Major clades in tropical
Agaricus
Ruilin Zhao&Samantha Karunarathna&Olivier Raspé&Luis A. Parra& Jacques Guinberteau&Magalie Moinard&André De Kesel&Gérard Barroso& Régis Courtecuisse&Kevin D. Hyde&Atsu K. Guelly&Dennis E. Desjardin& Philippe Callac
Received: 17 February 2011 / Accepted: 3 September 2011 #Kevin D. Hyde 2011
Abstract Agaricus(Basidiomycota) is a genus of saprobic fungi that includes edible cultivated species such as Agaricus bisporus, the button mushroom. There has been considerable ecological, nutritional and medicinal interest in the genus, yet the extent of its diversity remains poorly known, particularly in subtropical and tropical areas. Classification of tropical species has for a large part followed the classification of temperate species. The objective of our study was to examine to what extent this system of classification is appropriate for tropicalAgaricus species. Species from temperate sections were therefore compared to the major clades of tropical species using a phylogenetic approach. ITS1+2 sequence data from 128 species were used in the phylogenetic analysis. Specimens
included four species of genera closely related toAgaricus, 38 temperate species representing the eight classical sections of the genus, and 86 putative species of Agaricus from tropical areas of Africa, Asia and the Americas. Bayesian and maximum likelihood analyses produced relatively congruent trees and almost identical clades. Our data show that (i) only about one-third of tropical species belong to the classical sections based on temperate species; the systematics of the genus therefore needs to be expanded; (ii) among the remaining two-thirds of tropical species, those from the Americas and those from Africa and/or Asia group in distinct clades, suggesting that secondary diversification occurred in these two areas; (iii) in contrast, several clades of classical sections contain
R. Zhao
Key Laboratory of Forest Disaster Warning and Control in Yunnan Province, Faculty of Conservation Biology, Southwest Forestry University,
Kunming 650224, China
S. Karunarathna
:
K. D. HydeSchool of Science, Mae Fah Luang University, Chiang Rai 57100, Thailand
O. Raspé
:
A. De KeselNational Botanic Garden of Belgium, Domein van Bouchout,
1860 Meise, Belgium
L. A. Parra
Avda. Padre Claret 7, 5º G,
09400 Aranda de Duero, Burgos, Spain
J. Guinberteau
:
M. Moinard:
G. Barroso:
P. Callac (*) INRA,UR1264, Mycologie et Sécurité des Aliments, 33883 Villenave d’Ornon, France
e-mail: [email protected]
R. Courtecuisse
Département de botanique,
Faculté des sciences pharmaceutiques et biologiques, 59006 Lille, France
K. D. Hyde
Botany and Microbiology Department, College of Science, King Saud University,
Riyadh 11442, Saudi Arabia
A. K. Guelly
Département de Botanique, Faculté des Sciences, Université de Lomé, B.P. 1515,
Lomé, Togo
D. E. Desjardin
American and African+Asian species along with temperate species. In this study, we used approximately 50 distinct species from a small area of northern Thailand, most probably being novel species. This diversity indicates that Agaricusis a species-rich genus in the tropics as well as in temperate regions. The number of species and the hypo-thetical paleotropical origin of the genus are discussed.
Keywords Agaricus. Basidiomycota . Tropical biodiversity . Biogeography . ITS . Phylogeny
Introduction
Agaricus L. is a genus of basidiomycetes that includes numerous species of which about 200–250 are presently
known. Previous estimates of species are 300–400
world-wide, but the number is probably closer to 400 according to Bas (1991). Species live in various climates on all continents, with the exception of Antarctica. Agaricus species are saprobic and generally humicolous. Several species are collected (A. campestris L. : Fr.) or cultivated (A. bisporus (J.E. Lange) Imbach, the button mushroom) for consumption or for medicinal use (A. subrufescens Peck; Angeli et al.2006; Bernarshaw et al.2007). Despite their ecological and economical interest, the diversity of species remains poorly known, particularly in subtropical and tropical areas. Numerous species of Agaricus have however, been described from tropical areas over the last century (Baker and Dale1951; Berkeley and Broome1871; Heinemann1956a,b,c,1957,1961,1962a,1962b,1962c,
1971, 1978,1980,1982, 1990,1993; Murrill 1918,1942,
1945, 1946; Pegler 1966, 1968, 1969, 1977, 1983, 1986; Pegler and Rayner 1969; Peterson et al. 2000; and Rick
1906, 1919, 1920, 1930, 1939, 1961). Agaricus species have a limited number of characteristic phenotypic traits, and therefore identification of species can be challenging due to environmental effects and intraspecific variability; it is therefore often difficult to correctly identify specimens in the field.
In temperate areas and particularly in Europe and North America, this situation is changing because of recent progress in the classification facilitated by molecular characterization and phylogeny. The genus Agaricus has been shown to be monophyletic (Vellinga et al. 2011). Among the eight sections recognized in the subgenus Agaricus (Parra 2008), Bivelares (Kauffman) L.A. Parra and Xanthodermatei Singer have been phylogenetically reconstructed (Challen et al. 2003; Kerrigan et al. 2006; Kerrigan et al. 2008) and others are under investigation. Circumscription of the species and sections has been improved, but phylogenetic reconstruction for groups of temperate species is far from complete. For example,
section SanguinolentiJul. Schäff. & Møller ex L.A. Parra seems polyphyletic and the circumscription of species as frequently encountered as A. campestris remains obscure. Species identification, as well as description of new species, should take characteristic traits into consideration, not only of the species but also of the phylogenetic group to which they belong. However, different traits can be selected to characterize the groups. For instance, the yellow vs. red discolouration of the flesh of the sporophore that was considered a major trait 50 years ago, although still important, should not be heavily weighted, since two non-related sections, Arvenses Konrad & Maubl. and Xanthodermatei, share this trait (Parra2008). In contrast, certain odours appear to be synapomorphic and are crucial taxonomic traits, at least for temperate species (Parra
2008).
Despite the number of species described, mainly between 1956–1993 by P. Heinemann and D.N. Pegler
from tropical and subtropical areas, many species remain poorly known, and many remain to be described judging from the specimens we have collected and sequenced from these areas within our project of the European Distributed Institute of Taxonomy (EDIT). Similarly, tropical speci-mens are often difficult to identify or describe as novel species for two main reasons: (i) literature is scattered and type specimens are not easily available for comparison; and (ii) description and classification of tropical species, despite the introduction of the tropical subgenera Conioagaricus Heinem. andLanagaricusHeinem. and the tropical section Brunneopicti Heinem & Gooss.-Font. (Heinemann1956a), has been globally based on traditional systematics of temperate species, and therefore on the traits characterizing the temperate sections. Sequence data for less than ten identified tropical species are currently available in Gen-Bank; therefore in this study we mostly use sequences from our collections in the analyses.
The understanding of temperate sections has conceptu-ally changed in recent years (Parra 2008), however the changes may neither be appropriate or sufficient to incorporate tropical diversity. The objective of the present study was to assess whether tropical Agaricus species are distributed in the same clades as the temperate species of the genus and to establish if there are any exclusively tropical clades. Our approach was to use a limited number of temperate species assigned to the different sections and a large sample of tropical species. We used tropical in the broad sense to include “tropical and subtropical”samples:
most samples however were collected in tropical regions, but some were collected at high altitudes in Mexico, or outside the“tropics”on the western coast of the USA or in
have incorporated some named species as well as some type specimens of tropical taxa.
The phylogeny is inferred via the analysis of the DNA sequences of ITS1 and ITS2 (internal transcribed spacers 1 and 2) separating the rRNA genes and that have already been largely used for the temperate species of this genus. This study will facilitate future taxonomic work on tropical Agaricusspecies and allow evolutionary considerations and biogeographical analyses.
Materials and methods
Fungal materialAmong 178 sequenced samples listed in Table 1, 156 have been collected and sequenced by the authors or are from their Herbaria. MATA816 provided by Gérardo Mata is an exception. The remaining sequences (22) were provided by Richard W. Kerrigan (SP307818, RWK2019, JH1) or were downloaded from GenBank.
The 178 sequenced specimens included seven collec-tions belonging to genera closely related to Agaricus (Heinemannomyces, Hymenagaricus, or unidentified), 38 temperate species ofAgaricus, and 133 tropical collections of Agaricus from Africa, Asia, and the Americas. The seven collections of genera closely related toAgaricushave been used in preliminary analyses with species of other genera such asHymenagaricus,Micropsalliotaand Lepiota. Since they consistently formed a sister clade or one of the most closely related clades to the monophyletic genus Agaricus, they were selected as outgroups for the analyses. ZRL43, has a sequence similar to ecv3586 which Vellinga et al. (2011) identified as Heinemannomyces splendidissimus Watling and also found to be closely related toAgaricus. The 38 temperate species were chosen among about 100 sequenced species to represent all the major phylogenetic clades or taxonomic subgroups of the eight presently recognized sections of subgenus Agaricus (Challen et al.
2003; Kerrigan et al.2006; Kerrigan et al.2008; Parra2008; and unpublished data). The type species, or in section ChitonioidesRomagn. one of its most closely related species, was included for each section:A. bitorquis(Quèl.) Sacc. in section Bivelares, A. gennadii Chatin & Boud. in section Chitonioides,A. xanthodermusGenev. in section Xanthoder-matei, A. sylvaticus Schaeff. in section Sanguinolenti, A. campestris in section Agaricus, A. litoralis (Wakef. & A. Pearson) Pilát in sectionSpissicaules(Heinem.) Kerrigan,A. arvensisSchaeff. in sectionArvenses, andA. comtulusFr. in section Minores Fr.. Four temperate samples (WC913, CA279, CA486 and CA684) were not formally identi-fied and could represent new species, however, they were assigned to sections based on their morphology. The remaining 133 tropical collections ofAgaricuswere
of the study was to clarify the classification of the tropical species and to establish to what extent the temperate Agaricus classification is useful for accommodating tropical species.
DNA isolation Three DNA isolation methods were used depending on the laboratory. At the Institut National de la Recherche Agronomique (INRA), DNA was isolated following a CTAB protocol (Saghai-Maroof et al. 1984; Doyle and Doyle 1987) with ethanol precipitation and modified as follows: approximately 25 mg of dried mushroom were ground to a fine powder in liquid nitrogen. The samples were transferred into 2 mL reaction tubes. 700 μL of hot extraction buffer (CTAB 2% w/v; NaCl 1.4 M; Tris pH 8.0100 mM; EDTA 10 mM; ß-mercaptoe-thanol 2% v/v) were added. After 20 minutes of incubation at 56°C, cell debris, polysaccharides and proteins were separated from aqueous DNA portions through two purification steps with 700 μL chloroform:isoamylalcohol (24:1). DNA was washed with 700μL precipitation buffer (CTAB 1% w/v; Tris pH 8.0 50 mM; EDTA 10 mM). The pellet was resuspended in 500 μL NaCl 1 M. DNA was precipitated with the addition of two volumes of absolute ethanol. The DNA pellet was washed 3 times in 1 mL 70% ethanol, air-dried and resuspended in 50μL sterile H2O.
At the National Botanic Garden of Belgium (BR), a similar protocol was used except that only ca. 10 mg of tissue was ground with a Retsch 300 mill; only 0.2% ß-mercaptoethanol was added to the lysis buffer; samples were lysed for 1 hour at 60°C; proteins and polysaccharides were removed by two consecutive extractions with chloro-form:isoamylalcohol (24:1), after which DNA was imme-diately precipitated by the addition of 0.8 volume isopropanol to the aqueous phase; the pellet was washed once in 600μL 70% ethanol, air-dried, and resuspended in 100μL TE pH 8.0; RNA was then digested with RNase A. DNA was isolated mostly from dry specimens, but in a few cases from CTAB-preserved tissue samples.
At San Francisco State University (SFSU) and Hong Kong University (HKU), genomic DNA of the samples collected in 2004–2007 were isolated from dried fungal
specimens using the E.Z.N.A. Forensic DNA Extraction Kit (Omega Bio-Tek, Norcross, GA, U.S.A.).
Table 1 Collections of Agaricus and closely related genera
N°a Sample Identificationb Country Genbank Collectorc Date Locationd Habitat Herbariume
COLLECTIONS FROM TEMPERATE AREAS (grouped by section) SectionBivelares
28 RWK1462 A. bitorquis USA AF432898 From GenBank
29 MATA681 A. tlaxcalensisT Mexico EU363033 From GenBank SectionChitonioides
30 PEAR83340 A. pearsonii France JF797186 AM 11/1983 Perpignan LIP
31 CA684 A.sp. Australia JF797187 PW 22/12/2008 Canberra, Capital Territory Lawn,Acacia CGAB
32 ARP173 A. bernardi USA AF432880 From GenBank
33 CA387 A. gennadii France JF797188 ORo 2005 Seine-Maritime, Tancarville On roadside CGAB SectionXanthodermatei
34 CA217 A. phaeolepidotus France DQ185552 From GenBank
35 RWK1938 A. hondensis USA DQ182513 From GenBank
36 CA186 A. freirei France DQ185553 From GenBank
40 CA160 A. xanthodermulusT France AY899273 From GenBank
48 CA15 A. xanthodermus France AY899271 From GenBank
SectionSanguinolenti
49 WC913 (A. fuscofibrillosus) USA AY484684 From GenBank
50 LAPAG341 A. sylvaticus Spain JF797178 LP 06/11/2004 Madrid, Parque del Retiro UnderCedrus LAPAG
51 LAPAG283 A. benesii Spain JF797179 LP 13/11/2003 Burgos, Barrio de Cortes UnderPinus LAPAG 52 LAPAG531 A. bohusii Czech Repub. JF797180 OJ 15/09/2002 Tremosnice near Caslav UnderCarpinus LAPAG
55 RWK1415 A. pattersonae USA AY943974 From GenBank
56 CA123 A. boisseletii France DQ182531 From GenBank SectionAgaricus
57 CA87 A. cupreobrunneus France DQ182532 From GenBank
58 RWK1917 A. campestris USA AF432877 From GenBank
59 LAPAG141 A. langei Spain JF797181 LP 27/10/1999 Gumiel de Mercado Grass, oak LAPAG SectionSpissicaules
72 CA486 A.sp. France JF797189 JG, AG 23/09/2006 Gironde, Illats UnderRobinia CGAB
74 CA406 A. lanipes France JF797190 anonymous 13/10/2004 Paris, Salon MNHN CGAB
76 CA279 A.sp. France JF797191 TB 27/08/2004 Dordogne Brouchau Thenon Forest edge CGAB
78 CA829 A. litoralisf France JF727867 JG, AG 09/10/2010 Gironde, Préchac In grassland CGAB 79 RWK1940 A. subrutilescens USA AY943973 From GenBank
80 CA583 A. aff.impudicus France JF797192 JG 13/10/2007 Gironde, Léognan In grassland CGAB
88 CA177 A. bresadolanus France DQ185570 From GenBank SectionArvenses
94 CA590 A. augustus France JF797193 PC,JG 28/05/2008 Gironde, Labrède UnderCedrus CGAB
Fungal
97 WC777 A. fissuratus Denmark AY484683 From GenBank
99 ECVel2339 A. inapertus USA AF482834 From GenBank
98 CA640 A. arvensis France JF797194 JG 16/10/2008 Gironde, Villenave d’Ornon UnderPinus CGAB SectionMinores
103 CA101 A. aridicola France JF797195 JG 05/11/1997 Oléron Island In dune CGAB 118 Horak68/79 A. viridopurpurascensT New Zealand JF514525 EH 12/02/1968 Kowai, Monts Grey Nothofagus BR 121 LAPAG77 A. pseudolutosus Spain JF727868 LP 06/11/1997 Honrubia de la Cuesta In meadow LAPAG
122 CA490 A. brunneolusg France JF797203 JG 30/09/2006 Gironde, Blanquefort Glade of oak CGAB
123 LAPAG339 A. comtulus France JF715065 LP-RK-PC 15/10/2003 Yvelines, Mantes la Jolie In lawn LAPAG 125 GAL9420 A. campbellensisT New Zealand DQ232644 From GenBank
124 LAPAG111 A. heinemannianus Spain JF797182 LP 19/11/1999 Segovia, Pradales UnderPinus LAPAG Collections from tropical or subtropical areas(sorted by number of species; species1to4belong to genera closely related toAgaricus)
1 ZRL3043 cf.He. splendidissimus Thailand JF691559 ZR 10/06/2006 Chiang Mai, Mae Taeng In forest BBH 1 ZRL3062 cf.He. splendidissimus Thailand DD 13/06/2006 Chiang Mai, Mae Taeng In forest BBH 2 LAPAF9 Hy. ardosiicolor Togo JF727840 LP 21/05/2010 Lomé, University Campus Azadirachta LAPAF
2 LAPAF14 Hy. ardosiicolor Togo LP 21/05/2010 Lomé, University Campus Azadirachta LAPAF
3 CA833 Thailand JF727858 JG-GB 25/07/2010 Chiang Mai, University P In forest 4 CA801 Thailand JF727859 JG-GB 25/07/2010 Chiang Mai, University P In grassland
4 ZRL3103 Thailand ZR 15/09/2006 Chiang Rai, Ob Luang P In forest BBH
5 Goossens5066 A. heterocystisT RDCongo JF514522 MG 11/1948 Panzi, Kivu Grazed bush BR
5 NTF9 Thailand NT 25/06/2010 Chiang Mai, Mae Taeng In forest
6 ZRL10.072 China JF514543 ZR 25/07/2010 Yunnan, Xishuangbana In forest 7 CA819 Thailand JF727860 JG-GB-SK 27/07/2010 Chiang Mai, University P In grassland
8 CA799 Thailand JF727863 JG-GB 24/07/2010 Chiang Mai, University P In forest
9 CA820 Thailand JF727861 JG-GB-SK 27/07/2010 Chiang Mai, University P In grassland
10 ADK4732 A. subsaharianusT Burkina-Faso JF440300 EM 25/07/2004 Ouagadougou In urban park BR 11 NTT117 Thailand JF514534 SK-KW 27/07/2010 Chiang Mai, Mae Taeng In forest
12 CA800 Thailand JF727862 JG-GB 25/07/2010 Chiang Mai, University P In grassland
13 NTS116 Thailand JF514532 SK-JG 27/07/2010 Chiang Mai, University P In grassland 13 NTS115 Thailand SK-JG 27/07/2010 Chiang Mai, University P In grassland 14 NT019 Thailand JF727844 SK-KW 22/0720/09 Chiang Rai, Khun Kone W In forest
15 NTS113 Thailand JF514531 SK-JG 27/07/2010 Chiang Mai, University P In grassland
16 ADK2564 (A. brunneopictus) Bénin JF514518 AK 08/06/1999 Niaouli, Plateau Old termite hill BR 17 ZRL3031 Thailand JF691550 DD 07/06/2006 Chiang Mai, DSPNP In forest BBH
17 ZRL3064 Thailand TO 13/06/2006 Chiang Mai, DSPNP In forest BBH
Table 1 (continued)
N°a Sample Identificationb Country Genbank Collectorc Date Locationd Habitat Herbariume
18 NTT34 Thailand JF514536 SK 19/06/2010 Chiang Mai, Mae Taeng In forest
19 LAPAF1 A. inoxydabilis Togo JF727841 LP 12/05/2010 Ola In corn field LAPAF
20 ZRL4017 Thailand JF691549 PS 15/05/2007 Chiang Mai, Mae Taeng In forest BBH 20 NTT118 Thailand SK-KW-JG 27/07/2010 Chiang Mai, University P In grassland
20 ZRL3005 Thailand ZR 26/05/2006 Chiang Mai, Mae Taeng In forest BBH
21 CA856 Thailand JF797202 JG 25/07/2010 Chiang Mai, University P In grassland
22 ZRL2043 Thailand JF691553 JK 26/06/2005 Chiang Mai, Mae Taeng In forest BBH
22 ZRL3086 Thailand ZR 22/07/2006 Chiang Mai, Chiang Dao In forest BBH 22 ZRL2085 Thailand TB 03/07/2005 Chiang Mai, Mae Taeng In forest BBH
23 LAPAF2 A. campestroides Togo JF727842 LP 12/05/2010 Ola-Okpa-Fou Path in forest LAPAF
24 ZRL3099 Thailand JF691556 ZR 05/09/2006 Chiang Mai, Mae Taeng In forest BBH
25 ZRL2132 Thailand JF691558 ZR 21/08/2005 Chiang Mai, Mae Taeng In forest BBH 26 LAPAF4 A. trisulphuratus Togo JF727843 LP 12/05/2010 Ola-Okpa-Fou Path in forest LAPAF
27 ZRL2123 A.aff.trisulphuratus Thailand JF691557 KH 11/08/2005 Chiang Mai, Mae Taeng In forest BBH
27 NTS118 A.aff.trisulphuratus Thailand SK-JG 27/07/2010 Chiang Mai, University P In grassland
27 ZRL2128 A.aff.trisulphuratus Thailand EG 18/08/2005 Chiang Mai, Mae Taeng In forest BBH 27 ZRL3014 A.aff.trisulphuratus Thailand ZR 03/06/2006 Chiang Mai, Mae Taeng In forest BBH
37 Grinling70109 A. microvolvatulusT Congo Braz. JF514524 KG 15/01/1967 Brazzaville In forest edge BR
37 NTT84 Thailand KW 07/10/2010 Chiang Mai, Doi Suthep In forest
37 NTT38 Thailand KW 20/06/2010 Chiang Mai, Doi Suthep In forest 37 NTS117 Thailand SK-JG 27/07/2010 Chiang Mai, University P In grassland
38 ZRL3044 Thailand JF691555 TO 10/06/2006 Chiang Mai, Mae Taeng In forest BBH
39 NTF61 Thailand JF514528 NT 16/07/2010 Chiang Mai, Kiewtubyoung In forest
41 F2530 (A. caribaeus) Fr. Martinique JF727856 JF 15/04/2002 Tartane, pointe rouge In forest LIP 42 F2715 Fr. Martinique JF727847 JF 06/11/2002 Tartane, pointe rouge In forest LIP
43 ZRL3095 A.aff.endoxanthus Thailand JF691554 ZR 13/08/2006 Chiang Mai, Mae Taeng In forest BBH
44 Goossens5415 A. xanthosarcusT RDCongo JF514523 MG 12/1954 Panzi, Kivu Coffee plantation BR
44 Hendrickx515 RDCongo FH 15/04/1939 Mulungu BR
44 NTT50 Thailand KW 24/06/2010 Chiang Mai, New waterfall In forest
45 NTS7 Thailand JF514533 SK 11/05/2010 Chiang Rai, Khun Kone W In forest
45 ZRL3094 Thailand ZR 09/08/2006 Chiang Mai, Mae Taeng In forest BBH
45 NTT95 Thailand SK 17/07/2010 Chiang Rai, Doi Tung In forest 45 NT007 Thailand SK 15/08/2009 Chiang Rai, Khun Kone W In forest
46 F2767 A.aff.volvatulus Fr. Martinique JF727848 JF 19/06/2003 Grand rivière In forest LIP
46 ADK2785 A.aff.volvatulus Bénin AK 13/06/2000 Niaouli, Plateau In forest BR
47 NTF58 Thailand JF514527 NT 10/07/2010 Chiang Mai, DSPNP In forest
Fungal
53 ZRL3012 Thailand JF691551 ZR 03/06/2006 Chiang Mai, Mae Taeng In forest BBH
54 ZRL2136 Thailand JF691552 TBa 07/06/2005 Chiang Mai, DSPNP In forest BBH
54 ZRL2109 Thailand ZR 02/08/2005 Chiang Mai, Mae Taeng In forest BBH 60 F2047 A.aff.argyropotamicus Fr. Martinique JF727849 JF 28/06/1999 Fort-de-France In lawn LIP
61 F2272 Fr. Martinique JF727850 JF 12/07/2000 Lamentin LIP
61 LD026 Thailand JC 25/07/2010 Chiang Rai, MFL University In grassland
61 F3109 (A.aff.argyropotamicus) Fr. Martinique JF LIP
62 NTS05 Thailand JF514530 SK 29/04/2010 Chiang Rai, Muang In forest
63 F2389 (A. magnivelaris) Fr. Martinique JF727851 JF 02/06/2001 Gros-Morne, Rivière Rouge In forest LIP
64 F2187 Fr. Martinique JF727852 JF LIP
65 NT020 Thailand JF797197 SK 21/08/2009 Chiang Mai, Mae Taeng In forest
66 MATA816 Mexico JF727870 GM 07/2010 Veracruz, Coatepec XAL
67 F1779 (A. johnstonii) Fr. Martinique JF727853 JF 06/07/1998 Prêcheur, Anse Couleuvre In forest LIP
68 JH1 A.cf.floridanus USA JF896226 JH 07/2006 Suffolk Co., New York Mulch SFSU
69 LAPAM1 A.aff.rufoaurantiacus Venezuela JF797183 LP 01/04/2006 Choroni LAPAM
70 CL/GUAD05.099 A.aff.rufoaurantiacus Fr. Guadeloupe JF727857 CLu 29/11/2005 Sainte-Rose, Trace de Sofaïa LIP 71 CJL090302-05 Fr. Guiana JF727869 JLC 02/03/2009 Cayenne, La Mirande
73 ADK2171 A.cf.goossensiae Benin JF514517 AK 20/06/1998 Wari Maro, Borgou On soil BR
75 F2255 (A. parasilvaticus) Fr. Martinique JF797198 JF 02/04/2000 Pointe banane In forest LIP
75 F2039 Fr. Martinique JF 27/06/1999 Saint-Esprit, Bois la Charles In forest LIP 77 F2301 Fr. Martinique JF727854 JF 10/10/2000 Case-Pilote, Morne Rose LIP
81 RC/GUY07.019 Fr. Guiana JF797199 CL 27/02/2007 Sinnamary, Parcelle Guyaflux Litter, humus LIP
82 NYS122 (A.cfaugustus) Benin JF514540 SY 11/07/1999 Wari Maro, Borgou BR
83 NTT42 Thailand JF514538 KW 20/06/2010 Chiang Mai, Doi Suthep In forest 83 ZRL10.071 China ZR 25/07/2010 Yunnan, Xishuangbannan In forest
84 LAPAF3 A.aff.impudicus Togo JF797184 LP 12/05/2010 Ola In wood litter LAPAF
85 ZRL3093 Thailand JF691548 OM 08/06/2006 Chiang Mai, Mae Taeng In forest BBH
86 Rammeloo5756 A. kivuensis Burundi JF514541 JR 17/11/1978 Mugara Oil palm plantation BR 86 Goossens5323 A. kivuensisT RDCongo MG 11/1953 Panzi, Kivu Grevillea BR
86 Goossens5406 A. kivuensisT RDCongo MG 12/1954 Panzi, Kivu Coffee plantation BR
87 F2467 (A. porosporus) Fr. Martinique JF797200 JF 26/12/2001 Trinité, La Caravelle In forest LIP
89 LAPAM4 Brazil JF797185 JB 17/04/2005 Rio de Janeiro In forest edge LAPAM 90 DeMeijer1904 (A. argyropotamicus) Brazil JF797196 AMe 02/04/1991 Paraná, Paranagua In dune BR
91 RWK2019 A. deserticola USA JF896228 JS 07/2007 Bernalillo Co., New Mexico Sandy soil SFSU
92 F2285 A. fiardiih Fr. Martinique JF797201 JF 26/09/2000 Sainte-Anne In forest LIP
Table 1 (continued)
N°a Sample Identificationb Country Genbank Collectorc Date Locationd Habitat Herbariume
92 F2286L A. fiardiih Fr. Martinique JF 29/09/2000 Saint-Anne, Morne Manioc In forest LIP
93 NTF67 A.sp.h Thailand JF514529 SK 17/08/2010 Chiang Rai, Muang In forest
93 ZRL2036 A.sp.h Thailand RL 25/06/2005 Chiang Mai, Mok Fa W In forest BBH 93 NT001 A.sp.h Thailand SK 150/8/2009 Chiang Rai, Khun Kone W In forest
93 ZRL2134 A.sp.h Thailand KH 28/08/2005 Chiang Mai, Mae Taeng In forest BBH
95 Thoen7297 Senegal JF514542 DT 14/10/1984 Dakar In market BR
96 ZRL2127 Thailand JF691547 KH 16/08/2005 Chiang Mai, Mae Taeng In forest BBH
96 OR71 Thailand OR 03/07/2010 Chiang Mai, Mae Taeng In forest
96 ZRL3028 Thailand ZR 05/06/2006 Chiang Mai, Doi Inthanon In forest BBH
96 CA798 Thailand JG-SK 28/07/2010 Chiang Mai, Nat.Park In forest
96 ZRL2053 Thailand DD 27/06/2005 Chiang Mai, Doi Inthanon In forest BBH
100 F2815 A. martinicensis Fr. Martinique JF727855 JF-RC-CLu 02/09/2003 Prêcheur, Anse Couleuvre In forest LIP 100 CL/MART03.055 A. martinicensis Fr. Martinique CL 02/09/2003 Prêcheur, Anse Couleuvre In forest LIP
100 F2343 A. martinicensis Fr. Martinique JF 19/11/2000 Prêcheur, Anse Lévrier In forest LIP
101 SP307818 A. martinezianus Brazil JF896227 UP 14/12/2001 Sao Paulo On soil
102 NTT37 Thailand JF514537 KW 20/06/2010 Chiang Mai, Mae Taeng In forest
102 ZRL2110 Thailand ZR 03/08/2005 Chiang Mai, Mae Taeng In forest BBH
102 ZRL3039 Thailand TBa 08/06/2006 Chiang Mai, Mae Taeng In forest BBH
104 CA848 Thailand JF727864 JG-GB 25/07/2010 Chiang Mai, University P In forest
105 ZRL3088 Thailand JF691543 ZR 02/08/2006 Chiang Rai, Pamae Lao P In forest BBH
105 NTSCR1 Thailand SK 08/03/2010 Chiang Rai, Muang In forest
106 NTT33 Thailand JF514535 KW 18/06/2010 Chiang Mai, DSPNP In forest
106 NTS73 Thailand SK 30/06/2010 Chiang Mai, Mae Taeng In forest
107 NTF26 Thailand JF514526 NT 29/06/2010 Chiang Mai, Doi Suthep In forest 108 CA846 Thailand JF727865 JG-GB 25/07/2010 Chiang Mai, University P In forest
109 ZRL2044 Thailand JF691540 ZR 26/06/2005 Chiang Mai, Mae Taeng In forest BBH
109 ecv3614 Thailand HM488742 EV From GenBank
110 NT055 Thailand JF727846 SK 15/08/2009 Chiang Mai, DSPNP In forest
111 ADK2905 (A.cfbulbillosus) Benin JF514520 AK 19/09/2000 Wari Maro, Borgou In sandy soil BR
111 CA847 Thailand JG-GB 25/07/2010 Chiang Mai, University P In forest
112 ZRL3101 Thailand JF691544 ZR 13/09/2006 Chiang Mai, Mae Taeng In forest BBH
113 NT62 Thailand JF727845 SK 20/08/2009 Chiang Mai, Mae Taeng In forest 114 NTT72 Thailand JF514539 SK 07/03/2010 Chiang Mai, Mae Taeng In forest
115 CA843 Thailand JF727866 JG-GB 04/08/2010 Hua-Hin, Golf In grassland
116 ZRL3080 Thailand JF691542 ZR 10/07/2006 Chiang Mai, Mae Taeng In forest BBH
116 NTF063 Thailand NT 28/07/2010 Chiang Mai, Mae Taeng In forest
Fungal
117 ZRL3091 Thailand JF691546 ZR 02/08/2006 Chiang Rai, Pamae Lao P In forest BBH
119 MATA774 Mexico JF727871 GM-PC 29/11/2007 Veracruz, San Andrés Tuxtla In dune XAL
120 ADK2751 A. goossensiae Benin JF514519 AK 05/06/2000 Calavi Campus, Atlantique In sandy soil BR 126 ZRL3056 Thailand JF691541 ZR 12/06/2006 Chiang Mai, Mae Taeng In forest BBH 127 ZRL3102 Thailand JF691545 ZR 15/09/2006 Chiang Mai, Ob Luang P In forest BBH
128 LD030 Thailand JF514521 JC 03/08/2010 Chiang Rai, MFL University On litter MFLU
128 ZRL2124 Thailand KH 12/08/2005 Chiang Mai, Mae Taeng In forest BBH
128 NTS106 Thailand SK 24/07/2010 Chiang Mai, Mae Taeng In forest
a Number of species (as numbered in the phylogenetic tree); when there are redundant samples belonging the same presumed species, the first one appearing in the list was used in the phylogenetic analyses and the genbank accession number of its ITS1+2 sequence is given in the Table
b T, Type (holotype, paratype or isotype); the species names in brackets require complementary studies of the collection to be confirmed or not.
c AG, A. Guinberteau; AK, A. De Kesel; AM, A. Marchand; AMe, A. De Meijer; CL, C. Lechat; CLu, C. Lecuru; DD, D. Desjardin; DT, D. Thoen; EH, E. Horak; EG, E. Grand; EM, E. Maes; EV, E. C. Vellinga; FG, K. Grinling; FH, F. Hendrickx; GB, G. Barroso; GM, G. Mata; JB, J. Borovicka; JC, J. Chen; JF, J.P. Fiard; JG, J. Guinberteau; JH, J. Horman; JK, J. Kerekes; JLC, J.-L. Cheype; JR, J. Rammeloo; JS, J. W. Sparks II; KH, K. D. Hyde; KW, K. Wisitrassameewong; LP, L. Parra; MG, M. Gossens-Fontana; NT, N. Thongklong; OJ, O. Juhasz; OM, O. Myo Aung; OR, O. Raspé; ORo, O. Roblot; PC, P. Callac; PS, P. Sysouphanthong; PW, P. Wenzel; R. Courtecuisse; RK, R.W. Kerrigan; SK, S. Karunarathna; SY, S. Yorou Norou; TB, T. Boonpratuang; TBa, T. Baroni; TBo, T. Bouchara; UP, U.C. Peixoto; ZR, R.L. Zhao;
d P, Park or National Park; W, Waterfall; MNHN, Museum National d
’Histoire Naturelle; DSPNP, Doi Suthep Pui National Park.
e SFSU: Harry D. Thiers Herbarium, Department of Biology, San Francisco State University. USA; BBH: Biotec Bandkok Herbarium, National Science and Technology Development Agency, Klong Luang, Pathumthani, Thailand; LIP: Herbier du Département de Botanique, Faculté des Sciences Pharmaceutiques et Biologiques, Université de Lille, Lille, France; BR : National Botanic Garden of Belgium Herbarium, Meise, Belgium; CGAB:Collection du germoplasme des agarics à Bordeaux, INRA, Bordeaux, France; XAL : Herbario del Instituto de Ecología, A.C., Xalapa, Veracruz, Mexico; LAPAM, LAPAF, LAPAG: Luis Alberto Parra private herbarium for America, Africa and Europe respectively, Aranda de Duero, Burgos, Spain. MFLU: Mae Fah Luang University, Chiang Rai Prov., Thailand.
f syn
. A. spissicaulis
g syn.
A. porphyrizon
h
At INRA, the reaction mix (final volume 25 μL) contained 5 μL PCR GO Taq buffer (5X, Promega), 2.5 μL dNTP mix (1.2 mM, Eurobio), 0.5 μL BSA (10 mg/mL, Promega); 1 μl of each primer (25 μM); 0.2μL Taq polymerase (5 U/μL, Go Taq Promega); 1μL DNA extract; ddH2O up to 25 μL. The PCR profile was
5 min at 95°C; 35 cycles (1 min at 94°C, 1.5 min at 55°C, 1.5 min at 72°C); 5 min at 72°C. At BR, amplifications were usually performed in 50μL reactions using Dream-Taq DNA polymerase (Fermentas), according to manu-facturer’s instructions, except that reactions contained
0.2 mg/mL BSA (Fermentas) and 0.25μM of each primer. Cycling conditions were as follows: 3 min at 95°C; 35 cycles of 1 min at 94°C, 1.5 min at 55°C, 1.5 min at 72°C; 5 min at 72°C. At SFSU and HKU, the thermal cycles consisted of 3 min at 94°C, 30–35 cycles of 1 min at 94°C,
50 sec at 52°C and 1 min at 72°C, with a final extension step of 72°C for 10 min.
Sequencing was performed on ABI Prism Genetic analysers (Applied Biosystems) at the following institu-tions: Beckman Coulter Genomics, England; Macrogen, Republic of Korea; Genome Research Centre of the University of Hong Kong; Department of Biology, San Francisco State University.
Phylogenetic analysesBased on our previous knowledge on the circumscription of species in Agaricus, sequences were considered as redundant in the three following cases: they were identical; they differed only by heteromorphisms and shared presumed alleles; they differed at a single polymorphic position and possibly by heteromorphisms at others. The entities defined above must be considered as presumed species because such a method might in some cases be too stringent for taxonomic purposes.
After alignment using T-Coffee ver 8.99 (Notredame et al.2000), corrections were made by hand, firstly because T-Coffee did not interpret the heteromorphisms, secondly to suppress highly variable or ambiguous positions.
The maximum likelihood (ML) analysis was performed on the ATGC bioinformatics platform. The phylogenetic tree was constructed using the ML method implemented in the PhyML ver 3.0 aLRT (Guindon and Gascuel2003; Anisimova and Gascuel2006). This method does not allow partitions. The GTR substitution model was selected with an estimated proportion of invariable sites of 0.438 and assuming 4 gamma-distributed rate categories to account for rate heterogeneity across sites. The gamma shape parameter was estimated directly from the data (gamma= 0.782), and the likelihood was increased by using the SPR tree improvement. Reliability of internal branches was assessed using the approximate Likelihood-Ratio test (aLRT) which assesses that the branch being studied provides a significant likelihood gain, in comparison with
the null hypothesis that involves collapsing that branch but leaving the rest of the tree topology identical (Anisimova and Gascuel 2006). The analysis included 100 bootstrap replicates. We also performed a Bayesian analysis with MrBayes 3.1 (Huelsenbeck and Ronquist 2001; Ronquist and Huelsenbeck 2003). We first estimated the best evolutionary model for ITS1 and ITS2 separately with jModeltest (Posada 2008), using the Bayesian information criterion, and restricting the explored evolutionary models to the ones implemented in MrBayes. The best models were GTR +Γ+ I and HKY +Γ+ I, respectively. We partitioned the data and set the evolutionary models accordingly in MrBayes. The model parameters were unlinked across partitions. We ran two parallel analyses, each with one cold and three heated chains, with tree sampling every 200th generation. Since the two runs converged slowly (the average deviation of split frequen-cies decreased slowly), we ran the analyses for 10×106
generations, and when summarizing the tree samples the first 12,000 trees were discarded. The burnin was determined by inspecting the log-likelihood by generation plot generated with Tracer ver 1.5 (Rambaut and Drummond
2007). Graphical representation and editing of the phyloge-netic tree were performed with TreeDyn ver.198.3 (Chevenet et al.2006).
Results
Sampling for analyses using ITS1+2 sequences
We obtained complete sequence data from the 178 samples presented in Table 1. Ten of these were from types specimens, 51 tropical samples were collected in 2010, mainly in Thailand but also in China, Togo and Mexico; the oldest herbarium collection was from 1939. Among the 178 samples, 89 were from Asia, 37 from the Americas, 28 from Europe, 21 from Africa, and 3 from Oceania.
first, pairwise sequence comparisons indicated that 60 putative species from the tropics were found once, 15 two times, 10 three times, and 5 four times or more. Second, for six presumed species of tropical Agaricus, redundant sequences from different continents were found: four in Africa and Asia (5:A. heterocystisHeinem. & Gooss.-Font.; 37: A. microvolvatulus Heinem.; 44: A. xanthosarcus Heinem. & Gooss.-Font.; and 111), one in Africa and America/Caribbean (46: A. aff. volvatulus Heinem. & Gooss.-Font.), and one in Asia and America/Caribbean (61). The 80 remaining species of Agaricuswere found on a single continent: 47 in Asia, 22 in the Americas, and 11 in Africa; some of them have nearly but not entirely redundant sequences and belong to presumed closely related species, e. g.26and 27(A. trisulphuratus Berk. group), or92and 93
clarify their taxonomic status.
Phylogenetic results
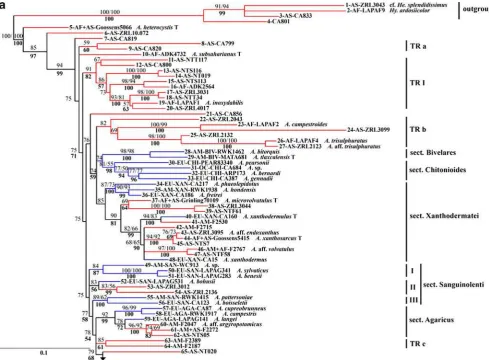
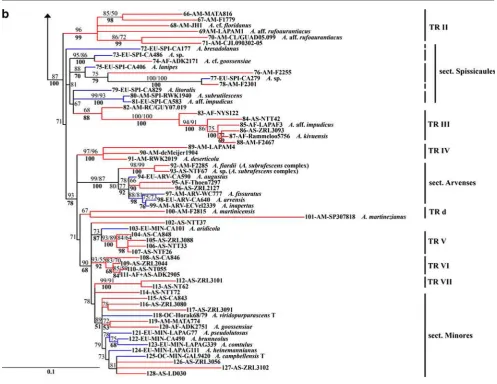
The most likely ML tree obtained by analysis is presented in Fig. 1. The branches having SH-like support values lower than 50% are collapsed. SH-like support values and Bayesian posterior probabilities of the 50% majority rule consensus tree are showed. Bootstrap support values were mostly lower than 50%; those above 50% are showed.
Most of the major clades were similar and relatively well supported in the ML and Bayesian analyses with SH-like support values and posterior probabilities greater than 80%, while the bootstrap support values were some-times much lower. Despite numerous polytomies and
Fig. 1 Most likely ML phylogram based on ITS1+2 sequences of 124 species ofAgaricus. The SH-like branch support values above 50%, the bootstrap support values above 50% (after slash) and the posterior probabilities of 50% majority rule consensus Bayesian tree (in bold type) are shown. Red and blue branches refer to tropical and temperate species ofAgaricus,respectively. Branch label indications
are: the number of the species (outgroups from 1 to 4; Agaricus
species from 5 to 128), the continent (Africa AF, the Americas AM, Asia AS, Europe EU, and Oceania OC), the section for the temperate species only (Bivelares BIV,Chitonioides CHI,SanguinolentiSAN, AgaricusAGA,SpissicaulesSPI,ArvensesARV,MinoresMIN), and
topological differences, the ML and Bayesian trees remain congruent enough to have most of their major clades globally appearing in the same order.
Because deep branches, except the branch bearing species 66 to 128, are very short, we preferred to emphasise the content of the clades (detailed in Table2) rather than phylogenetic relationships between clades. One consequence of these short branches is the low bootstrap support values that we obtained, while aLRT SH-like values are less sensitive to the shortness of the branches.
Phylogenetic distribution of the temperate species
Temperate species, except A. aridicola Geml, Geiser & Royse (103), were distributed into eight accepted taxonom-ic sections of subgenusAgaricus.
The sections Bivelares,Chitonioides,Xanthodermatei, SanguinolentiandAgaricusare related. In both trees, they are monophyletic, except the sectionSanguinolenti which is paraphyletic and constituted three clades. Among the
seven clades of these five sections, four are entirely temperate and the three remaining ones have an early temperate branch.
The section Spissicaules which is a less well-known section of the subgenus, is constituted by three clades and two branches that form a large polytomy with clade TR II and clade TR III in the Bayesian tree. In the ML tree, this section is paraphyletic with clade TR III. The possible polyphyly of section Spissicaules and the phylogenetic relationships with the closely related clades TR II and TR III remain unresolved.
The sectionsArvensesandMinoreswith the exception of the secotioid species A. aridicola are monophyletic in the Bayesian tree and in the ML tree.
Twelve temperate species that have not been included in previous phylogenetic analyses were also taxonomically distributed in the eight sections. Temperate species, except A. aridicola and two species of the section Spissicaules which is not clearly resolved, are distributed in 12 clades (Table 2). All of these clades have SH-like support values greater than 80%, eight of them have Bayesian posterior
Fig. 1 (continued)
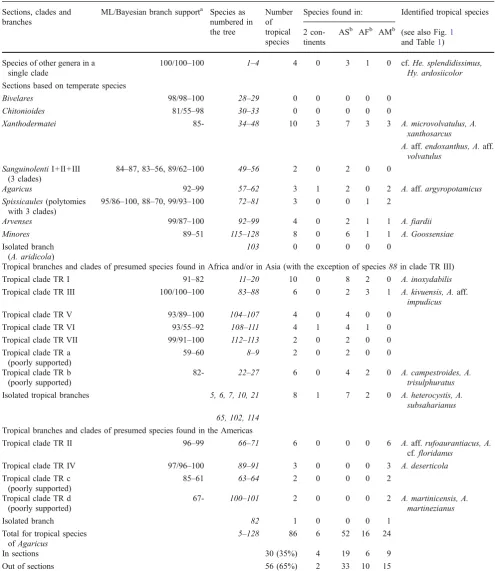
Sections, clades and branches
ML/Bayesian branch supporta Species as numbered in the tree
Number of tropical species
Species found in: Identified tropical species
2 con-tinents
ASb AFb AMb (see also Fig.1 and Table1)
Species of other genera in a single clade
100/100–100 1–4 4 0 3 1 0 cf.He. splendidissimus,
Hy. ardosiicolor
Sections based on temperate species
Bivelares 98/98–100 28–29 0 0 0 0 0
Chitonioides 81/55–98 30–33 0 0 0 0 0
Xanthodermatei 85- 34–48 10 3 7 3 3 A. microvolvatulus, A.
xanthosarcus
A.aff.endoxanthus, A.aff.
volvatulus SanguinolentiI+II+III
(3 clades)
84–87, 83–56, 89/62–100 49–56 2 0 2 0 0
Agaricus 92–99 57–62 3 1 2 0 2 A.aff.argyropotamicus
Spissicaules(polytomies with 3 clades)
95/86–100, 88–70, 99/93–100 72–81 3 0 0 1 2
Arvenses 99/87–100 92–99 4 0 2 1 1 A. fiardii
Minores 89–51 115–128 8 0 6 1 1 A. Goossensiae
Isolated branch (A. aridicola)
103 0 0 0 0 0
Tropical branches and clades of presumed species found in Africa and/or in Asia (with the exception of species88in clade TR III)
Tropical clade TR I 91–82 11–20 10 0 8 2 0 A. inoxydabilis
Tropical clade TR III 100/100–100 83–88 6 0 2 3 1 A. kivuensis, A.aff.
impudicus
Tropical clade TR V 93/89–100 104–107 4 0 4 0 0
Tropical clade TR VI 93/55–92 108–111 4 1 4 1 0
Tropical clade TR VII 99/91–100 112–113 2 0 2 0 0
Tropical clade TR a (poorly supported)
59–60 8–9 2 0 2 0 0
Tropical clade TR b (poorly supported)
82- 22–27 6 0 4 2 0 A. campestroides, A.
trisulphuratus
Isolated tropical branches 5, 6, 7, 10, 21 8 1 7 2 0 A. heterocystis, A.
subsaharianus 65, 102, 114
Tropical branches and clades of presumed species found in the Americas
Tropical clade TR II 96–99 66–71 6 0 0 0 6 A.aff.rufoaurantiacus, A.
cf. floridanus
Tropical clade TR IV 97/96–100 89–91 3 0 0 0 3 A. deserticola
Tropical clade TR c (poorly supported)
85–61 63–64 2 0 0 0 2
Tropical clade TR d (poorly supported)
67- 100–101 2 0 0 0 2 A. martinicensis, A.
martinezianus
Isolated branch 82 1 0 0 0 1
Total for tropical species ofAgaricus
5–128 86 6 52 16 24
In sections 30 (35%) 4 19 6 9
Out of sections 56 (65%) 2 33 10 15
a SH-like branch support values above 50%, bootstrap support values above 50% (after a slash), and posterior probabilities of 50% majority rule consensus Bayesian tree (after a dash)
probabilities greater than 80%, and four of the eight also have bootstrap support values greater than 80%.
Phylogenetic and geographic distributions of the tropical species
The four non-Agaricus species representing outgroups, formed a clade sister toAgaricusin both trees; although it was not necessary to impose them as outgroups in the ML analysis. This clade was supported at 100% by all the methods. These outgroups are from tropical Africa or Asia. Thirty five percent of the tropical species of Agaricus (30 species) were distributed amongst six of the eight accepted sections of the subgenus Agaricus (Table 2). In other terms, about one-third of tropical (including subtrop-ical) Agaricus species nested in seven of the 12 clades based on temperate species. The 56 remaining tropical species were distributed in eleven exclusively tropical clades and in nine isolated branches detailed in Table 2. Among the eleven tropical clades detected in the analyses, seven that are well supported by both analyses are numbered as TR I to TR VII. Both SH-like branch support and Bayesian posterior probability were greater than 95% for four of these seven clades, greater than 90% for two, and greater than 80% for the remaining clade. Moreover, the four clades III, IV, V, and VII have high bootstrap support values of 100, 96, 89, and 91 respectively.
Geographic information appears highly correlated with the phylogenetic data although African, Asian and Amer-ican samples were not equally represented. Firstly, all basally joined species including those of the closely related genera, from1to20, are tropical and were found in Africa and/or in Asia, but not in the Americas. Secondly, among the 11 tropical clades, three contained eight species collected in Asia, three contained 20 species found only in Africa and/or Asia, four contained 13 species found only in the Americas, and one contained six species from the three continents with a single species from the Americas (87from Martinique). Samples from Africa were underrep-resented, thus exclusively African clades possibly exist. Likewise, it is possible that the exclusively Asian clades would be in fact African and Asian. In Table 2, tropical clades and isolated branches of Africa and Asia are separated from those of the Americas.
The separation between the American and the African+ Asian tropical species is much less pronounced in the sections based on temperate species than in the tropical clades although the tropical species are 50% less numerous in the former than in the latter. Five sections contain both American and African+Asian tropical species: the four monophyletic sectionsXanthodermatei, Agaricus, Arvenses, Minores, and the possibly polyphyletic sectionSpissicaules. Section Sanguinolenti contains tropical species from Asia
only and the two remaining sections Bivelares and Chitonioideshave no tropical species. Moreover, the only two species for which samples were found in the Americas and in Africa+Asia (46 and 61) belong to clades of the monophyletic sectionsXanthodermateiandAgaricus.
Discussion
The number of recognized Agaricus species lies between 200 and 250 according to Bas (1991) and is estimated as ca 200 mostly temperate species in Kirk et al. (2008). These estimations are lower than our own calculation of 386 recognized species among which 203 are temperate and 183 are tropical. This estimation is updated with the most recently described species but remains approximate because some synonymies have not been detected between species described on different continents or climates. Thus, the numbers of tropical and temperate recognized species are quite similar. In our analyses, we used sequences from 86 tropical species and 38 temperate species carefully selected among about 100 species to represent their distribution in the eight presently recognized temperate sections of subgenus Agaricus (Challen et al. 2003; Kerrigan et al.
2006; Kerrigan et al. 2008; Parra 2008). Tropical and temperate samples are approximately equally represented. Concerning the number of species in existence, Bas (1991) recognized 200-250 described species in Agaricus, but estimated there were 300–400 species worldwide. We
believe this is also an underestimate since it is similar to the number of presently recognized species and since probably most of the species collected in Thailand in the present study are new. The total number of species collected in only a small part of northern Thailand during the last five years, including the about 50 used in the present study, is likely to be more than the 70 species recognized in Europe over one century. From the present study, we have also established that a large part of the American samples are taxonomically different from those of Africa and Asia. The number of tropical species is certainly much greater than the number of temperate species and the total number of species in existence should be therefore much greater than 400.
In the genus Agaricus ITS sequences were used for species characterization or for phylogenetic analyses of the sections of the genus; however, at the scale of the entire genus, it appears that most of the deep branches are short suggesting that major evolutionary radiations would have occurred in a relatively short time. In other respects, the low bootstrap support values of the deep branches are partly due to their shortness. The present analysis was not aimed at establishing a very strong phylogeny for the genus, but identifies the major clades and the distribution of the tropical species among these clades.
temperate species into eight sections with the inclusion of tropical species plus an additional dozen temperate species never included in previous phylogenetic analyses (except for the secotioid species A. aridicola). The nLSU rDNA sequence of our collection CA101 of A. aridicola has previously been obtained by Moncalvo et al. (2002) and also used by Geml et al. (2004) and Capelari et al. (2006) in analyses. This species, exhibiting a positive Schaeffer reaction (orange, red or violaceous-purple discoloration when aniline and nitric acid are consecutively applied on the same spot of a sporophore) and an odor of almond, was primarily assigned to sectionMinoresaccording to the data of Geml et al. (2004). However, tropical samples were mostly absent in this study and our present data better agree with the study of Capelari et al. (2006) who found that A. aridicolawas more closely related toA. martinezianusthan to sectionsArvensesand Minores. With caution, we do not include A. aridicola in the clade TR V which would be much less supported with this species. In other respect, the
‘temperate’status of this secotioid species is doubtful since
it is rare in France but abundant in Israel according to Wasser (2002). To classify the species in climatic groups is not easy: the geographical range of some temperate species such as A. bisporus and A. bitorquis extends into tropical areas and, reciprocally, the tropical speciesA. subrufescens exists also in Europe (Kerrigan 2005). Moreover, some species such as the tropical speciesA. endoxanthusBerk. & Broome are sometimes found in greenhouses (Parra et al.
2002) in Europe and are suspected to have been introduced with plants.
Our analyses are similar to those of Geml et al. (2004) and show that the three secotioid species,A. aridicola,A. deserticola G. Moreno, Esqueda & Lizárraga, and A. inapertus Vellinga, do not share a common ancestor. The secotioid form, considered as an adaptation to arid environment, has therefore evolved independently several times in a large clade that includes sections Arvenses and Minores, and five related tropical clades (samples from89 to128).
Two-thirds (56/86) of the tropical or subtropical species did not cluster in the recognized sections based on temperate species. Nine species occurred on isolated branches, 12 form poorly supported clades, TR a to TR d, and 35 belong to seven well-supported clades, I to VII. We accept these seven tropical clades of which four are more strongly supported in the analyses (III, IV, V, and VII). Characterization of these tropical clades will be challeng-ing. There are at least four examples where tropical species share some characteristics with the recognized temperate sections, although they do not belong to these sections. (i) Two samples both having affinity withA. impudicus(Rea) Pilát, a species of section Spissicaules, belong to two
section Spissicaules as expected, but the tropical species (85) to the most closely related tropical clade TR III. (ii) The sample of A. subsaharianus L.A. Parra, Hama & De Kesel (10) has traits that, until now characterized only sections Spissicaules, Minores and Arvenses (Hama et al.
and their circumscription need consideration. Only one-third of tropical Agaricus species are distributed in the recognized infrageneric organization of the genus based on temperate species. Although some species among the remaining two-thirds could be included in the tropical sections proposed by Heinemann, it appears necessary to expand the number of sections in the genus.
The fact that numerous clades are exclusively tropical indicates that geography and climate have had a major impact on the evolution of the genus. Adaptation to temperate climates has never occurred in certain genera of Agaricaceaesuch as Micropsalliota(Zhao et al. 2010). A relative tolerance to cold is required in temperate climates and the ability to fruit at ambient temperatures that is required under warm climates is not advantageous in temperate regions because the summer is not the wettest season and consequently the most favorable season for fruiting. Largeteau et al. (2011) showed that the rate of wild isolates able to fruit at 25°C is 100% inA. bisporus var. burnettii Kerrigan & Callac, a variety that lives in hot climates, while it is only about 50% in the populations ofA. bisporusvar.bisporusliving in temperate climates.
Our data also indicate that clades of four of the eight recognized temperate sections include both American and African+Asian tropical species. To our knowledge, all temperate sections contain North American and Eurasian temperate species, and the distribution range of numerous temperate species also extends to both continents (Challen et al. 2003; Kerrigan et al. 2006; Kerrigan et al. 2008). These species probably migrated via land before the complete separation of these continents. Moreover, for some species, such as A. bisporus and A. bitorquis, subpopulations that adapted to (sub)tropical climates in Africa or in North America are known. For example, the geographic range of A. bisporus extends from the boreal region of Alaska (Geml et al. 2008) to the equatorial climate of Congo (Heinemann1956a) in Africa and to the hot and dry climate of the Sonoran Desert of California in America (Callac et al.1993); the ancestral condition of such species, however, remains unknown. Finally, a similar hypothesis for the amphiatlantic distribution of the tempe-rate species and for the American and African+Asian distribution of the tropical species belonging to the same clades as the temperate species needs consideration.
Forty-seven tropical species are distributed in 11 tropical clades. Seven of these clades contain species exclusively from Africa+Asia with the exception of one species in clade III, while the four remaining clades contain species exclusively from the Americas. The different phylogenetic distribution of the American species versus the African+Asian species appears to be a reliable pattern since four tropical clades contained only species found in America although the Asian samples were overrepresented compared to the American
samples. The distinct African+Asian and American tropical clades also suggest that species diversification probably occurred independently in Africa+Asia and in the Americas. Moreover, some clades belonging to the African+Asian group are not phylogenetically closely related, indicating that species diversification occurred several times on these continents. This is also possible in the Americas but less evident from the data. One of the four species used in the outgroup and that does not belong inAgaricus, was identified as Hymenagar-icus ardosiicolor (Heinem.) Heinem. while another shares some characteristic traits withHeinemannomyces splendid-issimusbut others withHymenagaricusandXanthagaricus. However, when other GenBank sequences of Hymenagar-icusspecies were included in the analyses (data not shown), they diverged more from the Agaricusclade than the four species that we used, with the exception ofHymenagaricus epipastus (Berk. & Broome) Heinem. & Little Flower recently reported from Thailand by Zhao et al. (2010). Further studies will be necessary to clarify the taxonomic rank of these taxa. In Vellinga et al. (2011) one of the two species Heinemannomyces splendisissimus or Clarkeinda trachodes(Berk.) Singer is sister to the monophyletic genus Agaricus depending on the analysis. Hymenagaricus epipastus, Hymenagaricus ardosiicolor, Heinemannomyces splendisissimus, Clarkeinda trachodesand two unidentified species in our outgroups are from tropical Africa or Asia.
The geographical origin of Agaricus is unknown, however, the following evidences suggest a paleotropic origin. (i) Species of genera known to be the most closely related toAgaricusspecies are from tropical Africa or Asia. (ii) All the early branches and clades of our trees (from species 1 to 20) contain species exclusively from tropical Africa or Asia; and (iii) the first African+Asian clades appeared before the first American ones. This hypothesis remains to be confirmed because these basal branches and clades are not strongly supported in the analyses (Fig. 1). The analyses should also be extended to include species from tropical Australia and temperate areas of Asia and the South Hemisphere to better understand the biogeography of the genus and possibly correlate migrations with the known periods of connection between continents.
Acknowledgments The authors are grateful to Jean-Pierre Fiard,
Else Vellinga, Gerardo Mata, Marina Capelari, Marc-André Lachance, Rick Kerrigan, Peter Wenzel, Dario De Franceschi, Vincent Lefort, Stephane Welti and Saturnino (Nino) Santamaría. This work was supported by an Integration Research Grant from the European Distributed Institute of Taxonomy (EDIT).
The National Science Foundation (USA) (PEET-grant DEB-0118776 to Desjardin), the National Natural Science Foundation of China (Project ID: 31000013), and the project“value added products from Basidiomycetes: Putting Thailand’s biodiversity to use” (BRN049/2553) are thanked for providing partial support to this research. The Global Research Network for Fungal Biology and King Saud University are also thanked for support.
Angeli JPF, Ribeiro R, Gonzaga MLC, Soares S de A, Ricardo MPSN, Tsuboy MS, Stidl R, Knasmueller S, Linhares RE, Mantovani MS (2006) Protective effects ofβ-glucan extracted from Agaricus brasiliensis against chemically induced DNA
damage in human lymphocytes. Cell Biol Toxicol 22:285–291 Anisimova M, Gascuel O (2006) Approximate likelihood ratio test for
branches: A fast, accurate and powerful alternative. Syst Biol 55:539–52
Bas C (1991) A short introduction to the ecology, taxonomy and nomenclature of the genusAgaricus. In: Van Grievsen LJLD (ed) Genetics and breeding of Agaricus. Proceedings of the First International Seminar on Mushroom Science. Pudoe Wageningen pp 21–24
Baker RED, Dale WT (1951) Fungi of Trinidad & and Tobago. Mycol Papers 33:1–123
Berkeley MJ, Broome CE (1871) The fingi of Ceylon (Hymenomy-cetes from Agaricus to Cantharellus. J Linn Soc 9:494–567 Bernarshaw S, Lyberg T, Hetland G, Johnson E (2007) Effect of an
extract of the mushroomAgaricus blazeiMurill on expression of
adhesion molecular and production of reactive oxygen species in monocytes and granulocytes in human whole blood ec vivo Acta Pathologica. Microbiol Immunol 115(6):719–725
Callac P, Billette C, Imbernon M, Kerrigan RW (1993) Morpholog-ical, genetic, and interfertility analyses reveal a novel, tetrasporic variety of Agaricus bisporus from the Sonoran desert of California. Mycologia 85:835–851
Capelari M, Rosa LH, Lachance M-A (2006) Description and affinities of Agaricus martineziensis, a rare species. Fungal Diversity 21:11–18
Challen MP, Kerrigan RW, Callac P (2003) A phylogenetic recon-struction and emendation of Agaricus sectionDuploannulatae. Mycologia 95(1):61–73
Chevenet F, Brun C, Banuls AL, Jacq B, Christen R (2006) TreeDyn: towards dynamic graphics and annotations for analyses of trees BMC. Bioinformatics 7:439
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15 Geml J, Geiser DM, Royse DJ (2004) Molecular evolution of
Agaricus species based on IST and LSU rDNA sequences.
Mycol Prog 3(2):157–176
Geml J, Laursen GA, Taylor DL (2008) Molecular diversity assessment of artic and boreal Agaricus taxa. Mycologia 100 (4):577–589
Guindon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52:696–704
Hama O, Maes E, Guissou M-L, Ibrahim DM, Barage M, Parra LA, Raspé O, De Kesel A (2010) Agaricus subsaharianus, une nouvelle espèce comestible et consommée au Niger, au Burkina Faso et en Tanzanie. Cryptogamie, Mycologie 31:221– 234
Heinemann P (1956a) Champignons récoltés au Congo Belge par Mme M Goossens-Fontana, II Agaricus Fr ss. Bull Jard Bot État 26:1–127
Heinemann P (1956b) Flore Iconographique des Champignons du Congo, 5º fascicule:AgaricusI. Ministère de l’Agriculture-Jardin Botanique de l’État Bruxelles
Heinemann P (1956c) Champignons récoltés au Congo Belge par Mme M Goossens-Fontana, II Agaricus, Note complémentaire. Bull Jard Bot État 26:325–333
Heinemann P (1957) Flore Iconographique des Champignons du Congo, 6º fascicule: Agaricus II et Pilosace. Ministère de l’Agriculture-Jardin Botanique de l’État Bruxelles
Heinemann P (1962a) Agarici Austro-Americani II Agaricus de
Bolivie. Bull Jard Bot État 32:1–21
Heinemann P (1962b) Agarici Austro-Americani III TroisAgaricusde
la Jamaïque. Bull Jard Bot État 32:23–28
Heinemann P (1962c) Agarici Austro-Americani IV QuatreAgaricus
de Venezuela. Bull Jard Bot État 32:155–161
Heinemann P (1971) QuelquesPsalliotesdu Congo-Brazzaville. Cah
Maboké 9:5–10
Heinemann P (1978) Essai d’une clé de determination des genres
AgaricusetMicropsalliota. Sydowia 30:6–37
Heinemann P (1980) Les genres Agaricus et Micropsalliota en Malaisie et en Indonésie. Bull Jard Bot Belg 50:3–68
Heinemann P (1982) Quelques Psalliotes de Nouvelle Guinée (Papua New Guinea). Bull Jard Bot Belg 52:405–413
Heinemann P (1984) Agarici Austro-Americani VII. Agariceae des zones tempérées de l’Argentine et du Chili. Bull Jard Bot Belg 60:331–370
Heinemann P (1988) Novitates generisMicropsalliotae(Agaricaceae). Bull Jard Bot Belg 60:540–543
Heinemann P (1990)Agaricus singaporensis sp. nov. Bull Jard Bot
Belg 60:417–419
Heinemann P (1993) Agarici Austro-Americani VIII Agaricaceae des régions intertropicales d’Amérique du Sud. Bull Jard Bot Belg 62:355–384
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogeny. Bioinformatics 17:754–755
Kerrigan RW (2005) Agaricus subrufescens, a cultivated edible and medicinal mushroom, and its synonyms. Mycologia 97(1):12–24 Kerrigan RW (2007) Lectotypification of Agaricus brunnescens.
Mycologia 99(6):906–915
Kerrigan RW, Callac P, Challen M, Guinberteau J, Parra LA (2006 “2005”)AgaricussectionXanthodermatei: a phylogenetic recon-struction with commentary on taxa. Mycologia 97(6):1292–1315 Kerrigan RW, Callac P, Parra LA (2008) New and rare taxa in
Agaricus section Bivelares (Duploannulati). Mycologia 100 (6):876–892
Kirk PM, Cannon PF, Minter DW, Stalpers JA (2008) Dictionary of the fungi, Tenthth edn. CABI publishing, Wallingford
Largeteau ML, Callac P, Navarro-Rodriguez A-M, Savoie J-M (2011) Diversity in the ability ofAgaricus bisporuswild isolates to fruit
at high temperature (25°C). Fungal Biology, doi:10.1016/j. funbio.2011.08.004
Moncalvo JM, Vilgalys R, Redhead SA, Johnson JE, James TY, Aime MC, Hofstetter V, Verduin SJW, Larsson E, Baroni TJ, Thorn RG, Jacobsson S, Clémencon H, Miller OK Jr (2002) One hundred seventeen clades of Euagarics. Mol Phylogen Evo l23:357–400
Murrill WA (1918) Agaricaceae of tropical North America VIII. Mycologia 10:62–85
Murrill WA (1942) New fungi from Florida. Lloydia 5(2):136–157 Murrill WA (1945) New Florida fungi. Quart J Florida Acad Sci 8
(2):175–198
Murrill WA (1946) New and interesting Florida fungi. Lloydia 9 (4):315–330
Notredame C, Higgins DG, Heringa J (2000) T-Coffee: a novel method for multiple sequence alignments. J Mol Biol 302:205–217 Parra LA (2008) Agaricus L Allopsalliota Nauta & Bas. Pars 1
Edizioni Candusso Alassio, Italy
Parra LA, Villarreal M, Esteve-Raventos F (2002) Agaricus endox-anthusuna specie tropicale trovata in Spagna. Rivista Micol 45 (3):225–233
Pegler DN (1966) Tropical African Agaricales. Persoonia 4(2):73–124 Pegler DN (1968) Studies on African Agaricales I. Kew Bull 21:499–
Pegler DN (1969) Studies on African Agaricales II. Kew Bull 23:219– 249
Pegler DN (1977) A preliminary Agaric flora of east Africa. Kew Bull Add Series 6:1–615
Pegler DN (1983) Agaric flora of the Lesser Antilles. Kew Bull Add Series 9:1–667
Pegler DN (1986) Agaric flora of Sri Lanka. Kew Bull Add Series 12:1–519
Pegler DN, Rayner RW (1969) A contribution to the Agaric flora of Kenya. Kew Bull 23:347–412
Peterson KR, Desjardin ED, Hemmes DE (2000) Agaricales of the Hawaiian Islands. 6. Agaricxaceae I. Agariceae: Agaricus and Melanophyllum. Sydowia 52(2):204–257
Posada D (2008) jModelTest: phylogenetic model averaging. Mol Biol Evol 25:1253–1256
Rambaut A, Drummond AJ (2007) Tracer v1.5, Available fromhttp:// tree.bio.ed.ac.uk/software/tracer/
Rick J (1906) Pilze aus Rio Grande do Sul (Brazilien). Broteria 5:5–53 Rick J (1919) Contributio II ad monographiam Agaricinorum
Brasiliensium. Broteriaser Bot 17:101–111
Rick J (1920) Contributio ad monographiam Agaricinorum Brasilien-sium. Broteria ser Bot 18:48–63
Rick J (1930) Contributio IV ad monographiam Agaricinorum Brasiliensium. Broteria ser Bot 24:97–118
Rick J (1939) Agarici Riograndenses IV. Lilloa 4(1):75–104 Rick J (1961) Basidiomycetes Eubasidii in Rio Grande do Sul—
Brasilia 5 Agaricaceae. Iheringia ser Bot 8:296–450
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: Bayesian phylo-genetic inference under mixed models. Bioinformatics 19:1572– 1574
Saghai-Maroof MA, Solima KM, Jorgenson RA, Allard RW (1984) Ribosomal DNA spacer-length polymorphisms in barley: Men-delian inheritance, chromosomal location, and population dy-namics. Proc Natl Acad Sci USA 81:8014–8018
Vellinga EC, Sysouphanthong P, Hyde KD (2011) The family of Agaricaceae: phylogenies and two new white-spored genera. Mycologia 103(3):494–509
Wasser SP (2002) Family Agaricaceae (FR.) Cohn of Israel mycobiota I. Tribe Agariceae Pat., ed Nevo E & Volz PA, A.R.G. Gantner Verlag K.G., Ruggell, Liechtenstein
White TJ, Bruns T, Lee S, Taylor JW (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic, New York, pp 315–322
Zhao R, Desjardin DE, Soytong K, Perry BA, Hyde KD (2010) A monograph of Micropsalliota in Northern Thailand based on