RHIZOBACTERIA PENDUKUNG PERTUMBUHAN TANAMAN
Plant Growth Promotor Rhizobacteria
M AK ALAH
Oleh :
I ntan Ratna D ewi A. N I P. 132 30 6 0 8 1
Jur usan Budidaya Per tanian Pr ogr am Studi Agr onom i
FAK U LTAS PERTAN I AN U N I V ERSI TAS PAD JAD JARAN
JATI N AN GOR
KATA PENGANTAR
Puj i syukur dipanj at kan ke hadirat Allah SWT, yang t elah memberikan
berkat dan hidayah-Nya sehingga penulis dapat menyelesaikan Makalah
Plant Gr owt Pr omot ing Rhizobact er i a
Pada kesempat an ini t im penulis menyampaikan ucapan t erima kasih
kepada Prof . Dr Tualar Simarmat a at as saran dan masukan pada penulisan
makalah ini, Kepala Labar ot arium Produksi Tanaman ser t a st af pengaj ar
minat budidaya pada khususnya. Tanpa bant uannya sulit bagi penulis unt uk
dapat menyelesaikan penyusunan makalah ini.
Penulis t elah berusaha unt uk menyempurnakan t ulisan ini, namun
sebagai manusia penulis pun menyadar i akan ket erbat asan maupun
kehilaf an dan kesalahan yang tanpa disadari. Oleh karena it u, sar an dan
krit ik unt uk perbaikan makalah ini akan sangat dinant ikan.
RHIZOBACTERIA PENDUKUNG PERTUMBUHAN TANAMAN
Plant Growth Promotor Rhizobacteria
Pendahuluan
Pertumbuhan tanaman adalah proses terjadinya peningkatan jumlah dan ukuran daun dan batang. Hasil pertumbuhan tanaman adalah produk yang dapat dikonsumsi atau dimanfaatkan menjadi produk lain, atau hanya bersifat estetis. Pengambilan hasil dinamakan pemanenan, yang dapat dilakukan oleh manusia, hewan atau peralatan mesin.
Setiap proses pertumbuhan memerlukan energi. Tanaman mendapatkan energinya dari matahari melalui proses fotosintesis, yang merupakan proses penyerapan cahaya oleh pigmen hijau (klorofil) dalam daun. Energi cahaya, air dan CO2 menghasilkan O2 dan gula sederhana. Tanaman kemudian memanfaatkan gula sederhana ini untuk mensintesa gula yang lebih kompleks serta karbohidrat untuk disimpan sebagai energi yang dapat digunakan kembali jika dibutuhkan untuk mensintesa selulosa dan hemiselulosa pada dinding sel, atau menggabungkannya dengan nitrogen untuk mensintesa protein. Bagaimana tanaman memanfaatkan energi ini bergantung pada stadia pertumbuhan tanaman dan kondisi lingkungan (Rayburn, 1993).
Proses pertumbuhan tajuk dan akar merupakan proses yang saling berkaitan satu sama lain. Apabila terjadi gangguan pada salah satunya maka akan menyebabkan gangguan pada bagian lainnya. Misalnya pada kondisi kekurangan air dan nitrogen, pertumbuhan tajuk lebih mengalami hambatan daripada bagian akar. Hal ini disebabkan akar bertugas lebih banyak untuk mencari air dan sumber N dari dalam tanah untuk didistribusikan ke bagian tajuk. Pada saat ketersediaan air memadai maka pertumbuhan tajuk kembali ke arah normal sehingga distribusi fotosintat ke akar juga kembali normal.
Tanaman membutuhkan sedikitnya 13 unsur hara untuk pertumbuhan dan perkembangannya. Beberapa unsur berada dalam bentuk tersedia dalam semua jenis tanah, sedangkan lainnya dalam bentuk tidak tersedia sehingga membutuhkan tambahan dari luar tanah dalam bentuk pemupukan. Unsur hara ini berperan sebagai nutrisi bagi tanaman, sedangkan sistem yang mengendalikan pertumbuhan dan perkembangan tanaman adalah substansi kimia yang konsentrasinya sangat rendah, yang disebut substansi pertumbuhan tanaman, hormon pertumbuhan tanaman (fitohormon), atau pengatur pertumbuhan tanaman (plant growth regulator / PGR) (Gardner dkk., 1991).
Pengatur Pertumbuhan Tanaman
yang berarti suatu substansi yang disintesis dalam suatu organ yang pada gilirannya merangsang terjadinya respon pada organ yang lain. Hormon tanaman tidak khusus seperti hormon hewan dalam hal organ tempat sintesisnya ataupun organ tempat responnya, tetapi cenderung mengikuti pola tingkah-laku yang umum.
PGR pada saat ini dibagi menjadi 5 kelas: auksin, giberelin, sitokinin atau kinin, penghambat pertumbuhan (inhibitor), dan etilen. Dua hormon terakhir secara kimiawi tidak dapat masuk ke dalam salah satu dari 5 kategori di atas. Keduanya diisolasi berturut-turut dari biji sejenis tanaman kol (Brassica napus) dan tanaman tingkat tinggi tertentu lainnya. Substansi-substansi ini mungkin masih membutuhkan revisi dalam sistem klasifikasi yang berlaku saat ini. Banyak analogi dari kebanyakan hormon yang ada dalam 5 kelas ini yang diproduksi secara sintesis dan banyak di antaranya memperoleh pemanfaatan yang penting dalam bidang pertanian.
Sifat-sifat tertentu diperlukan bagi suatu senyawa agar dapat dikelompokkan sebagai fitohormon: (1) tempat sintesis berbeda dari tempat aktivitas (misalnya sintesis di pucuk dan daun muda, tetapi responnya pada batang, akar atau organ-organ lainnya); (2) respon dihasilkan oleh jumlah yang sangat kecil (yaitu konsentrasinya bisa sekecil 10-9 M); (3) tidak seperti pada vitamin dan enzim, respon mungkin berbentuk formatif dan plastik (tidak terpulihkan, misalnya respon trophy).
Seringkali pemasokan fitohormon secara alami di bawah optimal, dan dibutuhkan sumber dari luar untuk menghasilkan respon yang dikehendaki. Jumlah auksin di atas optimal umumnya bertindak sebagai herbisida. Umumnya suatu fitohormon bertindak secara sinergis dengan hormon-hormon lainnya dalam meningkatkan suatu respon.
Secara umum PGR menghasilkan respon yang sifatnya sebagai berikut: (1) auksin merangsang pertumbuhan dengan cara pemanjangan sel dan menyebabkan dominansi ujung; (2) giberellin meningkatkan pertumbuhan meristem samping dalam daun dan antar buku; (3) sitokinin merangsang pertumbuhan dengan cara pembelahan sel; (4) penghambat pertumbuhan (inhibitor) mengerdilkan pemanjangan dan mempercepat absisi dan penuaan; dan (5) etilen meningkatkan pematangan buah dan pertumbuhan horisontal.
Respon pengatur pertumbuhan pada tanaman tidak selalu berupa pertumbuhan secara fisik, namun juga perbaikan dalam proses fisiologi tanaman. Misalnya pada akar adanya PGR meningkatkan kemampuan akar dalam memfiksasi nitrogen, menyerap fosfor dalam kondisi ketersediaan terbatas, dan sebagainya. PGR yang dapat memperbaiki proses fisiologi tanaman melalui akar biasanya bersifat eksogen atau berasal dari luar tanaman. PGR ini berasal dari dalam tanah, khususnya dari interaksi akar tanaman dengan organisme yang ada dalam tanah.
Akar Tanaman
Akar merupakan organ vegetatif utama yang memasok air, mineral dan bahan-bahan yang penting untuk pertumbuhan dan perkembangan tanaman. Pertumbuhan akar yang kuat umumnya diperlukan untuk kekuatan dan pertumbuhan tajuk tanaman. Apabila akar mengalami kerusakan karena gangguan secara biologis, fisik atau mekanis sehingga mengurangi fungsinya maka pertumbuhan tajuk juga akan terganggu.
Fungsi akar bagi tanaman adalah: (1) penyerapan; (2) penambatan (anchorage); (3) penyimpanan; (4) transport; dan (5) perbanyakan (propagation). Akar juga merupakan sumber utama beberapa PGR bagi tanaman tertentu.
Penyerapan air dan mineral terutama terjadi melalui ujung akar dan bulu akar, walaupun bagian akar yang lebih tua dan lebih tebal juga menyerap sebagian. Akar yang lebih tua memainkan fungsi yang diperlukan untuk transport dan penyimpanan bahan, yang beranalogi dengan transport bahan dari dan ke daun melalui batang dan percabangan. Akar dikotil seringkali berfungsi sebagai organ utama penyimpan cadangan makanan.
Perakaran dari sejumlah spesies tanaman dapat digunakan untuk perbanyakan karena kemampuannya untuk membentuk pucuk tambahan dan menyimpan cadangan makanan yang mendukung pertumbuhan pucuk baru tersebut. Selain itu akar dapat menghasilkan PGR berupa giberellin dan sitokinin, yang mempengaruhi pertumbuhan dan perkembangan secara keseluruhan.
Panjang akar merupakan hasil perpanjangan sel-sel di belakang meristem ujung; sedangkan lebar akar yang lebih daripada pembesaran sel-sel ujung merupakan hasil dari meristem lateral atau pembentukan kambium, yang memulai pertumbuhan sekunder dari meristem kambium. Pertumbuhan panjang dan lingkar akar umumnya beranalogi dengan pertumbuhan panjang dan lingkar pada tajuk, tetapi pada percabangan lateral tidak terdapat analogi antara bagian tajuk dengan akar. Percabangan akar muncul dari lingkaran tepi yang jauh di dalam jaringan tua atau jaringan yang berdiferensiasi, berbeda dengan percabangan tajuk yang muncul dari ujung dan berasal dari permukaan.
yaitu kelembaban, temperatur, kandungan nutrisi, bahan-bahan toksin, kekuatan agregat dan agen biologis.
Umumnya karakteristik akar dikendalikan oleh sejumlah gen, sehingga terdapat perbedaan antar genotipe. Perbedaan genetik ini kemudian berinteraksi dengan lingkungan tanah. Mekanisme kendali genetik untuk perakaran sangat kompleks, tetapi seperti pada tajuk tanaman, ditunjukkan juga adanya kerja hormon pertumbuhan. Auksin (Indole Acetic Acid) meningkatkan pertumbuhan akar hanya bila dalam konsentrasi rendah.
Kebutuhan akan auksin dibuktikan dengan diperlukannya faktor daun pada perbanyakan dengan cara stek agar bakal tanaman dapat membentuk akar. Banyak spesies yang membutuhkan beberapa jaringan daun atau pucuk yang aktif, untuk menghasilkan pengatur pertumbuhan yang dapat berdifusi. Suatu kofaktor akar, yang berhasil dipisahkan dan diidentifikasi sebagai katekol dan pirogalol, bekerja secara sinergis dengan IAA untuk memacu pembentukan akar.
Rizosfer
Gambar 2. Perakaran (kiri), perbesaran mikroskop daerah perakaran / rizosfer (kanan)
Efek rizosfer selain tampak dalam bentuk melimpahnya jumlah mikroorganisme juga dalam adanya distribusi bakteri yang memiliki ciri mempunyai kebutuhan khusus, yaitu asam amino, vitamin-vitamin B, dan faktor pertumbuhan khusus (kelompok nutrisional). Laju kegiatan metabolik mikroorganisme rizosfer itu berbeda dengan laju kegiatan metabolik mikroorganisme dalam tanah non-rizosfer.
Hiltner pada tahun 1904 menggambarkan rizosfer sebagai bagian dari tanah yang secara langsung dipengaruhi oleh substansi yang dikeluarkan dari akar ke dalam larutan tanah, sehingga tercipta kondisi yang menyenangkan bagi bakteri tertentu (Bruehl, 1987). Ia juga menggambarkan adanya organisme yang merugikan di sekitar akar dari tanaman yang sakit dan organisme yang bermanfaat di sekitar akar dari tanaman yang sehat. Fakta biologi utama dari rizosfer atau daerah yang dipengaruhi akar adalah jumlah yang banyak dan aktivitas yang tinggi dari mikroorganisme tanah dalam area ini dibandingkan dengan tanah tanpa akar. Di antara dua area ini terdapat area transisi di mana pengaruh akar menurun seiring dengan jarak. Biasanya daerah rizosfer merupakan lapisan tipis yang tetap menempel pada akar setelah tanah di sekitar akar dihilangkan dengan cara menggoyangkan perakaran (Katznelson, 1965, dalam Bruehl, 1987).
sebelah luar dari akar itu sendiri (endorizosfer). Selain menghasilkan efek biologi, akar juga mempengaruhi sifat kimia dan sifat fisika tanah, sehingga secara tidak langsung mempengaruhi mikroorganisme tanah.
Clark (1942 in Bruehl, 1987) menyatakan rizoplan adalah habitat khusus atau lokasi aktivitas mikrobia. Rizoplan atau permukaan akar mendukung terjadinya aktivitas biologi yang tinggi serta memiliki sensitivitas yang tinggi terhadap pengaruh akar pada mikroflora dan mikrofauna tanah. Analisa terhadap struktur halus atau lapisan epitel dari perakaran tanaman setelah diinokulasi dengan bakteri khusus menunjukkan bahwa bakteri menjadi lekat pada permukaan perakaran dengan bantuan dari lapisan eksternal yang bersifat musilagen atau disebut ‘musigel’ yang secara normal terdapat pada sistem perakaran yang sedang aktif tumbuh.
Rasio rizosfer terhadap tanah (R : S) dapat digunakan untuk memperkirakan perubahan dalam populasi mikroba yang disebabkan pertumbuhan tanaman. Rasio R : S dihitung dengan membagi jumlah mikroorganisme dalam rizosfer tanah dengan jumlah mikroorganisme dalam tanah yang bebas dari pertumbuhan tanaman. Hasilnya dapat dinyatakan berdasarkan berat akar bersama dengan tanah yang melekat padanya. Efek rizosfer yang lebih besar dijumpai lebih banyak karena bakteri (nilai R : S memiliki rentangan dari 10 hingga 20 atau seringkali lebih) daripada karena actinomycetes atau jamur. Sedangkan karena protozoa atau alga hanya dapat dilihat perubahan yang sangat kecil.
mencapai hasil, yang mana ditegaskan bahwa hasil tanaman budidaya dibatasi hanya oleh lingkungan fisik alamiah tanaman dan potensial genetik bawaan.
Umumnya rizosfer dari kebanyakan tanaman mengandung bakteri Gram-negatif, tidak berspora, berbentuk batang, dan terdapat pada daerah rizoplan.
Beberapa genus bakteri ini adalah Pseudomonas, Arthrobacter, Agrobacterium, Azotobacter, Mycobanterium, Flavobacterium, Cellulomonas, Micrococcus, dsb.,
ditemukan dalam jumlah yang banyak namun ada juga yang tidak ditemukan sama sekali. Bakteri yang membutuhkan asam amino lebih banyak terdapat di daerah rizoplan dan daerah rizosfer dibandingkan tanah di luar rizosfer. Actinomycetes penghasil antibiotik lebih banyak terdapat dalam rizosfer dibandingkan tanah tanpa rizosfer.
Rizosfer dapat mengalami perubahan, di antaranya diakibatkan oleh: (1) penambahan tanah; (2) pemberian nutrisi melalui daun; dan (3) inokulasi artifisial biji atau tanah yang mengandung sediaan mikroorganisme hidup, terutama bakteri. Banyak percobaan telah dilakukan untuk meneliti pengaruh penambahan pupuk N, P, dan K terhadap mikroflora rizosfer. Hasilnya masih belum dapat digenuslisasikan karena penambahan maupun penurunan R : S telah dilaporkan terjadi sebagai suatu akibat dari penggunaan pupuk.
Jumlah rizosfer meningkat pada tanah-tanah yang kering dibandingkan pada tanah-tanah basah. Temperatur dan kelembaban secara langsung berpengaruh terhadap mikroorganisme, dan secara tidak langsung terhadap tanaman. Pengaruh tidak langsung inilah yang kelihatannya lebih penting. Beberapa organisme secara nyata dapat langsung beradaptasi dengan rizosfer, namun dalam keberhasilannya membentuk koloni dengan akar dipengaruhi oleh adanya kompetisi dengan organisme lain dan kondisi tanamannya (Bruehl, 1987).
Ketergantungan satu mikroorganisme terhadap mikroorganisme lain dalam hal produk ekstra-selular, terutama asam amino dan faktor perangsang pertumbuhan, dapat dianggap sebagai suatu efek asosiatif dalam rizosfer. Beberapa penelitian menunjukkan adanya peningkatan kandungan asam amino dalam tanaman yang ditumbuhkan pada tanah yang diinokulasi dengan mikroorganisme khusus. Pengamatan serupa dilakukan dalam hal pengaruhnya terhadap peningkatan vitamin-B, auksin, giberellin, dan antibiotik. Diketahui bahwa senyawa giberellin dan yang serupa giberellin dihasilkan oleh genus-genus bakteri yang umumnya dijumpai di dalam rizosfer, seperti Azotobacter, Arthrobacter, Pseudomonas dan Agrobacterium.
Sekresi antiobiotik oleh mikroorganisme dan penghambatan pertumbuhan secara biologis terhadap mikroorganisme lain yang peka, ditemukan terjadi baik dalam penanaman di lapangan maupun dalam kultur murni. Efek antagonistik dalam rizosfer ini diharapkan terjadi secara alami bahkan dalam tanah yang tidak dibudidayakan. Namun dari segi agronomi adanya penghambatan yang berlebihan terhadap pertumbuhan Azotobacter atau Rhizobium di daerah perakaran akan menyebabkan penurunan fiksasi nitrogen atau pembentukan bintil akar.
Proses-proses Mikrobia dalam Rizosfer
terjadi karena adanya stimulasi dalam rizosfer, meskipun manfaatnya bagi tanaman tidak selalu nyata. Proses-proses tersebut dijelaskan dalam uraian berikut ini:
Sifat Asosiasi yang Memfiksasi Nitrogen
Bakteri pemfiksasi nitrogen yang tidak bersimbiosis biasanya terdapat dalam rizosfer dan di bawah kondisi nitrogen yang terbatas memungkinkan terjadinya kompetisi yang lebih menguntungkan dari mikroorganisme lain. Tidak terdapat bukti yang jelas yang dapat mendukung stimulasi selektif dari organisme ini, dan bakteri ini kelihatannya tidak menginvasi akar. Bagaimana pun, terdapat asosiasi spesifik antara Azotobacter paspali dengan Paspalium notatum, serta antara Azospirillum sp dengan akar sereal.
Kelangsungan asosiasi rizosfer-pemfiksasi nitrogen bergantung pada pasokan karbon yang dapat dioksidasi serta efisiensi dari konversinya. Fiksasi nitrogen termasuk “boros” karena membutuhkan ATP dan efisiensi untuk organisme bebasnya berkisar dari 4 g C g-1 N dalam Azospirillum brasiliense sampai dengan 174 g C g-1 N dalam Aerobacter aerogenes. Hanya senyawaan karbon sederhana yang dapat digunakan. Jika diasumsikan bahwa konversi dari efisiensi karbon berasal dari akar adalah 10 g C g-1 N; bakteri pemfiksasi nitrogen meliputi 10% populasi rizosfer; dan keseluruhan karbon yang hilang dari akar adalah merupakan ketersediaan bagi seluruh bakteri, maka jika 150 g C m-2 adalah hilang dari akar potensi maksimum pemfiksasi nitrogen hanyalah 1,5 g N m-2.
Perubahan dalam Ketersediaan Nutrisi
Dalam kondisi pasokan nutrisi yang rendah, populasi rizosfer akan bersaing dalam memperebutkan beberapa jenis nutrisi sehingga kemudian mereduksi pasokan nutrisi tersebut bagi tanaman. Sebagai contoh, sejumlah fosfat tersedia bagi tanaman dapat menjadi berkurang atau malah meningkat, bila dibandingkan dengan tanah tanpa rizosfer. Mangan dalam entuk b Mn2+ dapat teroksidasi menjadi mangandioksida yang dapat larut dalam rizosfer, menyebabkan defisiensi mangan pada tanaman gandum, meskipun larutan kultur bakteri rizosfer menghasilkan senyawaan (ionophores) yang mendukung pengambilan mangan oleh akar.
Imobilisasi nutrisi dapat terjadi dalam rizosfer, disebabkan oleh adanya materi berasal dari akar yang memiliki rasio C : N yang tinggi. Hal ini dapat menjadi berguna untuk nutrisi yang mobil seperti NO3- yang dapat tercuci dari daerah perakaran. Terlebih lagi bagian yang banyak dari bakteri tanah adalah bakteri fakultatif anaerob dan respirasi oleh keduanya serta oleh populasi rizosfer dapat mereduksi potensi redoks dengan baik untuk memungkinkan terjadinya denitrifikasi. Bagaimana pun evapotranspirasi dapat menyebabkan rizosfer menjadi kering, meningkatkan difusi oksigen dan pengambilannya oleh tanaman dapat memindahkan nitrat dari daerah yang memiliki potensi mendenitrifikasi. Ketersediaan oksigen juga dapat ditingkatkan melalui sel-sel aerenkhim di dalam tanaman sehingga memungkinkan difusi ke dalam rizosfer, terutama pada tanaman akuatik.
Produksi Hormon Tumbuh
Mikroorganisme rizosfer menghasilkan senyawaan seperti growth hormon dan phytotoxin yang dapat mempengaruhi pertumbuhan tanaman. Keanekaragaman substrat dalam rizosfer yang tersedia untuk pertumbuhan tanaman menunjukkan banyaknya produk yang bermanfaat.
yang mendukung keberadaannya dalam tanah. Pengukuran dalam tanah menjadi sulit pada saat konsentrasi senyawaan sangat rendah serta dihasilkan secara lokal. Identifikasi senyawaan ini biasanya dilakukan melalui bioassay yang hanya dapat mendeskripsikannya sebagai senyawaan “serupa auksin”. Beberapa senyawaan, misalnya auksin dan etilen, menghambat pertumbuhan tanaman pada satu konsentrasi tetapi menstimulir pertumbuhan pada suatu konsentrasi yang rendah. Kebanyakan jenis utama hormon tanaman dapat dihasilkan oleh bakteri dan fungi.
Asam indole asetat (IAA) adalah suatu auksin yang diproduksi dari triptofan. Enzim ini terdapat dalam rambut akar yang menggulung pada akar legum yang disebabkan adanya rizobia tertentu, dan juga dimetabolisir oleh bakteri tanah sehingga keberadaannya dalam tanah akan bergantung pada tingkat akumulasinya. Di dalam kondisi anaerob, etilen dapat terbentuk pada konsentrasi yang cukup dapat menghambat perpanjangan akar sereal. Selain juga diperoduksi dari metionin, IAA juga dimetabolisir oleh mikroorganisme. Sedikit informasi yang ada mengenai sitokinin dan asam absisat dalam tanah.
Efek fitotoksik dari suatu tanaman yang ditanam sebelumnya dalam suatu sistem pertanaman, dipengaruhi oleh penguraian mikroba terhadap residu tanaman. Apabila efek dari suatu mikroorganisme terjadi pada tahap perkecambahan benih, maka tidak mudah untuk membedakan efek metabolik dari efek fisik seperti misalnya konsentrasi oksigen tereduksi. Penelitian sebelumnya menunjukkan bahwa asam dihidrostearat dan vanilin adalah fitotoksin utama, tetapi asam alifatik seperti asetat dan asam oksalat juga dihasilkan oleh bakteri dan fungi, terutama dalam kondisi anaerob yang juga mencegah penguraiannya. Substratnya berasal dari bahan organik, pupuk hijau, residu tanaman dan limbah hewan. Sedikit sekali diketahui efek asam aromatik seperti p-hidroksibenzoat dan asam p-kumarat yang telah ditemukan dalam tanah. Kebanyakan senyawaan fenolik pada akhirnya menjadi bagian dari fraksi asam humat pada bahan organik tanah.
dapat menghambat atau merangsang pertumbuhan tanaman secara langsung atau secara tidak langsung.
Gambar 3. Produksi antibiotik secara in situ dari mikroba tanah dan rizosfer.
Secara tidak langsung antibiotik ini menghambat pertumbuhan dengan cara pemindahan mikroorganisme patogen yang mengganggu pertumbuhan. Jika senyawaan ini dihasilkan, maka dapat dinonaktifikan dengan cara penjerapan oleh liat atau penguraian oleh mikroorganisme. Senyawaan mengandung sulfur yang tidak stabil seperti hidrogen sulfida dihasilkan oleh bakteri pereduksi sulfat, dalam rizosfer dapat menimbulkan toksisitas pada tanaman padi.
Alelopati
Telah banyak terdapat laporan penelitian mengenai interaksi inhibitor atau stimulator antar tanaman, dan antara tanaman dengan mikroorganisme. Interaksi ini dinamakan alelopati. Sebagai contoh couchgrass (Agropyron repens), suatu gulma yang ditemukan di banyak negara, mengurangi pertumbuhan gandum (Triticum aestivum) dan lucerne (Medicago sativa). Vegetasi di sekitar kacang hitam / black
walnut (Juglans nigra) sangat jarang, dan hal ini disebabkan adanya suatu substansi bersifat racun yang dihasilkan tanaman kacang hitam untuk tanaman di dekatnya.
Terdapat serangkaian bukti yang menunjukkan bahwa nitrifikasi dipengaruhi oleh bahan kimia yang bersifat alel yang dihasilkan oleh beberapa spesies tanaman dan mikroorganisme. Juga terdapat bukti bahwa tingkat nitrifikasi sangat menurun pada saat tahap pertumbuhan tanaman menuju tahap vegetasi maksimum.
Bakteri sebagai Mikroorganisme Tanah
Organisme yang menghuni tanah meliputi mikroorganisme, tanaman dan hewan. Adanya organisme hidup dalam tanah menyebabkan perubahan biokimia dalam tanah, dan untuk memahami caranya dalam mempengaruhi fungsi-fungsi tanah maka diperlukan informasi aktivitas organisme tersebut. Hal ini termasuk reaksi-reaksi yang dilakukan oleh organisme, interaksi yang terjadi antar organisme dan antara organisme dengan lingkungannya (Wood, 1989).
Mikroorganisme yang menghuni tanah dapat dikelompokkan menjadi bakteri, actinomycetes, jamur, alga dan protozoa. Bakteri merupakan kelompok mikroorganisme tanah yang paling dominan dan mungkin meliputi separuh dari biomassa mikroba dalam tanah. Bakteri terdapat dalam segala macam tipe tanah tetapi populasinya menurun dengan bertambahnya kedalaman tanah. Secara umum profil horizon A terdiri dari lebih banyak mikroorganisme daripada horizon B dan C. Dalam kondisi anaerob, bakteri mendominasi tempat dan melaksanakan kegiatan mikrobiologi dalam tanah karena jamur dan actinomycetes tidak dapat tumbuh baik tanpa adanya oksigen (Subba Rao, 1994).
genetik antara sel donor dan penerima pada pasangan gandaannya (Paul and Clark, 1989).
Pengelompokkan terhadap bakteri dapat dilakukan antara lain berdasarkan reaksinya dengan penanda/pewarna Gram, yang berdasarkan komponen dinding sel di mana bakteri yang menyerap pewarna dikelompokkan sebagai bakteri Gram-positif; sedangkan bakteri yang tidak menyerap pewarna dikelompokkan sebagai bakteri Gram-negatif. Pengelompokkan juga dapat dilakukan berdasarkan proses fisiologi,
yaitu autochtonous bagi bakteri yang pertumbuhannya terjadi secara lambat dalam tanah yang tidak mengandung substrat yang mudah dioksidasi, serta zymogenous bagi bakteri yang pertumbuhan dan aktivitasnya cepat pada saat residu segar ditambahkan ke dalam tanah.
Pertumbuhan bakteri dalam kondisi keberadaan oksigen dan tidak ada oksigen, juga digunakan sebagai kriteria untuk membedakan bakteri menjadi: anaerobik (tidak ada oksigen); aerobik (ada oksigen); dan anaerobik fakultatif (tidak
ada oksigen atau ada oksigen). Bentuk sel bakteri adalah khas berbentuk bola seperti batang atau spiral, berukuran panjang 1,5 – 2,5 µm dan diameter 0,5 – 1,0 µm. Jumlah bakteri dalam 1 g tanah bervariasi dari 106 – 109, tetapi tidak terdistribusi secara merata dalam tanah melainkan mengambil tempat dalam koloni kecil, seringkali berasosiasi dengan sumber dari substrat organik (misalnya akar tanaman).
Salah satu dari karakteristik penting bakteri sebagai suatu kelompok adalah berbagai manfaat biokimia yang dimilikinya. Suatu organisme seperti Pseudomonas sp mampu memetabolisir berbagai jenis bahan kimia termasuk pestisida, sedangkan
Nitrobacter sp hanya mampu menghasilkan energi dari oksdasi nitrit menjadi nitrat.
Thiobacillus ferrooxidans mendapatkan energi dari oksidasi senyawa sulfur tereduksi
dan dari ion-ion ferro, dan memiliki nilai pH optimum untuk pertumbuhan sekitar 2. Clostridium sp mampu untuk tumbuh dalam kondisi ketiadaan oksigen dan dapat
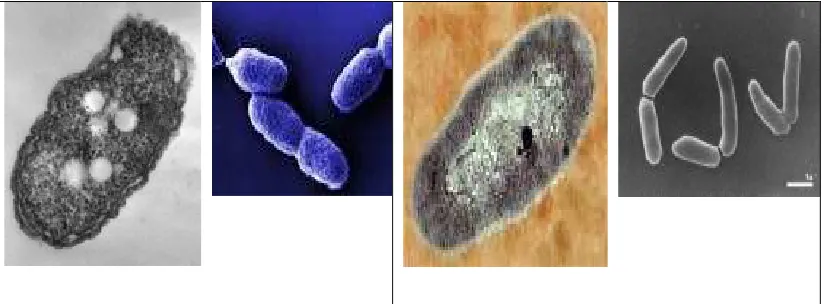
Gambar 4. Pseudomonas sp hasil pembesaran mikroskop electron (kiri), transmisi mikroskop electron dari bakteri PGPR Azospirillum brasiliense (Az) yang menempati satu ruang sel yang sama (kanan).
Dalam sistem Bergey mengenai Bakteriologi Determinatif (Bergey’s Manual of Determinative Bacteriology), bakteri diklasifikasikan menjadi kelompok
taksonomi ordo, famili, genus, dan spesies berdasarkan konsep Linneaeus klasik yaitu tata nama ganda (binomial nomenklatur). Sepuluh ordo termasuk ke dalam kelas Schizomycetes. Dari 10 ordo tersebut, 3 di antaranya yaitu Pseudomonales, Eubacteriales dan Actinomycetales berisi spesies bakteri yang seringkali dijumpai dalam tanah.
Dari 190 genus bakteri yang terdapat dalam manual Bergey, 97 genus (57%) terdiri dari spesies yang diperkirakan sebagai bakteri tanah. Sebagai contoh, Arthrobacter sp., membentuk sekitar setengah dari total koloni bakteri yang tumbuh
Bakteri tanah yang paling umum termasuk dalam genus Pseudomonas, Arthrobacter, Clostridium, Achromobacter, Bacillus, Micrococcus, Flavobacterium,
Corynibacterium, Sarcina dan Mycobacterium. Kelompok bakteri lain yang umum
dijumpai dalam tanah adalah myxobacteria yang termasuk genus Myxococcus, Chondrococcus, Archangium, Polyangium, Cytophaga dan Sporocytophaga. Dua
genus terakhir termasuk selulolitik dan karenanya dominan dalam lingkungan yang kaya selulosa. Myxobacteria menjadi predator bagi bakteri Gram-negatif lainnya melalui proses lisis.
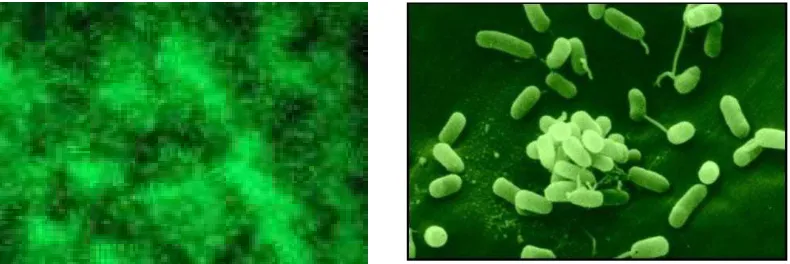
Gambar 5. Contoh genus bakteri yang ada dalam tanah, Azotobacter (kiri) dan Arthrobacter (kanan)
Dalam tanah selain terdapat bakteri yang menguntungkan, juga terdapat bakteri yang merugikan atau bersifat pathogen. Sebagai contoh adalah Clostridium sp yang umumnya terdapat dalam tanah dan kotoran hewan. Bakteri ini merupakan organisme anaerobik yang menghasilkan spora, dan beberapa spesies seperti C. tetani dan C. perfringens adalah penyebab tetanus dan gas gangren. Penyakit ini dapat bersifat
letal.
Bakteri juga digolongkan berdasarkan caranya memperoleh makanan. Bakteri autotrof dapat mensintesis sendiri kebutuhan makanannya, sedangkan bakteri
heterotrof bergantung dari makanan yang sudah terbentuk sebelumnya untuk
berlawanan dengan bakteri kemoautotrof yang mengoksidasi bahan anorganik untuk memperoleh energi dan pada waktu bersamaan memanfaatkan karbon dari CO2 untuk pertumbuhannya.
Pada kelompok bakteri kemoautotrof termasuk juga kelompok bakteri kemoautotrof obligat yang lebih menyukai substrat khusus tertentu. Contohnya adalah Nitrobacter yang memanfaatkan nitrit; Nitrosomonas yang memanfaatkan amonium; Thiobacillus yang mengubah senyawa belerang anorganik menjadi sulfat; dan Ferrobacillus yang mampu mengubah besi fero menjadi feri.
Bakteri Akar (Rhizobacteria)
Selama dasawarsa terakhir istilah “rhizobacteria” digunakan untuk menggambarkan bakteri rizosfer yang membentuk koloni dengan akar (Schroth and Hancock, 1982 dalam Kloepper, et al., 1985). Kolonisasi akar adalah suatu proses di mana bakteri bertahan melakukan inokulasi ke dalam benih tanaman atau ke dalam tanah, penggandaan diri dalam spermosfer dalam responnya terhadap eksudat benih yang kaya akan karbohidrat dan asam amino, menempel pada permukaan akar, dan mengkoloni sistem perakaran yang sedang berkembang.
Berbagai manfaat positif dari bakteri dalam rizosfer telah menjadikannya sumber potensial bagi ketersediaan nutrisi dalam tanah serta mendorong pertumbuhan tanaman sehingga menjadi lebih baik. Beberapa bakteri tanah berasosiasi dengan akar tanaman budidaya dan memberikan pengaruh yang bermanfaat pada tanaman inangnya. Bakteri ini dikelompokkan ke dalam PGPR (Plant Growth Promoting Rhizobacteria). Strains PGPR yang sering ditemukan di antaranya Pseudomonas fluorescent.
diperoleh oleh tanaman yang bersimbiosis dengan legum diperoleh oleh tanaman tanpa simbiosis legum dengan bakteri tanah lainnya.
Selama 70 tahun ke belakang, penggunaan inokulan mikrobia lebih banyak melibatkan rizobia dan tanaman legum. Hak cipta pertama kali bagi inokulan rizobia diberikan pada awal abad ini, kemudian diikuti dengan eksploitasi secara komersial. Adanya berbagai kendala dan ketidaksesuaian yang ditimbulkan oleh rizobia hasil produksi massal mengarahkan penelitian pada pengembangan perbaikan strain Rhizobium yang akan memperbaiki rizobia asal tanah dan membentuk nodul yang
efektif dalam jumlah besar pada tanaman. Selain itu teknik aplikasinya dimodifikasi untuk mendistribusikan inokulum yang viabilitasnya tinggi pada perkecambahan benih. Banyak hambatan ekologis membatasi keberhasilan rizobia yang diinokulasikan pada tanah (Young and Burns, 1993).
Seringkali diasumsikan bahwa peningkatan pertumbuhan tanaman setelah inokulasi adalah respon langsung terhadap bakteri yang diinokulasikan. Dalam hal rizobia, penampakkan nodul didampingkan dengan pengukuran sensitif terhadap fiksasi nitrogen menggunakan 15N dan teknik reduksi asetilen, memungkinkan terjadinya korelasi yang jelas antara respon tanaman dengan inokulan. Bagaimana pun masih terdapat sebab dan akibat yang membingungkan pada saat mengkaji fungsi dari inokulan mikrobia lainnya.
Rizobia Pendukung Pertumbuhan Tanaman (PGPR)
beberapa tahun yang lalu, membawa pada beberapa sistem penandaan yang baur yang dapat diaplikasikan untuk mengukur kolonisasi akar oleh bakteri, termasuk Azospirillum.
Pada saat mengembangkan konsep mengenai kolonisasi akar, sangat penting untuk diperhatikan bahwa kapasitas kolonisasi akar adalah spesifik-strain. Oleh karenanya tidak semua pseudomonas fluoresens menunjukkan kolonisasi akar, dan salah satu yang harus diekstrapolasi dari laporan individual mengenai kolonisasi oleh suatu strain adalah semua strain dari takson yang sama akan berkoloni dengan akar. Maka koloni dengan akar mewakili suatu pengelompokkan dari total komunitas bakteri rizosfer, dan kelompok ini dinamakan rizobakteri.
Istilah ”kolonisasi akar” dapat digunakan untuk menggambarkan proses-proses yang secara fundamental berbeda. Ada suatu persetujuan umum di antara para peneliti bakteri bahwa kolonisasi akar adalah suatu proses aktif, bukan suatu peristiwa yang kebetulan ditemukan antara bakteri tanah dan akar tanaman. Kolonisasi akar adalah suatu proses di mana bakteri diinokulasikan ke dalam benih atau tanah, dapat bertahan hidup dan menggandakan diri dalam spermosfer dalam responsnya terhadap eksudat benih yang kaya akan nutrisi, berasosiasi dengan permukaan akar dan mengkoloni sistem akar yang sedang berkembang dalam tanah dengan mikroflora asli tanah.
Rizobia dapat menghasilkan 1 dari 3 jenis efek pada tanaman inang yang diinokulasi: menghilangkan, menetralkan atau bermanfaat. Rizobakteri yang bermanfaat dinamakan Plant Growth-Promoting Rhizobacteria (PGPR). Oleh karena itu, PGPR dapat dipertimbangkan secara fungsional sebagai bakteri bermanfaat yang mengkolonisasi akar.
pertumbuhan dan hasil dari tanaman kentang, tetapi gagal mempengaruhi pertumbuhan tanaman yang ditumbuhkan dalam kondisi gnotobiotic. Dan growth promotion yang terjadi pada kondisi tanah lapang berkaitan dengan reduksi populasi rizoplan asli, yaitu fungi dan bakteri.
Pertumbuhan tanaman distimulasi PGPR secara tidak langsung dengan cara mereduksi aktivitas organisme lainnya, sehingga dinamakan biokontrol. Sebaliknya, beberapa strain PGPR mendukung pertumbuhan tanaman secara langsung dalam ketiadaan mikroflora asli rizosfer. Meskipun inhibisi dari mikroflora asli tidak terlibat dengan growth promotion, biokontrol dapat terjadi pada saat PGPR diuji dalam kajian penyakit atau pada percobaan lapang dengan patogen asli.
Biokontrol pada beberapa kasus diperkirakan muncul akibat dari penyakit yang terbebaskan. Akar menunjukkan pemanjangan atau percabangan yang berlebih akibat perlakuan PGPR, dapat meloloskan infeksi dari fungi patogen asal tanah yang lebih mudah menginfeksi benih muda. Selain itu infeksi patogen yang terlokalisir dalam 1 area sistem perakaran mungkin diseimbangkan oleh suatu peningkatan global dalam biomassa akar sebagai kompensasi.
Apabila dilakukan evaluasi PGPR dalam penelitian lapangan atau tanah lapangan yang disimpan dalam penelitian greenhouse, memungkinkan untuk menggambarkan efek yang teramati dari PGPR pada tanaman inang secara prinsip sebagai pendukung pertumbuhan atau biokontrol dengan mencatat perkembangan pertumbuhan tanaman dan simptom yang terjadi selama pertumbuhan tanaman.
Gambar 6. Scanning elektron micrograph Pseudomonas fluoresens
Kisaran Tanaman Inang bagi PGPR
Selama 5 tahun yang lalu penelitian PGPR dilanjutkan dengan tambahan 3 tujuan. Pertama, pekerjaan yang telah dilakukan pada tanaman “tanpa akar” sebagai tanaman inang menunjukkan bahwa mayoritas tanaman kondusif terhadap induksi PGPR terhadap pertumbuhan. Tujuan kedua melibatkan karakterisasi dampak spesifik PGPR, yaitu dampak lain selain mendukung hasil produksi. Studi pada tujuan kedua ini telah mengarahkan pada keberadaan 2 sub-kelas baru dari PGPR dan menunjukkan bahwa PGPR dapat juga digunakan sebagai agen biokontrol. Tujuan ketiga dari pekerjaan ini membuktikan bahwa beberapa strain PGPR dapat mendukung pertumbuhan tanaman secara langsung, yaitu dengan meniadakan kehadiran mikroorganisme patogenik atau yang merugikan.
Tanaman inang bagi bakteri PGPR memiliki kisaran yang cukup luas, di antaranya adalah :
Barley
Kedelai
Strain Pseudomonas putida mengkolonisasi akar lateral dan akar utama tanaman kedelai (Phaseolus vulgaris L.) dalam kultur hidroponik. Dihasilkan peningkatan kadar lignin dalam akar, bobot tanaman meningkat dalam perlakuan P. putida setelah diinokulasi dengan Fusarium solani f. sp. phaseoli.
Kanola
Potensi untuk mendapatkan peningkatan hasil pada kanola (Brassica campestris L dan B. napus L.) melalui perlakuan PGPR dilaporkan pada tahun 1988. Lebih dari 4000 strain bakteri dikumpulkan dari zona akar dan secara individu dievaluasi untuk tumbuh pada temperatur 4 – 14oC, metabolisme eksudat benih, kemotaksis terhadap aspargin dan kolonisasi akar. 887 dari strain ini diuji kemampuan growth promotor-nya dalam percobaan green house menggunakan tanah dari lapangan. 35 strain meningkatkan area daun, 13 strain meningkatkan hasil sampai 57% selama 2 tahun, 3 strain meningkatkan hasil 6 – 13% selama 2 tahun. Strain PGPR yang diidentifikasi dalam pengujian ini termasuk P. putida, P. fluorescens, Serratia liquefaciens, P. putida biovar B, dan Arthrobacter citreus.
Kapas
Dua strain dari P. fluorescens yang ditapis (di-screening) untuk antagonisme secara in vivo pada 2 patogen tanaman, jamur dan bakteri, meningkatkan bobot tanaman 8 –
40% pada tanaman kapas berusia 4 minggu dalam percobaan green house dengan tanah lapangan.
Jagung
mengkoloni akar pada kepadatan populasi rata-rata Log 3 cfu/cm akar dan mempengaruhi peningkatan hasil dari 3 – 3,5 bu/acre dibandingkan dengan kontrol pada berbagai lokasi sekitar 5 tahun.
Kacang-kacangan
Strain A-13 dari Bacillus subtilis diketahui dapat meningkatkan pertumbuhan tanaman kacang di lapangan. Penelitian berikutnya mengindikasikan bahwa strain mempengaruhi peningkatan produksi sekitar 14 – 24%. Strain A-13 ini tidak seperti kebanyakan strain Bacillus sp., di mana A-13 adalah pengkoloni akar. Kolonisasi akar berkaitan dengan peningkatan keseluruhan pertumbuhan tanaman, pertumbuhan akar yang lebih cepat dan lebih tersedianya nutrisi tanaman. Oleh karena itu strain A-13 ini dinyatakan termasuk dalam golongan PGPR.
Padi
Sakthivel et al. (1986) mengisolasi strain P. fluorescens dari rizosfer berbagai tanaman dan strain terseleksi menunjukkan spektrum lebar secara in vitro dalam hal antibiosis terhadap fungi dan bakteri patogen. Pada saat strain ini dilapiskan pada benih padi yang ditanam dalam pot menggunakan tanah dari lapangan, 4 strain menginduksi peningkatan tinggi tanaman sekitar 12 – 14% lebih tinggi.
Tanaman sayuran
bobot kering pucuk dan akar dalam percobaan di rumah kaca dengan tanah lapangan pada tomat, ketimun, jagung manis, wortel, dan seledri. Tim peneliti di Kalifornia menyelidiki pengaruh PGPR pada seledri (M.N. Schroth, komunikasi pribadi). Tiga puluh bakteri yang mengkolonisasi akar, termasuk yang dikonfirmasi sebagai PGPR pada tanaman lainnya, ditapis langsung di lapangan untuk peningkatan pertumbuhan seledri dengan Fusarium oxysporum f.sp. apii yang secara alami ada di lapangan. Empat strain dipilih untuk digunakan dalam tiga percobaan tindak lanjut yang diulang tiga kali. Tercatat bahwa ada spesifitas genotipik yang sangat kuat dalam respons terhadap inokulasi PGPR. Satu strain PGPR menstimulasi peningkatan yang signifikan dalam pertumbuhan awal (peningkatan bobot kering dan/atau segar) pada ketiga percobaan dan peningkatan panen yang signifikan, yaitu 12 sampai 15% lebih besar daripada kontrol pada dua percobaan. Strain yang sama tidak memiliki pengaruh ketika diuji pada kultivar yang lain.
Kolonisasi Rizosfer (Rhizosphere Colonization)
Plant growth-promoting rhizobacteria (PGPR) pertama kali diteliti oleh Kloepper dan Scroth (1982) untuk menggambarkan bakteri tanah yang mendiami daerah perakaran tanaman yang dinokulasikan ke dalam benih dan ternyata meningkatkan pertumbuhan tanaman. Proses kolonisasi selengkapnya adalah sebagai berikut: kemampuan mempertahankan diri (survive) dari proses inokulasi ke dalam benih, penggandaan diri dalam spermosfer (daerah di sekeliling benih) dalam responsnya terhadap eksudat benih, penyerangan terhadap permukaan akar, dan berkolonisasi / mendiami daerah perkaran untuk memperkuat sistem perakaran (Nelson, 2004).
spesifik komponen permukaan, kemampuan dalam menggunakan komponen spesik dari eksudat akar, sekresi, dan quorum sensing. Genus mutan mengubah ekspresi dari sifat ini dan membantu pemahaman kita mengenai aturan yang tepat yang masing-masing memegang peranan dalam proses kolonisasi ini.
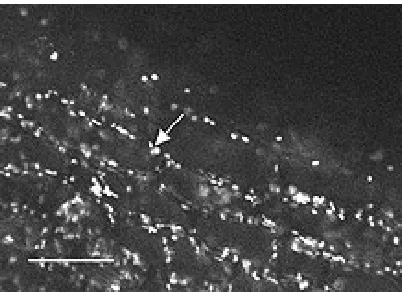
Gambar 7. Hasil penapisan (screening) laser mikrograph dari akar kanola yang baru 5 hari dikolonisasi oleh Pseudomonas putida strain 6-8, ditandai dengan protein fluoresens hijau (ditunjukkan oleh arah panah). Garis putih di pojok kiri bawah berukuran 60 µm.
Kemajuan dalam sistem identifikasi yang baru, di mana sebelumnya gen yang tidak berkarakter dibuat menggunakan metode penapisan (screening) nonbias yang mengandalkan teknologi perpaduan gen (gene fusion). Metode ini menggunakan reporter transposons dan teknologi ekspresi in vitro (IVET) untuk mendeteksi ekspresi gen selama kolonisasi (www.plantmanagementnetwork.org).
Mekanisme Kerja PGPR
PGPR meningkatkan pertumbuhan tanaman dengan cara langsung dan tidak langsung, tetapi mekanisme spesifiknya tidak hanya melibatkan karakter yang baik (Glick, 1995). Mekanisme langsung dari plant growth promotion oleh PGPR dapat diperlihatkan dalam ketiadaan patogen tanaman. Atau mikroorganisme rizosfer lainnya, sedangkan mekanisme tidak langsung melibatkan kemampuan PGPR dalam menurunkan pengaruh yang merusak / mengganggu dari patogen tanaman terhadap hasil tanaman budidaya.
Pengaruh PGPR secara langsung dalam meningkatkan pertumbuhan tanaman terjadi melalui bermacam-macam mekanisme, di antaranya fiksasi nitrogen bebas yang ditransfer ke dalam tanaman, produksi siderophore yang meng-khelat besi (Fe) dan membuatnya tersedia bagi akar tanaman, melarutkan mineral seperti fosfor dan sintesis phytohormon. Peningkatan langsung dari pengambilan mineral melalui peningkatan dalam spesifik flux ion di permukaan tanaman karena keberadaan PGPR ini telah juga dilaporkan. Strains PGPR bisa jadi menggunakan satu atau lebih mekanisme ini dalam rizosfer. Telah diketahui bahwa PGPR mensintesis auksin dan sitokinin atau terlibat dalam sintesis etilen tanaman.
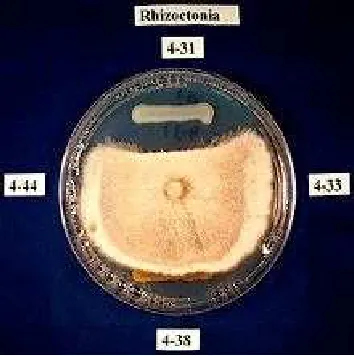
Gambar 8. Contoh kajian in vitro terhadap penghambatan (inhibisi) pertumbuhan fungi. Isolat bakteri yang berbeda diuji kemampuan inhibisinya terhadap pertumbuhan Rhizoctonia sp., penyakit tular tanah pada tanaman legum. Daerah inhibisi terlihat sekitar isolat 4-31 pada bagian atas pelat agar.
Indikasi adanya mekanisme kerja yang mendukung pertumbuhan oleh PGPR adalah pada saat strain bakteri meningkatkan pertumbuhan secara tidak langsung dengan cara mengubah keseimbangan mikrobia dalam rizosfer. Siderofor pengkhelat Fe, antibiotik, dan HCN diproduksi oleh beberapa PGPR dan telah dikaitkan dengan kemampuannya mereduksi patogen tanaman serta rizobakteria yang bersifat toksik. Kaitan HCN dalam mendukung pertumbuhan secara langsung melalui penemuan bahwa beberapa rizobakteria yang bersifat toksik menghasilkan HCN, yang menghambat pertumbuhan tanaman dan bahwa rizobakteria yang merugikan ini dapat dihambat oleh beberapa strain PGPR (Schippers, 1988, dalam Kloepper, et al., 1985).
Interaksi dengan Bakteri Pelarut Fosfat
zone pengurasan fosfat mengelilingi akar. Jamur mikoriza arbuskular berkontribusi terhadap pengambilan P dan ketersediaannya melalui mata rantai biotik dan geokimia dalam ekosistem tanah sehingga berpengaruh pada kecepatan siklus P dan polanya dalam sistem pertanian dan ekosistem alamiah. Fosfat tersedia karena bakteri pelarut fosfat berperan dengan cara menghemat pelarutan sumber fosfat yang tidak bisa diraih oleh permukaan akar untuk membatasi difusi, dimaksudkan bahwa jika fosfat terlarut diambil oleh miselium mikoriza arbuskula, interaksi mikrobia sinergistik seharusnya memperbaiki ketersediaan P dalam tanaman (Barea et.al., 1983).
Simbiosis Rhizobium dengan Tanaman Legum
Anggota tertentu komunitas rizosfer dapat memasuki hubungan simbiotik dengan tanaman inang membentuk mikoriza dan nodul akar. Nodul dibentuk pada akar tanaman aktinoriza oleh Francia sp. dan pada akar legum oleh Rhizobium sp. dan Bradyrhizobium sp. Bakteria tersebut memperoleh karbohidrat dari tanaman inang dan memasok tanaman dengan senyawa nitrogen yang diperoleh dari nitrogen di atmosfer. Legum merupakan komponen penting dari komunitas tanaman alami dan sistem pertanian. Sebagian besar informasi yang tersedia adalah mengenai legum pertanian.
Peran legum dalam sistem pertanian
Mayoritas genus dan spesies legum adalah di daerah tropis, dan tempatnya pada jaman dahulu mungkin di tanah tercuci di hutan tropis. Legum temperat dan subtropis berkembang dari sini. Beberapa darinya, terutama yang termasuk dalam famili Trifolieae, seperti Trifolium sp. dan Medicago sp, dan famili Viceae, seperti Pisum sp. dan Cicer sp. telah beradaptasi dengan tanah netral atau berkapur yang memiliki nutrisi lebih tinggi. Sebagian besar legum yang ditanam di pertanian temperat, yang menerima perhatian lebih besar, termasuk ke dalam kelompok tersebut dan peran legum di daerah lainnya tidak dipelajari secara luas. Tanaman legum pada umumnya dapat dibagi ke dalam tanaman legum yang ditanam karena bijinya yang kaya akan protein, seperti biji kedelai (Glycine max) dan kacang merah (Cicer arietinum) dan legum pakan ternak, seperti semanggi putih (T. repens) dan kudzu (Pueraria phaseoloides). Kacang kedelai saat ini merupakan tanaman legum bulir yang dominan dengan produksi tahunan 108 ton di seluruh dunia.
Nodul hanya dapat secara aktif memfiksasi nitrogen apabila tanaman tersebut dipasok secara memadai dengan semua unsur yang penting untuk pertumbuhan. Legum memiliki kebutuhan tambahan yang tinggi akan molibden, komponen dari nitrogenase. Legum sering kali ditanam di tanah asam, terutama di daerah tropis dan subtropis, yang memiliki konsentrasi H+, Al3+, dan Mn2+ tinggi dan konsentrasi fosfat dan Ca2+ rendah, dapat membatasi pertumbuhan, nodulasi, dan fiksasi nitrogen. Legum yang mengandalkan fiksasi nitrogen umumnya lebih sensitif terhadap kemasaman daripada legum sama yang memanfaatkan NH4+ dan NO3- (mineral N). Legum bernodul secara efektif dapat tumbuh dengan baik di tanah dengan kandungan mineral nitrogen rendah; bagaimanapun juga, konsentrasi mineral nitrogen tinggi dapat menghalangi nodulasi dan fiksasi nitrogen di atmosfer, sehingga menghilangkan manfaat legum.
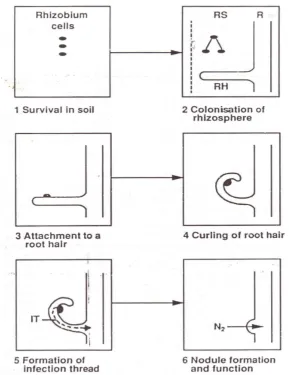
Infeksi
termasuk ke dalam genus Rhizobium dan Bradyrhizobium, yang mengakibatkan pembentukan nodul akar. Hingga saat ini, keberadaan nodul baru diperiksa hanya pada 48% genus legum dan pada 86% dari 48% ini ditemukan nodul. Tidak semua legum dinodulasi oleh semua rizobia. Sebagai contoh, rizobia lucerne (Medicago sativa) hanya menodulasi lucerne dan bukan legum lainnya, dan rizobia yang
menodulasi legum lainnya tidak menodulasi lucerne. Pengelompokkan legum yang dinodulasi oleh spesies bakteri yang sama menimbulkan konsep spesifitas inokulasi silang.
Dalam Bergey’s Manual of Systematic Bacteriology (1984), klasifikasi awal bakteri nodul sebagai genus tunggal, Rhizobium, yang dibagi ke dalam spesies menurut spesifitas inokukasi silang dimodifikasi untuk mencakup dua genus. Rhizobium sekarang dikasifikasikan sebagai genus spesies yang berkembang cepat
dan menghasilkan asam di media laboratorium, yang terutama menodulasi legum temperat. Genus yang baru, Bradyrhizobium, terdiri atas spesies yang berkembang lambat dan menghasilkan alkalin di media laboratorium, yang terutama menodulasi legum tropis.
Dalam keadaan non-simbiotik, rizobia biasanya Gram-negatif, tidak membentuk spora, berbentuk bulat, hidup secara saprofit pada kisaran yang lebar dari sumber karbon organik, tetapi tidak dapat memfiksasi nitrogen. Dengan adanya akar legum yang sesuai, rizobia ini mampu mengatasi mekanisme pertahanan tanaman, menginfeksi akar, membentuk nodul, mensintesis nitrogenase dan komponen penting lainnya serta mengikat nitrogen.
terlihat pada tanaman legum sebagai inang, kemungkinan disebabkan oleh produksi IAA oleh rizobia dari triptofan yang dikeluarkan oleh akar. Derajat penggulungan rambut akar yang terlihat nyata (360o atau lebih) merupakan karakteristik dari interaksi yang kompatibel.
Gambar 9. Diagram skematik pembentukan nodul pada kebanyakan tanaman legum.
semanggi putih (T. repens) dan R. leguminosarum biovar trifolii. Suatu tahap kedua dari penempelan diperkirakan dengan kuat menempelkan rizobia pada permukaan rambut akar, kemungkinan disebabkan produksi selulosa mikrofibria selular. Rizobia yang menempel seringkali terjadi pada saat rambut akar menggulung, hal ini membawa pada peningkatan konsentrasi sinyal kimia dari mikrobia secara lokal serta respon yang mengikutinya dari tanaman.
Bagian akar yang terinfeksi hanya sedikit dan bervariasi, selain itu kebanyakan infeksi (68 – 99% dalam Trifolium sp.) gugur sebelum mencapai bagian dasar rambut akar. Benang-benang infeksi merupakan struktur tubular yang membawa rizobia, seringkali dalam berkas tunggal, dari titik masuk ke dalam rambut akar ke sel-sel bagian dalam pada korteks akar. Rizobia dapat menyebabkan perubahan arah pertumbuhan dinding sel tanaman inang sehingga rambut akar dapat menerima penetrasi rizobia. Cara lainnya, eksopolisakarida rizobia dapat menstimulasi suatu peningkatan aktivitas enzim pektat tanaman (poligalakturonase) yang menyebabkan pelunakan dinding sel rambut akar sehingga memungkinkan terjadinya penetrasi oleh rizobia. Pembentukan benang infeksi dapat dianggap sebagai pengendalian terhadap reaksi perlawanan patogen tanaman inang.
Tidak semua tanaman legum diinfeksi melalui rambut akar. Pada tanaman kacang tanah (Arachis hypogaea), legum mimosoid umumnya dan batang bernodul Sesbania sp., infeksi terjadi pada akar lateral yang baru muncul, atau secara langsung
melalui epidermis. Karena kebanyakan legum tidak memiliki rambut akar dan juga nodul yang berasosiasi dengan akar lateral, umumnya infeksi sering terjadi dalam sel-sel epidermal.
Pembentukan Nodul
kemampuan menggandanya menghilang, mulai mensintesis nitrogenase serta dikelilingi oleh membran yang dibentuk oleh dinding sel tanaman inang. Bakteroid terendam dalam suatu larutan yang mengandung leghaemoglobin, yang mentransport oksigen untuk respirasi pada tekanan parsial yang sangat rendah, yang dengan cara ini memproteksi nitrogenase sensitif-oksigen. Leghaemoglobin, yang serupa dengan haemoglobin, menyebabkan nodul berwarna merah muda. Tahanan difusi dari nodul juga menahan tingkat difusi oksigen ke dalam nodul.
Nodul mungkin saja berbentuk bulat, silindris, datar, dan seringkali bidentat (berlubang di kedua sisinya) atau dengan percabangan coralloid (tertutup / terisolasi), atau mungkin saja bentuknya tidak beraturan. Dalam sebuah nodul mungkin terdapat 1010 bakteroid, biasanya mengandung sebuah strain tunggal Rhizobium, meskipun terjadi juga penempatan oleh strain ganda. Seiring usia nodul, sel-sel kehilangan leghaemoglobinnya dan terbentuk sebuah vakuola besar diikuti dengan nekrosis. Belumlah jelas benar apakah rizobia yang viable dilepaskan ke dalam tanah mengikuti kematian nodul, berasal dari bakteroid ataukah dari sel-sel yang tersisa dalam benang infeksi yang tidak membentuk nodul.
Strain rizobia yang membentuk nodul pada legum dinamakan infektif. Strain yang kemampuan fiksasi nitrogennya tinggi dinamakan efektif, dan strain yang kemampuan fiksasi nitrogennya rendah dinamakan inefektif. Bagaimana pun, meski legum dinodulasi oleh strain yang sangat efektif, dan secara berkelanjutan mengambil sejumlah porsi nitrogen dari tanah dalam bentuk NO3- dan NH4+, pasokan nitrogen akan tetap tersedia. Strain yang sama dapat menunjukkan perbedaan tingkat efektivitas pada spesies atau varietas tanaman inang yang berbeda. Akar yang diinfeksi oleh strain inefektif memiliki nodul yang lebih kecil dan lebih pucat dibandingkan yang terinfeksi oleh strain efektif.
inokulasi-silang tidak dapat dihindari untuk tahap awal penempelan strain pada akar. Polisakarida Rhizobium nampaknya penting dalam menentukan kesesuaian legum, di mana pada mutan strain ini kekurangan eksopolisakarida normal yang memperlihatkan infeksi dan nodulasi yang kurang baik.
Pada Rhizobium, gen-gen yang menyebabkan rambut akar menggulung, pembentukan benang infeksi dan induksi nodul spesifik tanaman inang, ditempatkan pada sejumlah besar Symplasmids indigenous, yang juga membawa gen-gen untuk pembentukan nitrogenase (nif) dan fiksasi nitrogen (fix). Pada Bradyrhizobium, analogi fungsional dari gen-gen nod, nif dan fix ini dilakukan oleh kromosom. Empat gen nodulasi, nod D, A, B, C adalah berdekatan dan sangat terlindungi. Gen-gen ini mengendalikan penggulungan rambut akar, dan nod D adalah suatu regulator gen yang konstitutif (selalu dihasilkan daripada menjadi penginduksi) dalam Rhizobium leguminosarium biovar trifolii.
Gen-gen nod adalah teratur dan harmonis, melalui nod D, dengan cara bersenyawa dalam eksudat akar legum. Pada semanggi putih, diproduksi hidroksiflavon pada zone yang dapat diinfeksi dari munculnya rambut akar,
menstimulasi ekspresi gen nod dalam hitungan menit, sedangkan coumarins dan isoflavon diproduksi di belakang ujung akar menahan transkripsi gen nod.
Keberhasilan dari suatu infeksi tertentu bergantung pada rasio senyawaan penstimulir dan penghambat (inhibitor), yang melengkapi tanaman dengan suatu mekanisme pengaturan nodulasi. Saat nod D berinteraksi dengan flavonoid, gen nod lainnya seperti nod F dan nod E yang terlibat dalam kisaran spesifik tanaman inang diinduksi, dan pada akhirnya meregulasi gen-gen non-symplasmid lainnya sedemikian rupa seperti yang terlibat dalam produksi poligalakturonase.
terjadinya kekeringan seringkali menyebabkan tanaman mengurangi nodul-nodulnya. Nodul dari tanaman atau pepohonan legum dapat bertahan hidup selama beberapa tahun. Nodul dapat juga berumur pendek apabila diparasiti oleh larva serangga seperti hama polong (Sitonia lineata). Parah atau ringannya serangan dapat dikurangi dengan melakukan irigasi pada musim kering, sehingga dapat memperpanjang umur nodul.
Ekonomi karbon dari legum
Bakteri dalam nodul harus dipenuhi kebutuhan energinya agar dapat memfiksasi nitrogen, dan kemudian juga membutuhkan oksigen untuk mengoksidasi karbohidrat. Nodul dan sistem akar yang berasosiasi dengannya menerima 15 – 30% dari nett fotosintat yang diproduksi oleh legum. Meskipun hampir tidak mungkin untuk mengukur “biaya” fungsi nodul secara individual, teori perkiraan menduga bahwa aktivitas nitrogenase adalah “biaya” utama dalam nodul yang memanfaatkan sistem respirasi. Sebagian dari “biaya” ini sehubungan dengan reduksi proton menjadi gas hidrogen oleh enzim sementara mereduksi nitrogen. Beberapa strain rizobia memiliki suatu sistem pengambilan hidrogen (Hup = hydrogen uptake) yang mendaur ulang hidrogen melalui oksidasinya menjadi air. Nodul memiliki sistem Hup yang harus memanfaatkan fotosintat secara lebih efisien; bagaimana pun belum ada keuntungan hasil bersih yang diketahui.
Ammonium (NH4+) diproduksi melalui fiksasi, diekskresikan ke dalam sel sitoplasma tanaman inang di mana ia kemudian berasimilasi dan digunakan untuk mensintesis nitrogen organik untuk transport dalam tanaman. Legum dapat dikelompokkan berdasarkan bentuk nitrogen yang dikirim keluar dari nodul. Legum-legum dari daerah temperat seperti semanggi (Trifolium sp.) dan lucerne (Medicago sativa) mengirim keluar amida seperti asparagin dan glutamin, sedangkan legum
daerah tropis seperti Phaseolus sp. dan kedelai (Glycine max) mengirim keluar ureida seperti allantoin dan asam allantoic. Transport ureida kemungkinan lebih efisien daripada transport amida dalam pengertian mol-C yang digunakan dalam sintesis.
Sehubungan pentingnya legum dalam pertanian dan ekosistem alami, maka tidak ada pendugaan yang dapat diandalkan dari jumlah nitrogen difiksasi dalam kondisi lapangan. Tingkat fiksasi untuk beberapa legum telah diketahui, tetapi pendugaan ini beragam, sebagai contoh tingkat fiksasi N 4,5 – 67,3 g m-1 yr-1 pada tanaman semanggi putih (Trifolium repens), dan sangat bergantung pada metode pengukuran yang digunakan.
Inokulan Rizobium
Karakteristik strain Bradyrhizobium adalah tumbuh dengan lambat, Gram-negatif, merupakan bakteri tanah. Genus Bradyrhizobium mewakili kelompok heterogen dari bakteri bernodul dalam hubungan taksonomi yang tidak begitu dimengerti (Jordan, 1984 dalam FNCA, 2006). Genus Bradyrhizobium saat ini terdiri dari 6 spesies, yaitu : Bradyrhizobium japonicum, Bradyrhizobium elkanii, Bradyrhizobium liaoningense, Bradyrhizobium yuanmingense, Bradyrhizobium
betae, dan Bradyrhizobium canariense.
Isolasi strain Rizobium
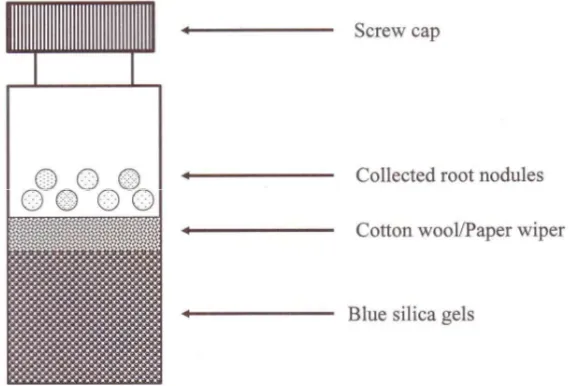
1. Koleksi dan konservasi (penyimpanan) nodul akar di lapangan. Peralatan untuk koleksi dan konservasi nodul akar :
Vial (tabung) sampling dengan dessicated silica gel : Untuk perjalanan ke lapangan selama lebih dari 2 hari, nodul akar yang dikumpulkan harus dicegah dari proses dekomposisi dan invasi mikroorganisme tanah yang tercampur dengan prosedur isolasi selanjutnya. Oleh karena itu, nodul akar yang dikumpulkan disimpan dalam sebuah vial (tabung) dengan dessicated silica gel. Jika warna gel berubah menjadi merah muda, dessicant harus diganti oleh yang baru yang berwarna biru.
Gambar 10. Vial sampling untuk nodul akar.
Sekop yang kuat dibutuhkan untuk mengkoleksi nodul akar dari legum liar, karena tanah tempat tumbuhnya sangat keras. Karena itu dibutuhkan sebuah sekop yang kuat untuk menggali ke atas akar legum.
Hal-hal yang harus dipertimbangkan sebelum melakukan sampling :
a. identifikasi lokasi
b. identifikasi tanaman inang c. identifikasi tanah.
Koleksi dan perawatan dari nodul akar :
Gali seluruh tanaman untuk mendapatkan nodul akar; pindahkan dengan hati-hati tanah di sekitar nodul akar; nodul akar yang terlihat dapat dikoleksi dengan forsep. Seluruh nodul dari sebuah tanaman inang tunggal mewakili 1 unit dari bahan yang dikoleksi dan disimpan dalam vial yang sama. Nodul akar dari tanaman berbeda tetapi berasal dari 1 spesies tidak dapat disatukan karena masing-masing menggambarkan lingkungan tanah berbeda meskipun jika hanya terpisah beberapa meter. Vial yang berisi nodul akar kering disimpan dalam pendingin pada suhu 4oC sampai dilakukan isolasi bradyrhizobia.
2. Isolasi Bradyrhizobia dari nodul akar kering.
Hari pertama : cuci nodul akar kering di bawah air mengalir untuk membuang kontaminasi tanah, kemudian pindahkan nodul akar yang bersih ke air steril.
Biarkan nodul akar dalam air steril dalam pendingin pada suhu 4oC semalaman agar menyerap air.
Hari kedua : sterilisasi permukaan nodul akar dengan cara mencelupkannya dalam ethanol 70% untuk 30 detik, dan dalam larutan Na-hipoklorit 3% untuk 3 menit. Nodul akar selanjutnya dicuci sekurangnya 5 kali dengan air steril. Setiap nodul akar dihancurkan dalam sebuah tabung microfuge dengan larutan gliserol 15% sebanyak 100 µl. 10 µl dari suspensi berlumpur dengan larutan gliserol 15% ditempatkan dalam formasi baris ke permukaan media ekstrak yeast mannitol agar.
Dua minggu kemudian : Pelat diinkubasi pada suhu 28oC selama 2 minggu dalam posisi terbalik. Koloni tunggal yang berhasil diisolasi ditempatkan ulang ke dalam pelat baru untuk memperoleh kultur yang murni.
Gambar 11. Isolasi Bradyrhizobium dari nodul akar kering. Nodul akar kering ditempatkan dalam silika gel biru (kiri), koloni Bradyrhizobium berasal dari nodul akar tanaman Vigna liar yang berhasil diisolasi (kanan).
3. Isolasi dari nodul segar.
flask. Larutan merkuri klorida (0,1% berat/volume) atau larutan dari hidrogen peroksida (3% w/v) dapat digunakan untuk sterilisasi nodul.
Nodul tersebut dihancurkan di dalam tabung steril dengan menggunakan batang kaca steril dan air steril. Campuran nodul dan air tersebut kemudian diencerkan dan ditempatkan dalam formasi baris di atas permukaan piring YMA (Yeast-Mannitol Agar) mengandung Red Congo. Piring Petri yang diinokulasi diinkubasi pada 25-28 °C selama 3 sampai 10 hari, bergantung pada strain dan spesies hingga koloni muncul. Koloni rhizobia berlendir, bundar, dan tidak atau sedikit menyerap Red Congo. Isolat dari satu koloni rhizobia tunggal kemudian dimurnikan dan dikonfirmasi sebagai Rhizobium dengan mendemonstrasikan kemampuan membentuk nodul pada legum inang uji dalam kondisi terkendali (otentikasi isolat).
Bakteri Non Simbiotik pada Rizosfer dan Jamur
Istilah rizobakteria digunakan untuk menggambarkan bagian dari bakteri rhizosfer yang mampu berkoloni dalam lingkungan perakaran (Kloepper et al., 1985). Koloni akar yang bermanfaat, rizosfer bakteria, PGPR ditentukan oleh tiga (3) karakteristik intrinsik yakni :
1. Bakteri harus mampu berkolonisasi di dalam akar.
2. Bakteri harus dapat mempertahankan diri / survive dan menggandakan diri dalam mikro habitatnya yang berasosiasi dengan perakaran tanaman, dalam kompetisinya dengan mikrobiota lainnya, paling tidak pada waktu diperlukan untuk memperlihatkan kativitas perlindungannya,
3. Bakteri harus mendukung/meningkatkan pertumbuhan tanaman.
PGPR diketahui berperan dalam proses ekosistem penting, seperti kontrol biologi dari patogen tanaman, siklus nutrisi / unsur hara dan atau pertumbuhan benih (Persello-Cartieaux et al., 2003; Barea et al., 2004; Zahir et al., 2004). Pseudomonas dan Bacillus merupakan genus yang hampir sebagian besar digambarkan memiliki PGPR, tetapi banyak juga takson lainnya juga berisi PGPR.
PGPR dibagi dalam dua kelompok, yakni PGPR yang terlibat dalam siklus nutrisi / unsur hara dan phytostimulasi, dan PGPR yang terlibat dalam biokontrol dari patogen tanaman (Bashan dan Holguin, 1998). Proses dalam PGPR melibatkan siklus nutrisi / unsur hara termasuk hubungannya dengan fiksasi nitrogen non simbiotik dan perannya untuk meningkatkan ketersediaan fosfat dan nutrisi / unsur hara lainnya di dalam tanah. Banyak bakteria diazotroph asimbiotik telah banyak diketahui dan dicoba sebagai biofertilizer (Kennedy et al.,2004). Beberapa hasil penelitian tidak sampai pada kesimpulan, tetapi cukup mendorong untuk memperbaiki prosedur seleksi dan produksi kualitas inokulan untuk aplikasi praktis. Seleksi PGPR diazotroph yang efektif adalah sangat penting untuk perkembangan selanjutnya dari teknologi ini.
2000; Richardson, 2001; Vessey et al., 2004). Phosphate solubilizing bacteria (PSB) telah diidentifikasi, tetapi keefektifannya dalam sistem tanah-tanaman masih belum jelas (Barea, et al., 2002a). Pertama, inokulasi PSB harus berada pada akar yang berasosiasi dengan habitat tanahnya. Karenanya direkomendasikan bahwa inokulasi PSB dipilih dari populasi PGPR yang ada untuk mengambil keuntungan dari kemampuannya untuk berkolonisasi pada lingkungan mikro rizosfer.
Kedua, kemampuan inokulasi PSB untuk menyediakan P bagi tanaman bisa jadi terbatas, salah satunya akibat senyawa yang dilepaskan oleh PSB untuk melarutkan fosfat secara cepat menurun atau karena fosfat terlarut diikat kembali sebelum fosfat itu mencapai permukaan akar. Bagaimanapun, jika fosfat dilepaskan oleh PSB diambil oleh miselium mikoriza, hasilnya akan menjadi interaksi mikrobia sinergis kooperatif yang dapat memperbaiki perolehan P oleh tanaman.
Spesies Azospirilium juga dipertimbangkan menjadi PGPR (Okon, 1994; Bashan, et al., 1990). Aktivitas yang signifikan dari bakteri ini adalah produksi auksin – merupakan tipe fitohormon yang berpengaruh pada morfologi akar dan dengan demikian, memperbaiki pengambilan unsur hara dalam tanah. Ini mungkin bisa lebih penting daripada aktivitas pengikatan nitrogen (Dobbelaere et al., 1999). Spesies Azospirilium sudah digunakan sebagai inokulan pada benih di bawah kondisi lapang (Dobbelaere et al., 2001). Walaupun banyak studi melaporkan keuntungan dari inokulasi Azospirillium, beberapa di antaranya menampilkan hasil yang tidak konsisten. Namun demikian kita dapat menetapkan bahwa dengan manajemen praktek yang sesuai, penggunaan inokulan ini akan bermanfaat bagi nutrisi tanaman.
(van Loon et al., 1998). Hal ini dicapai melalui pelepasan antibiotik oleh PGPR. Di antara faktor antifungal yang diproduksi oleh PGPR, acetyphloroglucinols (Landa et al., 2003; Picard et al., 2004) dan phenacines (Chin-A-Woeng et al., 2003; Ownley
et al., 2003) merupakan produk yang menerima perhatian terbanyak.
Beberapa mikroorganisme menguntungkan bagi tanaman dengan beberapa cara, contohnya spesies Trichoderma mengontrol patogen jamur melalui perannya sebagai mikrobia antagonis dan menyebabkan terlokalisasi dan respons pertahanan sistemik tanaman (Harman et al., 2004). Endophytic bakteria dan jamur berperan sebagai peningkat pertumbuhan dan sebagai agen biokontrol (Harman et al., 2004). Baru-baru ini terdapat postulat bahwa mekanisme tambahan untuk pertumbuhan tanaman yang dinaikkan oleh PGPR terjadi karena perubahan komunitas mikrobial rizosfer (Ramos et.al., 2003). Mekanisme tidak langsung akan menarik untuk dievaluasi pengaruh nyatanya dari aktivitas ini dalam biologi rizosfer.
Tantangan dalam Seleksi dan Karakterisasi PGPR
suppresive terhadap patogen itu (Weller, et al., 2002). Pendekatan lain yang dilibatkan dalam seleksi berdasarkan ciri / sifatnya yang diketahui berasosiasi dengan PGPR seperti kolonisasi akar, 1-aminocyclopropane-1-carboxylate (ACC) aktivitas deaminasi (Glick, et al., 1995; Giacomodonato, et al., 2001), antibiotik dan produksi siderofore. Perkembangan dari high throughput assay systems and effective bioassays akan memfasilitasi seleksi dari superior strains.
Tantangan dalam Aplikasi PGPR di Lapangan
Pencapaian penampilan yang konsisten di lapangan, di mana terdapat heterogenitas faktor biotik dan abiotik serta kompetisi dengan organisme indigenous, adalah lebih sulit. Pengetahuan mengenai faktor ini dapat membantu dalam penentuan konsentrasi yang optimal, waktu dan penempatan inokulan dan manajemen tanah dan tanaman yang strategis untuk meningkatkan ketahanan dan perkembangbiakan inokulan (Bowen and Rovira, 1999). Kosep dari teknik atau pengelolaan rizosfer untuk meningkatkan fungsi PGPR dengan memanipulasi tanaman inang (host plant), substrat untuk PGPR atau melalui praktek agronomis adalah untuk memperoleh perhatian yang meningkat.
Peningkatan formulasi yang lebih baik untuk meningkatkan ketahanan dan aktivitasnya di lapangan dan kesesuaian dengan perlakuan kimia dan biologi pada benih adalah perhatian pada lokasi lainnya; pendekatan termasuk di dalamnya optimisasi kondisi pertumbuhan sebelumnya untuk perumusan dan peningkatan dari perbaikan carriers dan aplikasi teknologi (Bashan, 1998; Bowen and Rovira, 1999; Date, 2001).
Teknik nuklir dan teknik terkait lainnya digunakan untuk mengevaluasi keefektifan inokulan dalam hubungannya dengan tanaman inang dan tanah Sejumlah formula telah menunjukkan kemampuan memfiksasi N asosiatif sebesar 15 – 21% dengan teknik isotop 15N. Sementara sterilisasi bahan pembawa pupuk hayati digunakan radiasi 60Co. Alur produksi dalam penelitian ini adalah sebagai berikut :
Hasil tanaman pada perlakuan kontrol (kiri) dan pada perlakuan Azora (kanan).
Hasil tanaman pada perlakuan 10 ton pupuk kandang (kiri) dan pada perlakuan 10 ton pupuk kandang+Azora (kanan). Gambar 12. Hasil penelitian aplikasi PGPR Azora pada tanaman sayuran.
Prospek yang Akan Datang