikan peragaan geometrik di mana variabel yang diteliti dipetakan menjadi titik-titik dalam salib sumbu. AFK ini cocok untuk menganalisis variabel dan hasil observasi yang telah disajikan dalam bentuk tabel kontingensi atau matriks (Lebart dkk., 1982). Aplikasi AFK dalam penelitian ini bertujuan memberikan peragaan terbaik secara simultan antara kelompok observasi (lajur) dan kelompok variabel (baris), untuk mendapatkan korespondensi atau hubungan yang benar antara dua variabel yang diteliti (spesies dan stasiun). Pengolahan data untuk Analisis Klasifikasi dan Analisis Faktorial Koresponden digunakan paket komputer MINITAB 15.
Pada masing-masing stasiun akan dihitung juga: Indeks Keanekaragaman Shannon-Wiener (H') (Ludwig dan Reynolds, 1988):
s i i i n n n n H 1 ln ,di mana ni adalah jumlah individu spesies i dan n adalah jumlah total individu dalam sampel.
Indeks Kekayaan Spesies (SR) (Ludwig dan Reynolds, 1988): SR = s-1/ln n, di mana s adalah jumlah spesies. Indeks Kemerataan Spesies (J') (Ludwig dan Reynolds, 1988): J' = H'/ln s. Indeks Dominansi Berger-Parker (d) (Gray dan Elliott, 2009): d = nmax/n, di mana nmax adalah jumlah
individu dari spesies yang paling berlimpah.
Selain itu, dipetakan kurva persentase kelimpahan kumulatif (ordinat) dengan rangking spesies (absis) yang dinamakan kurva ‘K-dominance’ (Lambshead dkk, 1983 dalam Bakus, 2007) untuk menggambarkan tingkat dominansi spesies. Kurva K-dominans ini juga digunakan sebagai pelengkap untuk menilai struktur komunitas maupun untuk menduga tingkat suksesi komunitas makrozoobentik pada stasiun-stasiun pengamatan (Warwick dan Ruswahyuni, 1987).
Gambar 1. Peta lokasi penelitian.
Tabel 1. Posisi geografis serta karakteristik stasiun sampling Stasiun (kode) Posisi geografis Kedalaman
(meter)
Tipe sedimen Kondisi Hidrodinamis Sumber Stres Buyat 1 (BUY1) Buyat 2 (BUY2) Buyat 3 (BUY3) Buyat 4 (BUY4) Totok 1 (TOT1) Totok 2 (TOT2) Totok 3 (TOT3) Likupang 1 (LIK1) Likupang 2 (LIK2) Likupang 3 (LIK3) 0° 50’ 27,6” LU 124° 42’ 22,6” BT 0° 50’ 33,4” LU 124° 42’ 12” BT 0° 50’ 40” LU 124° 42’ 10,1” BT 0° 50’ 33,7” LU 124° 42’ 0,6” BT 0° 51’ 23,8” LU 124° 42’ 34” BT 0° 53’ 22,6” LU 124° 44’ 57,5” BT 0° 53’ 24,2” LU 124° 44’ 58,3” BT 0° 41’ 2,1” LU 125° 4’ 20,8” BT 0° 42’ 48,3” LU 125° 2’ 26,9” BT 0° 42’ 42,1” LU 125° 1’ 25,4” BT 60 55 40 15 23 22 28 20 20 45 lumpur lumpur lumpur lumpur + serasah lumpur + serasah pasir kasar pasir kasar pasir sedang pasir sedang pasir halus terbuka/tinggi terbuka/tinggi terbuka/tinggi terbuka/tinggi terlindung/rendah terlindung/rendah terlindung/rendah terbuka/tinggi terbuka/tinggi terlindung/sedang tailing tailing tailing pertanian/muara sungai tambang rakyat/ muara sungai pertanian/ domestik pertanian/ domestik pertanian pertanian pertanian
3. Hasil dan Pembahasan 3.1 Hasil
Tabel 2 berisi komposisi spesies makrozoobentos dan kelimpahannya pada setiap stasiun sampling di tiga lokasi penelitian (teluk
Buyat, teluk Totok dan selat Likupang). Dari 10 stasiun (sampel) di Semenanjung Minahasa ditemukan 543 individu yang termasuk dalam 114 spesies (beberapa takson seperti Nemertea, Hemichordata, Copepoda dan lainnya yang tidak teridentifikasi dianggap masing-masing sebagai satu spesies). Sebanyak 90,8% dari jumlah individu
tersebut tersusun dari Polychaeta (289 individu atau 53,2%), Crustacea (143 individu atau 26,3%) dan Mollusca (61 individu atau 11,2%). Dari jumlah spesies tersebut, 89,5% merupakan gabungan dari ketiga grup taksonomis masing-masing Polychaeta (52 spesies atau 45,6%), Crustacea (28 spesies atau 24,6%) dan Mollusca (22 spesies atau 19,3%). Kecuali di stasiun Buyat 2, Polychaeta mendominasi makrozoobentik yang ditemukan, dengan proporsi tertinggi pada stasiun Totok 1 (77,8%). Pada stasiun Buyat 2, Crustacea mendominasi proporsi makro-zoobentos dengan 33,3% sedangkan Polychaeta hanya 23,8%. Polychaeta, Crustacea dan Mollusca menyusun lebih dari 90 % makrozoobentos pada stasiun-stasiun Likupang 2, Likupang 3, Totok 1, Totok 2, Buyat 1 dan Buyat 4. Pada stasiun Totok 1 dan Totok 2 tidak ditemukan Mollusca sedangkan di stasiun Totok 3 tidak ditemukan Crustacea.
Pada Tabel 3 terlihat bahwa stasiun Likupang 2 memiliki kelimpahan individu tertinggi dengan 140 individu atau dengan kepadatan mencapai 2333 individu/m2, sedangkan yang terendah kelimpah-annya adalah stasiun Totok 3 dengan 9 individu/m2 atau dengan kepadatan 150 individu/m2.
Jumlah spesies (takson) tertinggi terdapat pada stasiun Likupang 2 dan stasiun Buyat 4 dengan masing-masing memiliki 39 spesies dan jumlah spesies terendah terdapat pada stasiun Totok 3 dengan hanya memiliki 8 spesies. Nampaknya stasiun-stasiun Likupang memiliki kelimpahan baik individu maupun spesies makrozoobentos lebih tinggi dibanding stasiun-stasiun lainnya (43-140 individu/grab dan 23-39 spesies) diikuti stasiun-stasiun Buyat (21-113 individu/grab dan 14-39 spesies) dan yang terendah adalah stasiun-stasiun Totok (9-21 individu/grab dan 8-15 spesies).
Syllis sp adalah spesies yang paling berlimpah khususnya di stasiun Likupang 2 yang mencapai 31 individu dalam satu grab. Takson Nemertea memiliki tingkat kehadiran paling tinggi yakni terdapat hampir pada semua stasiun kecuali stasiun Buyat 1. Tiga spesies dari grup Polychaeta errant yang memiliki kehadiran yang tinggi (terdapat pada tujuh stasiun) adalah Glycera sp, Nephtys sp dan Paralacydonia paradoxa.
Pada Tabel 3 terlihat juga bahwa indeks keanekaragaman Shannon-Wiener (H’) berkisar dari 1,71 (stasiun Totok 1) sampai 3,29 (stasiun Buyat 4). Nilai indeks kekayaan spesies (SR) terendah terdapat di stasiun Totok 1 dengan nilai 2,77 dan tertinggi terdapat di stasiun Buyat 4 dengan nilai 8,04. Untuk kemerataan spesies, nilai indeksnya (J’) berkisar dari 0,78 (stasiun Totok 1) sampai 0,98 (stasiun Totok 3). Nampak bahwa nilai-nilai H’, J’ dan SR berkorelasi negative dengan nilai indeks dominasinya (d) yang berkisar dari 0,12 (stasiun Likupang 1 dan stasiun Buyat 4) sampai 0,5 (stasiun Totok 1).
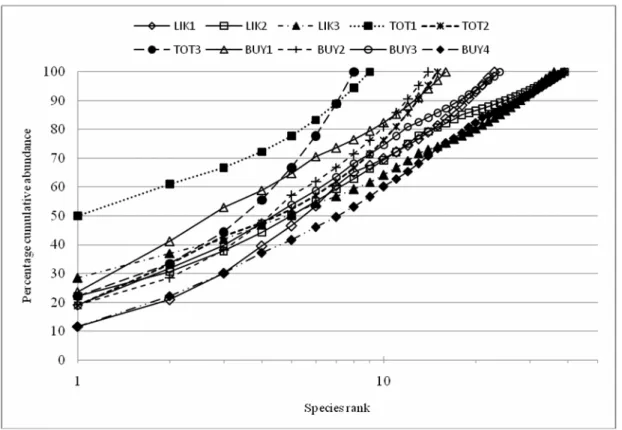
Perbandingan kurva-kurva K-dominans berdasarkan kelimpahan individu untuk kesepuluh stasiun sampling dapat dilihat pada Gambar 2. Pada gambar tersebut nampak bahwa kurva stasiun Totok
1 cepat mencapai puncak yang menunjukkan adanya dominasi satu spesies (Nereis sp) dan hanya tersusun dari sedikit spesies dengan keanekaragaman yang rendah. Kondisi ini hampir sama dengan komunitas makrozoobentik di stasiun Totok 3 yang memiliki keanekaragaman spesies yang rendah tetapi tidak menunjukkan adanya dominasi satu spesies kecuali karena rendahnya jumlah spesies.
Sebaliknya kurva-kurva stasiun Buyat 4, stasiun Likupang 2 dan stasiun Likupang 3 nampak mencapai puncak lebih lambat, yang berarti tingginya tingkat keanekaragaman spesies. Hal ini bukan saja karena banyaknya jumlah spesies penyusun komunitas tetapi juga dengan tingkat kesama-rataan spesies yang tinggi. Untuk kurva stasiun Likupang 1, walaupun jumlah spesies tidak terlalu tinggi tetapi menunjukkan keanekaragaman spesies yang relatif tinggi karena rendahnya dominasi dan tingginya kemerataan spesies. Kurva-kurva yang terletak di tengah-tengah seperti stasiun-stasiun Buyat 1, Buyat 2, Buyat 3 dan Totok 2 menunjukkan keaneka-ragaman spesies dengan tingkat sedang.
Gambar 3 merupakan dendogram yang mengklasifikasikan kesepuluh stasiun sampling ke dalam 4 grup yang didasarkan pada kelimpahan 114 spesies. Keempat grup tersebut adalah Grup A (Likupang 1-Likupang 2), Grup B (Totok 2-Totok 3), Grup C (Totok 1-Buyat 1-Buyat 3-Buyat 2-Buyat 4) dan Grup D (Likupang 3). Nampaknya keempat grup stasiun tersebut berhubungan dengan tipe sedimen (lihat Tabel 1). Stasiun Grup A terdiri atas stasiun-stasiun dengan tipe substrat bersedimen pasir sedang, Grup B terdiri atas stasiun-stasiun berpasir kasar, Grup C terdiri atas stasiun-stasiun bersedimen lumpur dan Grup D yang hanya terdiri atas satu stasiun (Likupang 3) bersedimen pasir halus.
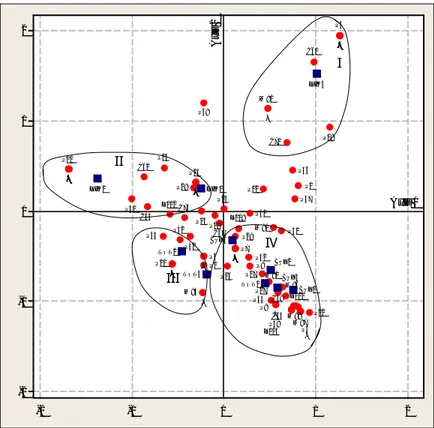
Analisis Faktorial Koresponden (AFK) dilakukan berdasarkan data kelimpahan setelah tranformasi log (x+1) dalam tabel kontigensi dua arah yakni 114 baris spesies dan 10 kolom stasiun. Dalam analisis tersebut, diperoleh total inertia untuk 9 axis (sumbu) adalah 3,6207 dengan kontribusi sumbu 1, sumbu 2 dan sumbu 3 masing-masing 0,6576 (18,16 %), 0,6062 (16,74 %) dan 0,5569 (15,38 %) atau dengan total 50,28 % untuk 3 sumbu pertama. Perbedaan relatif dalam % antara sumbu 2 dan 3 adalah maksimal sehingga yang diinterpretasi hanya 2 sumbu (faktor) pertama (Thouzeau, 1989) (Gambar 4).
Variabel stasiun yang paling bertanggung-jawab terhadap pembentukan sumbu 1 adalah Likupang 2 (58,3% kontribusi absolut), Likupang 3 (23,5%), Buyat 3 (6%) dan Buyat 1 (5,9%) dengan kontribusi varians total sebesar 93,7%. Sedangkan untuk sumbu 2, variabel stasiun yang bertanggung-jawab pembentukannya adalah Likupang 3 (57,4%), Buyat 3 (13,9%), dan Buyat 1 (8,6%) dengan kontribusi absolut total sebesar 79,9%. Stasiun-stasiun yang paling berkontribusi dalam pembentukan sumbu juga sebagai stasiun
karakteristik sumbu tersebut karena memiliki kontribusi relatif yang juga tinggi.
Spesies-spesies dengan kode P11, P21, P41, P44, C61, C63, C67, C69, C70, C72, C75, C76, C77, C78, M83, M89, M96, M97, M100, M101, M102, L104, L113, P3, P6, P10, P14, P16, P23, P34, P35, P45, P46, C53, C79, M82, M85, M90, M93, M98, P51, C60, C65 merupakan spesies-spesies yang bertanggung-jawab atas pembentukan sumbu 1 dengan kontribusi absolute total sebesar 66,6%. Sedangkan spesies-spesies yang bertanggung-jawab atas pembentukan sumbu 2 adalah: P3, P6, P10, P14, P16, P23, P34, P35, P45, P46, C53, C79, M82, M85, M90, M93, M98, C60, M87, M88, C65, P49 dan C59 dengan kontribusi absolute total sebesar 63,5%. Spesies-spesies ini juga memiliki nilai kontribusi relatif yang tinggi sehingga juga merupakan spesies-spesies karakteristik ekslusif sumbu-sumbu tersebut.
Nampaknya sumbu 1 berhubungan dengan faktor granulometri yang memisahkan stasiun-stasiun bersubstrat pasir sedang sampai pasir kasar pada bagian negatif (Likupang 1, Likupang 2, Totok 2 dan Totok 3) dengan stasiun-stasiun bersubstrat lumpur sampai pasir halus pada bagian positif sumbu (Totok 1, Buyat 1, Buyat 2, Buyat 3, Buyat 4 dan Likupang 3). Sedangkan sumbu 2 memisahkan stasiun-stasiun Likupang di bagian positif dan stasiun-stasiun Buyat dan Totok di bagian negatif sumbu.
Secara keseluruhan, dapat dikelompokkan 4 grup stasiun yakni grup I yang hanya terdiri atas stasiun Likupang 3 dengan substrat pasir halus, grup II yang terdiri atas stasiun Likupang 1 dan stasiun Likupang 2 dengan substrat berpasir sedang, grup III yang terdiri atas stasiun Totok 2 dan stasiun Totok 3 dengan substrat berpasir kasar dan grup IV yang terdiri atas stasiun-stasiun Totok 1, stasiun Buyat 1, stasiun Buyat 2, stasiun Buyat 3 dan stasiun Buyat 4 dengan substrat lumpur.Dari Gambar 4 dapat diisolasi 4 grup (assemblage) makrozoobentik dengan karakteristik sebagai berikut:
Grup I adalah makrozoobentos penghuni stasiun Likupang 3 dengan karakteristik bersubstrat pasir halus, terdiri dari 22 spesies: Aonides oxycephala, Armandia sp, Ditrupa arietina, Eurythoe parvecarunculata, Goniada maculate, Lumbrineris aberrans, Megalomma quadrioculatum,
Polyophthalmus pictus, Prionospio cirrifera, Scolaricia sp, Scoloplos sp, Albunea sp, unidentified crab, Copepoda, Paranthura sp, Bellucina civia, Clathrotellina pretium, Hawaiarca uwaensis, Neilonella dubia, Semelangulus miyatensis, Solen brevis, Laevicardium lobulatum;
Grup II adalah makrozoobentos penghuni stasiun-stasiun Likupang 1 dan Likupang 2 dengan karakteristik bersubstrat pasir sedang, terdiri dari 34 spesies: Euchone sp, Eunice sp, Lygdamis sp, Nematonereis unicornis, Onuphis (Nohtria) conchylega, Protodorvillea sp, Sabellides sp, Syllis sp, Ampelisca sp, Cumacea, Galathea sp, Gammaropsis sp, Macrophthalmus sp, Orchestia sp, Ostracoda, Oxyrhyncha, Paguroidea (Hermit crabs), Planopilumnus sp, Sergestes sp, Tanaidacea, Thalamita sp, unidentified Amphipoda, Calliostoma sp, Circa sp, Fragum carinatum, Polyplacophora, Primovula frumentum, Strombus sp, Tellina perna, Trachycardium sp, unidentified Sipuncula, Echinocyamus sp, Holothuroidea, Endomyaria (sea anemon);
Grup III adalah makrozoobentos penghuni stasiun-stasiun Totok 2 dan Totok 3 dengan karakteristik bersubstrat pasir kasar, terdiri dari 13 spesies: Amphiglena sp, Armandia intermedia, Lumbrineris sp, Phyllodoce sp, Pseudeurythoe sp, Pygospio sp, Spiochaetopterus sp, Alima sp, Bathyporeia sp, Gammarus sp, Portunus sp, Pitar sp, Macrophiothrix sp;
Grup IV adalah makrozoobentos penghuni stasiun-stasiun Totok 1, Buyat 1, Buyat 2, Buyat 3 dan Buyat 4, dengan karakteristik bersubstrat lumpur, terdiri dari 31 spesies: Aricidea sp, Chaetozone sp, Dasybranchus sp, Diopatra sp, Heteromastus sp, Magelona sp, Nereis sp, Notomastus aberans, Notomastus sp, Phyllodoce malmgreni, Poecilochaetus serpens, Prionospio sp, Spiophanes sp, Sternaspis scutata, Sthenelais sp, Terebellides stroemi, Branchiopoda, Callianassa sp, Hexapus sexpes, Abrina kinoshitai, Clinocardium sp, Corbula erythrodon, Cycladicama lunaris, Natica stellata, Peronidia sp, Phascolosoma sp, Ophiuroidea, Nemertea, Hemichordata, Dactyloptena orientalis, Pseudoceros sp.
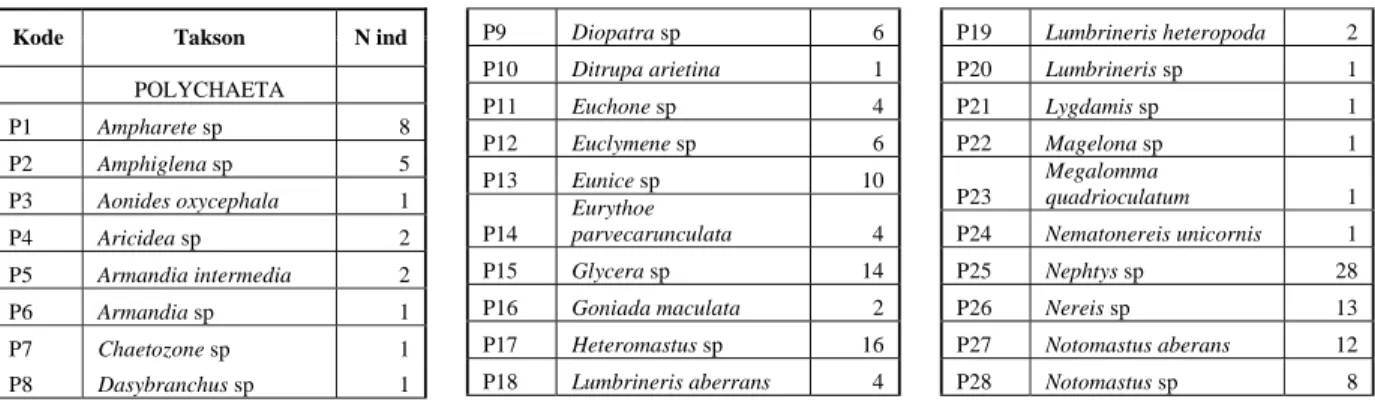
Tabel 2. Komposisi spesies makrozoobentos dan kelimpahannya di semenanjung Minahasa (teluk Buyat, teluk Totok dan selat Likupang)
Kode Takson N ind
POLYCHAETA P1 Ampharete sp 8 P2 Amphiglena sp 5 P3 Aonides oxycephala 1 P4 Aricidea sp 2 P5 Armandia intermedia 2 P6 Armandia sp 1 P7 Chaetozone sp 1 P8 Dasybranchus sp 1 P9 Diopatra sp 6 P10 Ditrupa arietina 1 P11 Euchone sp 4 P12 Euclymene sp 6 P13 Eunice sp 10 P14 Eurythoe parvecarunculata 4 P15 Glycera sp 14 P16 Goniada maculata 2 P17 Heteromastus sp 16 P18 Lumbrineris aberrans 4 P19 Lumbrineris heteropoda 2 P20 Lumbrineris sp 1 P21 Lygdamis sp 1 P22 Magelona sp 1 P23 Megalomma quadrioculatum 1 P24 Nematonereis unicornis 1 P25 Nephtys sp 28 P26 Nereis sp 13 P27 Notomastus aberans 12 P28 Notomastus sp 8
P29 Onuphis (Nohtria) conchylega 17 P30 Paralacydonia paradoxa 13 P31 Phyllodoce malmgreni 2 P32 Phyllodoce sp 2 P33 Poecilochaetus serpens 4 P34 Polyophthalmus pictus 7 P35 Prionospio cirrifera 1 P36 Prionospio ehlersi 4 P37 Prionospio pinnata 3 P38 Prionospio saldanha 2 P39 Prionospio sexoculata 1 P40 Prionospio sp 6 P41 Protodorvillea sp 1 P42 Pseudeurythoe sp 10 P43 Pygospio sp 7 P44 Sabellides sp 1 P45 Scolaricia sp 2 P46 Scoloplos sp 1 P47 Spiochaetopterus sp 2 P48 Spiophanes sp 1 P49 Sternaspis scutata 4 P50 Sthenelais sp 2 P51 Syllis sp 36 P52 Terebellides stroemi 4 CRUSTACEA C53 Albunea sp 1 C54 Alima sp 1 C55 Ampelisca sp 9 C56 Bathyporeia sp 1 C57 Birubius sp 3 C58 Branchiopoda 3 C59 Callianassa sp 20 C60 Copepoda 24 C61 Cumacea 1 C62 Galathea sp 6 C63 Gammaropsis sp 4 C64 Gammarus sp 1 C65 Hexapus sexpes 17 C66 Macrophthalmus sp 1 C67 Orchestia sp 8 C68 Ostracoda 1 C69 Oxyrhyncha 1 C70 Paguroidea (Hermit crabs) 2 C71 Paranthura sp 2 C72 Planopilumnus sp 1 C73 Portunus sp 2 C74 Processa sp 15 C75 Sergestes sp 2 C76 Tanaidacea 10 C77 Thalamita sp 1 C78 Unidentified amphipod 4 C79 Unidentified crab 1 C80 Utica sp 1 MOLLUSCA M81 Abrina kinoshitai 7 M82 Bellucina civia 1 M83 Calliostoma sp 1 M84 Circa sp 1 M85 Clathrotellina pretium 1 M86 Clinocardium sp 1 M87 Corbula erythrodon 12 M88 Cycladicama lunaris 6 M89 Fragum carinatum 1 M90 Hawaiarca uwaensis 1 M91 Laevicardium lobulatum 2 M92 Natica stellata 3 M93 Neilonella dubia 2 M94 Peronidia sp 12 M95 Pitar sp 1 M96 Polyplacophora 1 M97 Primovula frumentum 1 M98 Semelangulus miyatensis 2 M99 Solen brevis 2 M100 Strombus sp 1 M101 Tellina perna 1 M102 Trachycardium sp 1 SIPUNCULA L103 Phascolosoma sp 4 L104 Unidentified Sipuncula 2 ECHINODERMATA L105 Echinocyamus sp 1 L106 Holothuroidea 1 L107 Macrophiothrix sp 2 L108 Ophiuroidea 3 L109 NEMERTEA 15 L110 HEMICHORDATA 1 FISHES L111 Dactyloptena orientalis 3 CEPHALOCHORDAT A L112 Asymmetron sp 8 ANTHOZOA L113 Endomyaria (sea anemon) 9 PLATHELMINTHES L114 Pseudoceros sp 1
Tabel 3. Kelimpahan individu, kepadatan, jumlah spesies (takson), keanekaragaman spesies, kekayaan spesies, kemerataan dan dominasi makrozoobentos di teluk Buyat, teluk Totok dan selat Likupang
Stasiun Variabel Komunitas
LIK1 LIK2 LIK3 TOT1 TOT2 TOT3 BUY1 BUY2 BUY3 BUY4
Jumlah Individu (n) 43 140 81 18 21 9 34 21 63 113 Kepadatan (n/m2) 717 2333 1350 300 350 150 567 350 1050 1883 Jumlah Takson (S) 23 39 36 9 15 8 16 14 24 39 Indeks Shannon (H') 2,93 3,05 3 1,71 2,56 2,04 2,44 2,52 2,82 3,29 Indeks Kekayaan Spesies (SR) 5,85 7,69 7,96 2,77 4,6 3,19 4,25 4,27 5,55 8,04 Indeks Kemerataan Spesies (J') 0,93 0,83 0,84 0,78 0,94 0,98 0,88 0,95 0,89 0,9 Indeks Dominansi Berger-Parker (d) 0,12 0,22 0,28 0,5 0,19 0,22 0,24 0,19 0,19 0,12
Gambar 2. Kurva K-dominance berdasarkan kelimpahan individu untuk kesepuluh stasiun sampling. L I K 3 B U Y 4 B U Y 2 B U Y 3 B U Y 1 T O T 1 T O T 3 T O T 2 L I K 2 L I K 1 1 . 0 1 0 . 6 7 0 . 3 4 0 . 0 0 V a r i a bl e s (S ta ti o n s ) Di s ta n c e A ve r a g e L i n k a g e , C o r r e l a t i o n C o e f f i c i e n t D i s t a n c e
Gambar 3. Dendogram yang menunjukkan klasifikasi dari 10 stasiun sampling makrozoobentos di semenanjung Minahasa. Empat kelompok utama stasiun tersebut telah ditetapkan secara “arbitrary”.
2 1 0 -1 -2 2 1 0 -1 -2 Axis 1 Ax is 2 BUY4 BUY3 BUY2 BUY1 TOT3 TOT2 TOT1 LIK3 LIK2 LIK1 . . L112 L111 . L109 . . .. . L103 ... . . .. M95 . . M92 M91 . . M88 M87 M86 . . . . M81 . . .... C74 . . C71 .. . . . C65 . . C62 . C60 C59 . C57 . C55 . . P52 P51 . P49 . . .. . P43 P42 . P40 . P38 P37 P36 .. P33 P32 . P30 P29 P28 P27 P26 P25 P24 . P22 . P20 P19 P18 P17 . P15 . P13 P12 P11 . P9 P8 P7 . P5 P4 P3 P2 P1 I II III IV
Gambar 4. Proyeksi simultan dari stasiun dan spesies dalam bidang dua dimensi (sumbu 1 dan sumbu 2) dengan menggunakan Analisis Faktorial Koresponden. Didasarkan pada kelimpahan individu makrozoobentos di Semenanjung Minahasa setelah transformasi log (x+1) (Titik-titik yang tumpang tindih: dengan P3 adalah P6, P10, P14, P16, P23, P34, P35, P45, P46, C53, C79, M82, M85, M90, M93, M98; dengan M91 adalah M99; dengan P24 adalah C66, C68, M84, L105, L106; dengan P11 adalah P21, P41, P44, C61, C63, C67, C69, C70, C72, C75, C76, C77, C78, M83, M89, M96, M97, M100, M101, M102, L104, L113; dengan P20 adalah C54, C56, C64; dengan M95 adalah L107; dengan P5 adalah P47, C73; dengan P4 adalah P50; dengan P7 adalah P31, P48, C58, M94, L110, L114, L108; kode lihat Tabel 2).
3.2 Pembahasan
Beberapa peneliti ekologi bentik telah berupaya mengukur perubahan ekologi akibat berbagai stres alamiah dan anthropogenik terhadap ekosistem (Barrett dan Rosenberg, 1981). Oleh karena kesulitan dalam mengukur keseluruhan ekosistem, kebanyakan penelitian hanya terfokus pada tingkat organisasi komunitas. Dalam penelitian ini, telah digunakan dua pendekatan yang saling melengkapi untuk mengkaji struktur komunitas makrozoobentik substrat lunak di Semenanjung Minahasa. Pertama, mengkaji variabel komunitas seperti kelimpahan dan keanekaragaman spesies (termasuk indeks keanekaragaman spesies Shannon-Wiener (H’), indeks kekayaan spesies (SR), indeks kemerataan spesies (J’) dan indeks dominasi Berger-Parker (d)) serta menilai struktur komunitas dengan kurva K-dominance; dan kedua, mengkaji ‘assemblage’ (grup) makrozoobentik dengan analisis multivariat baik analisis klasifikasi (khususnya pengelompokan stasiun yang memiliki karakteristik
spesies yang sama) maupun analisis ordinasi (pengelompokan simultan spesies-stasiun).
Suatu kendala dalam penentuan komposisi spesies makrozoobentos substrat lunak di Semenajung Minahasa adalah terbatasnya pustaka kunci identifikasi makrozoobentos untuk daerah tropis Indo-Pasifik, khususnya Crustacea, Sipuncula dan Nemertea, sehingga beberapa taksa tidak dapat diidentifikasi sampai tingkat spesies. Kendala ini tentu berpengaruh pada penentuan kekayaan maupun keanekaragaman spesies. Oleh karena itu pembahasan hanya terbatas pada pembandingan antar lokasi sampling atau stasiun (variasi spasial).
Selain respon makrozoobentos terhadap stress athropogenik, struktur komunitas juga nampaknya ditentukan oleh respon secara alami baik terhadap kondisi substrat (mikrohabitat) maupun interaksi biotik. Walaupun teluk Buyat pernah dijadikan tempat pembuangan ‘tailing’ tetapi dibanding dengan teluk Totok, kondisi ekologis teluk Buyat masih relatif lebih baik. Pada umumnya komunitas yang mengalami stres anthropogenik mudah kehilangan taksa. Beberapa grup taksonomik yang lebih sensitive
seperti Mollusca dan Crustacea relatif sangat kurang atau bahkan menghilang di teluk Totok.
Demikian juga, rendahnya kelimpahan individu dan keanekaragaman spesies serta tingginya dominasi satu spesies di stasiun Totok 1 menandakan adanya tingkat gangguan maupun stres yang cukup tinggi di stasiun tersebut. Stasiun Totok 1 terletak dekat muara sungai Ratatotok di mana tingkat sedimentasi yang berasal dari daerah pertambangan rakyat sangat tinggi dan masih berlangsung terus. Selain terlindung dari pengaruh hidrodinamika, kondisi substrat di stasiun ini sangat berlumpur dengan warna kehitaman sebagai indikasi rendahnya kelarutan oksigen dan banyak mengandung serasah (detritus dan hancuran tumbuhan) yang berasal dari daratan (Gray dan Elliott, 2009). Stasiun ini didominasi oleh cacing polikhaeta erantia herbivor Nereis sp dan beberapa spesies spionid dan capitellid pemakan deposit yang opportunis seperti Prionospio spp dan Dasybanchus sp serta beberapa penghuni substrat berlumpur seperti Sternaspis scutata dan Callianassa sp.
Dua stasiun lainnya di teluk Totok yakni Totok 2 dan Totok 3, walaupun sama-sama terletak dalam teluk Totok tetapi kedua stasiun ini relatif berbeda kondisi mikrohabitatnya dibanding stasiun Totok 1. Kedua stasiun ini relatif jauh dari muara sungai Ratatotok tetapi dekat dengan pemukiman nelayan yakni desa Basaan 1 dengan substrat miskin bahan organik, berpasir kasar yang berasal dari cangkang foram, moluska dan karang.
Kondisi habitat teluk Buyat sangat spesial karena walaupun sudah 6 tahun masa pemulihan kondisi ekologis tetapi kondisi substrat masih didominasi lumpur yang berasal dari ‘tailing’, terutama pada stasiun-stasiun Buyat 1, Buyat 2 dan Buyat 3. Agak sulit mencari stasiun pembanding atau stasiun referensi di luar teluk Buyat yang memiliki karakteristik habitat yang mirip dengan stasiun-stasiun ini. Biasanya substrat berlumpur hanya ditemukan pada lokasi yang terlindung dari pergerakan massa air (hidrodinamika) dengan kelarutan oksigen yang rendah (hipoksi) atau bahkan anoksi seperti misalnya stasiun Totok 1.
Stasiun di teluk Buyat yang mirip dengan stasiun Totok 1 adalah stasiun Buyat 4, tetapi berbeda dalam hal kondisi hidrodinamika dan tingkat sedimentasi dari sungai. Tidak sebagaimana di stasiun Totok 1 di mana bermuara sungai yang keruh (dari daerah tambang emas tradisional), di stasiun Buyat 4 tingkat sedimentasi dari sungai Buyat relatif rendah. Demikian juga tingkat hidrodinamika yang tinggi di stasiun yang dangkal ini (sekitar 15 m) menyebabkan percampuran massa air yang meningkatkan kelarutan oksigen yang kemungkinan sampai beberapa cm di bawah permukaan sedimen. Kondisi abiotik seperti ini memungkinkan mempercepat struktur komunitas makrozoobentiknya mencapai tahap akhir suksesi (ekuilibrium) setelah mengalami gangguan ekologis baik alami maupun anthropogenik. Stasiun Buyat 4 ini dapat
dikategorikan sebagai stasiun referensi dengan kelimpahan individu, kekayaan dan keanekaragaman spesies yang relatif tinggi.
Dibanding dengan stasiun Buyat 4, stasiun-stasiun Buyat 1, Buyat 2 dan Buyat 3 dapat dikategorikan sebagai stasiun-stasiun dengan struktur komunitas makrozoobentik yang masih dalam tahap suksesi menuju kestabilan (biodiversitas ekuilibrium). Kelimpahan individu, kekayaan spesies dan keanekaragaman spesies makrozoobentos di stasiun-stasiun ini berada pada tingkat menengah. Lambat pulihnya kondisi struktur komunitas makrozoobentik di 3 stasiun Buyat ini diduga disebabkan faktor hidrodinamika, misalnya adanya gangguan tahunan resuspensi sedimen pada musim tertentu. Jika dugaan ini benar maka dapat diramalkan bahwa pulihnya struktur komunitas makrozoobentik mencapai komunitas ekuilibrium akan sejalan dengan perubahan sedimen menuju kondisi alami.
Struktur komunitas makrozoobentik di stasiun-stasiun sampling selat Likupang pada umumnya menunjukkan komunitas stabil dengan kekayaan dan keanekaragaman spesies yang relatif tinggi. Komposisi spesies di stasiun-stasiun Likupang ini agak berbeda dengan yang ada di stasiun-stasiun Buyat. Spesies-spesies makrozoobentos sensitif dari kelompok polychaeta errantia yang karnivora (misalnya Syllis sp, Nephthys sp, Glycera sp) dan Crustacea (misalnya Copepoda, Amphipoda, Processa sp) mendominasi stasiun-stasiun ini. Dibanding dengan stasiun-stasiun Buyat 1, Buyat 2 dan Buyat 3, variable komunitas di stasiun-stasiun Likupang menunjukkan nilai yang lebih baik. Di sekitar perairan Selat Likupang, gangguan dan stress anthropogenik dapat dikategorikan rendah karena jumlah penduduk di pesisir masih relatif sedikit dan kegiatan pertanian umumnya masih tradisional.
Dua stasiun yang menunjukkan dominasi tinggi dan awal suksesi (‘komunitas juvenile’) adalah stasiun Totok 1 dan Totok 3 sedangkan stasiun-stasiun Likupang 2, Likupang 3 dan Buyat 4 menunjukkan komunitas akhir suksesi (‘komunitas klimaks’). Komunitas makrozoobentik di stasiun-stasiun lainnya menunjukkan pola antara (tahap suksesi).
Analisis multivariate baik analisis ‘cluster’ maupun analisis faktorial korespondens telah memisahkan dengan jelas 4 grup (‘assemblage’) makrozoobentik. Oleh karena analisis klasifikasi mengelompokkan stasiun-stasiun berdasarkan kemiripan spesies penghuninya, serta sebaran dan kelimpahan spesies makrozoobentos sangat dipengaruhi oleh kondisi substratnya, maka 4 grup tersebut tersusun dari stasiun-stasiun yang memiliki kondisi substrat (granulometri) yang mirip. Demikian juga dengan analisis ordinasi (AFK), kedekatan antar stasiun dapat berarti adanya kesamaan dalam profil spesies dan sebaliknya, kedekatan titik-titik spesies karena adanya kesamaan profil stasiun. Dengan demikian maka pembentukan 4 grup stasiun adalah
juga merupakan pengelompokan 4 grup spesies yang adalah karakteristik masing-masing stasiun. Spesies-spesies di luar grup adalah Spesies-spesies-Spesies-spesies antara yang lebih toleran terhadap berbagai karakteristik stasiun.
Nampaknya terdapat dua faktor utama pengendali pembentukan 4 grup ekologis ini yakni faktor sedimen khususnya granulometri dan faktor anthropogenik. Sumbu 1 memisahkan kelompok spesies karakteristik habitat bersubstrat pasir sedang (grup II) dan pasir kasar (grup III) di bagian negatif sumbu dengan kelompok spesies karakteristik substrat pasir halus (grup I) dan substrat lumpur (grup IV) pada bagian positif sumbu. Sumbu 2 memisahkan grup (stasiun-stasiun Likupang) dengan dominasi spesies sensitive (ekuilibrium) pada bagian positif sumbu dengan grup (stasiun-stasiun Totok dan Buyat) spesies tolerant (oportunistik) pada bagian positif sumbu. Perubahan struktur komunitas makrozoobentik sering merefleksikan variasi faktor fisika-kimia lingkungan perairan karena fauna bentik memiliki sifat yang terintegrasi dengan kondisi lingkungan sekitarnya (Le Guellec, 1990). Dengan teknik klasifikasi dan ordinasi, Junoy dan Vieitez (1990) telah menyimpulkan bahwa karakteristik sedimen dan tinggi pasang merupakan faktor yang paling bertanggungjawab mengatur sebaran dan kelimpahan komunitas makrozoobentik intertidal. 4. Kesimpulan
Beberapa kesimpulan yang dapat ditarik dari penelitian adalah pertama, struktur komunitas (kelimpahan, sebaran, keanekaragaman serta kekayaan spesies) dan ‘assemblage’ (grup) makrozoobentik substrat lunak di Semenanjung Minahasa merupakan hasil respon terhadap kondisi substrat terutama granulometri dan juga terhadap tingkat stres anthropogenik khususnya sedimentasi, baik berasal dari ‘tailing’ maupun dari kegiatan pertambangan rakyat. Kedua, struktur komunitas serta ‘assemblage’ makrozoobentik di perairan dangkal Selat Likupang umumnya berada pada kondisi alamiah dengan dominasi spesies sensitif, sedangkan yang berada di Teluk Totok khususnya dekat muara Sungai Ratatotok berada pada kondisi buruk (awal suksesi) dengan dominasi spesies toleran (oportunistik). Ketiga, berdasarkan karakteristik struktur komunitas serta ‘assemblage’ makro-zoobentiknya, ekosistem bentos di Teluk Buyat belum mencapai tahap akhir suksesi (ekosistem ekuilibrium) kecuali pada kedalaman yang lebih dangkal dekat muara Sungai Buyat.
Ucapan Terima Kasih
Penelitian ini dibiayai oleh Direktorat Penelitian dan Pengabdian Kepada Masyarakat, Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan Nasional Republik Indonesia sesuai dengan Surat Perjanjian Pelaksanaan Hibah Penelitian No. 009/SP2H/PP/DP2M/III/2010, oleh karenanya Penulis menyampaikan terima kasih.
Daftar Pustaka
Abbott, R. T., 1977, Les Coquillages du Monde, Marabout, Paris.
Abbott, R. T. and S. P. Dance, 1990, Compendium of seashells, American Malacologists, Inc., Melbourne.
Bakus, G. J., 2007, Quantitative Analysis of Marine Biological Communities. Field Biology and Environment, John Wiley and Sons. Inc., Hoboken, New Jersey.
Barrett, G. W. and R. Rosenberg, 1981, Stress Effects on Natural Ecosystems, John Wiley, New York.
Blackwood, G. M. and E. N. Edinger, 2007, Mineralogy and Trace Element Relative Solubility Patterns of Shallow Marine Sediments Affected by Submarine Tailings Disposal and Artisanal Gold Mining, Buyat-Ratatotok District, North Sulawesi, Indonesia, Environ. Geol., 52, 803-818. Campbell, A. C. and J. Nicholls, 1979, Guide de la
faune et de la flora littorales des mers d’Europe, Delachaux and Niestle, Paris. Collignon, J., 1991, Ecologie et biologie marines:
Introduction à l'halieutique. Mason, Paris. Coumans, C., 2002, Submarine Tailings Disposal -
STD Toolkit, Mining Watch Canada, Ottawa and Project Underground, Berkeley, CA, USA.
Dance, S. P., 1993, Les Coquillages, Bordas, Paris. Dauer, D. M., 1984, The use of Polychaete Feeding
Guilds as Biological Variables, Mar. Pollut. Bull., 15:8, 301-305.
Dauer, D. M., 1993, Biological Criteria, Environment Health and Estuarine Macrobenthic Community Structure, Mar. Pollut. Bull., 26, 249-257.
Day, J. N., 1967, A Monograph on the Polychaeta of Southern Africa, Publ. British Mus. (Nat His.), London.
Gray, J. S. and M. Elliott, 2009, Ecology of Marine Sediments. From Science to Management, 2nd ed., Oxford University Press. USA. Gray, J. S., M. Aschan, M. R. Carr, K. R. Clarke, R.
H. Green, T. H. Pearson, R. Rosenberg, and R. M. Warwick, 1988, Analysis of Community Attributes of the Benthic Macrofauna of Frierfjord/Langesundfjord and in a Mesocosm Experiment, Mar. Ecol. Prog. Ser., 46, 151-165.
Guille, A., P. Laboute et J. L. Menou, 1986, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, Orstom, Paris.
Junoy, J. and J. M. Vieites, 1990, Macrozoobenthic Community Structure in the Ria de Foz, an Intertidal Estuary (Galicia, Northwest Spain), Mar. Biol., 107, 329-339.
Kozloff, E. N., 1990, Invertebrates, Sounders College Publ.
Lebart, L., A. Morineau, and J. P. Fénelon, 1982, Traitement des données statistiques, Méthodes et programmes, Dunod, Paris. Le Guellec, C., 1990, Contribution d'une Station
d'épuration pour les Installations Maritimes de Lanvéoc-Poulmic (Finistère) état Initial des sédiments et des Communautes Benthiques, Contrat Travaux Maritimes/ MICROMER/ECOSYSTEM U. B. O., Brest.
Ludwig, J. A. and J. F. Reynolds, 1988, Statistical Ecology, a Primer on Methods and Computing, A Willey Interscience Publ. New York.
Lumingas, L. J. L., 1990, Les Structures Trophiques au sein de la Macrofaune des sédiments, Rapport du DEA, Fac. des Science et Technique, Univ. de Bretagne Occidentale, Brest.
Orlando-Bonaca, M., L. Lipej, and S. Orfanidis, 2008, Benthic Macrophytes as a Tool for Delineating, Monitoring and Assessing Ecological Status: The Case of Slovenian Coastal Waters, Mar. Pollut. Bull. (article in press).
Pearson, T. H. and R. Rosenberg, 1978, Macrobenthic Succession in Relation to Organic Enrichment and Pollution of the Environment, Oceanogr. Mar. Biol. A. Rev., 16, 229-331
Pranovi, F., F. Da Ponte, and P. Torricelli, 2008, Historical Changes in the Structure and Functioning of the Benthic Community in the Lagoon of Venice, Estuar. Coast. and Shelf Sci., 76, 753-764.
Smith, R. W., M. Bergen, S. B. Weisberg, D. Cadien, A. Dalkey, D. Montagne, J. K. Stull, and R. G. Velarde, 2001, Benthic Response Index for Assessing Infaunal Communities on the Southern California Mainland Shelf, Ecol. Applic. 11 :4, 1073-1087.
Thouzeau, G., 1989, Déterminisme du pré-recrutement de Pecten maximus L. en Baie de Saint-Brieuc, Thèse (Tesis S3), Fac. des Science et Technique, Univ. de Bretagne Occidentale, Brest.
Warwick, R. M. and Ruswahyuni, 1987, Comparative Study of the Structure of Some Tropical and Temperate Marine Soft-bottom Macrobenthic Communities., Mar. Biol., 95, 641-649.