Directory UMM :Data Elmu:jurnal:T:Tree Physiology:Vol16.1996:
Teks penuh
Gambar
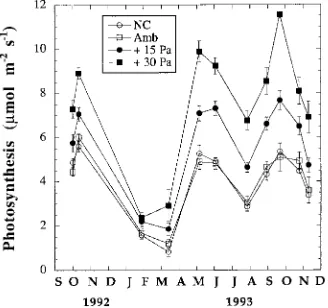
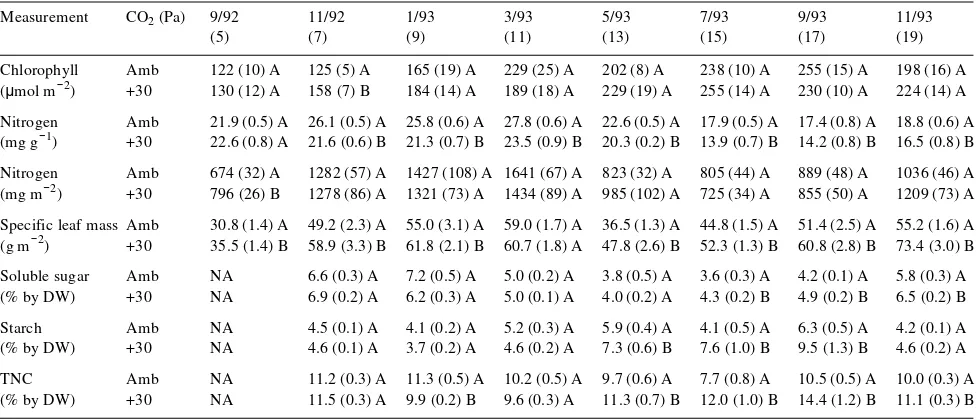
Dokumen terkait
Sesuai dengan teori yang dipaparkan oleh David (2011) terkait enam pedoman pengembangan pasar dapat menjadi sebuah strategi yang efektif yaitu PT Bintang
Batu, 2 Juli 2012 Panitia Pengadaan Barang/Jasa Dinas Cipta Karya dan Tata Ruang. Kota
Demikian berita acara penjelasan pekerjaan ini dibuat dengan sesunguhnya dan dapat dipergunakanan sebagaimana mestinya. PANITIA
Maka melalui penelitian ini peneliti berharap dapat menjadi acuan bagi perusahaan dalam menerapkan prinsip- prinsip etika bisnis yang benar sesuai dengan aturan
Pokja ULP Pengadaan Truk Armroll pada Dinas Kebersihan dan Pertamanan Kota Banjarmasin akan melaksanakan Pelelangan Sederhana dengan pascakualifikasi untuk paket
SANYOTO
[r]
Terhadap penawaran yang dinyatakan tidak memenuhi persyaratan administrasi tidak dilanjutkan ke tahap evaluasi
