HASIL DAN PEMBAHASAN
HasilAnatomi area postrema
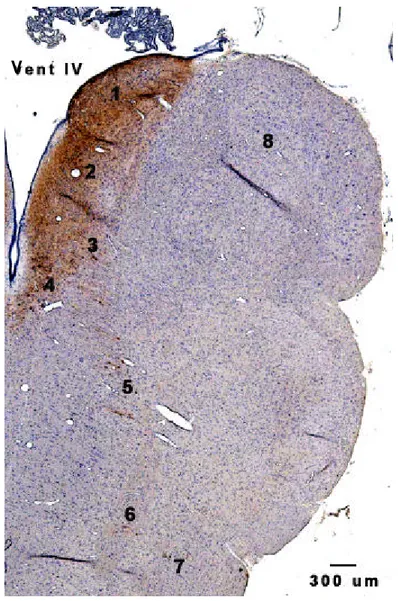
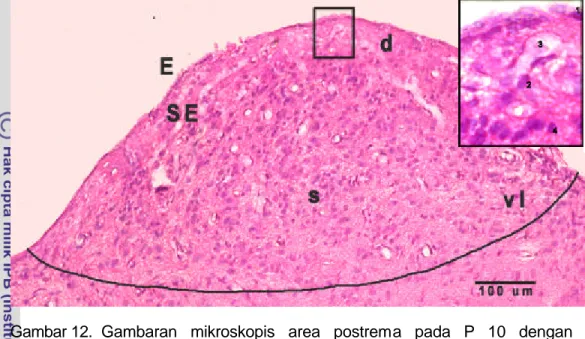
Bentuk dan posisi anatomi AP pada monyet ekor panjang secara umum diwakili oleh gambar area postrema di medula oblongata MEP pada P 105, dengan pewarnaan imunohistokimia terhadap TH (Gambar 11). Area postrema terletak pada bagian dorsal medula oblongata, berbentuk setengah bulat dan berbatasan dengan ventrikel IV. Bagian kaudal AP berbatasan dengan nukleus grasilis, sedangkan rostral dan ventral berbatasan dengan nukleus traktus solitarius. Secara histologis AP dibagi menjadi bagian dorsal, sentral dan ventrolateral. Pada bagian dorsal terdapat lapisan ependima yang berbatasan dengan ventrikel IV dan tersusun oleh sel berbentuk kubus atau pipih. Di profundal lapisan ependima terdapat lapisan subependima atau bagian internal yang tersusun oleh neuron yang secara umum berbentuk oval atau bulat, astrosit dengan inti kecil, dan kaya pembuluh darah (Gambar 12).
Perkembangan morfologi neuron KA di AP
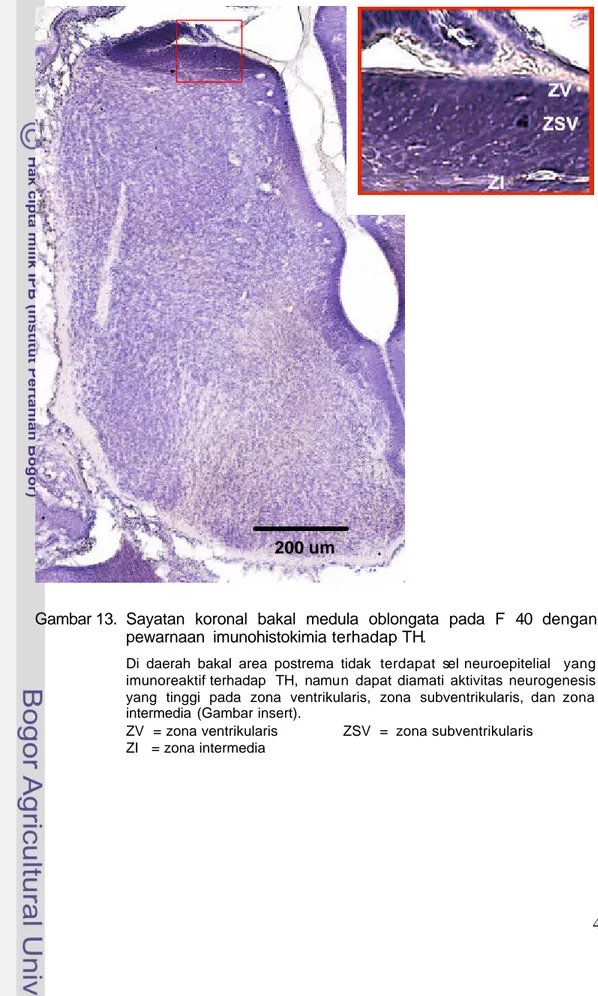
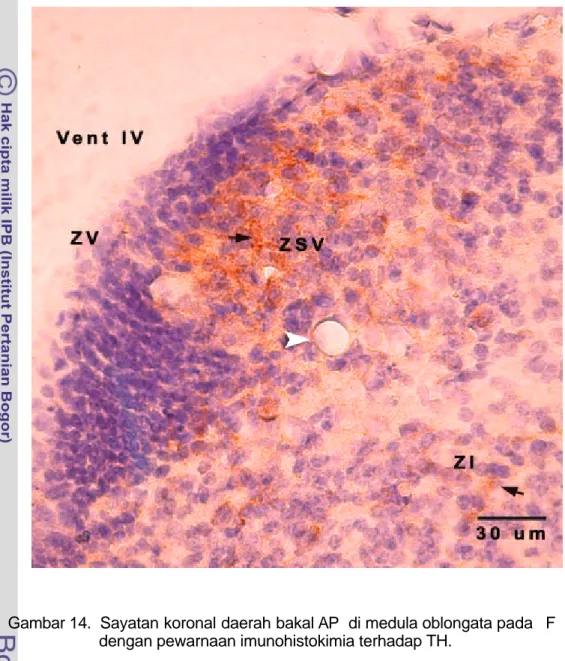
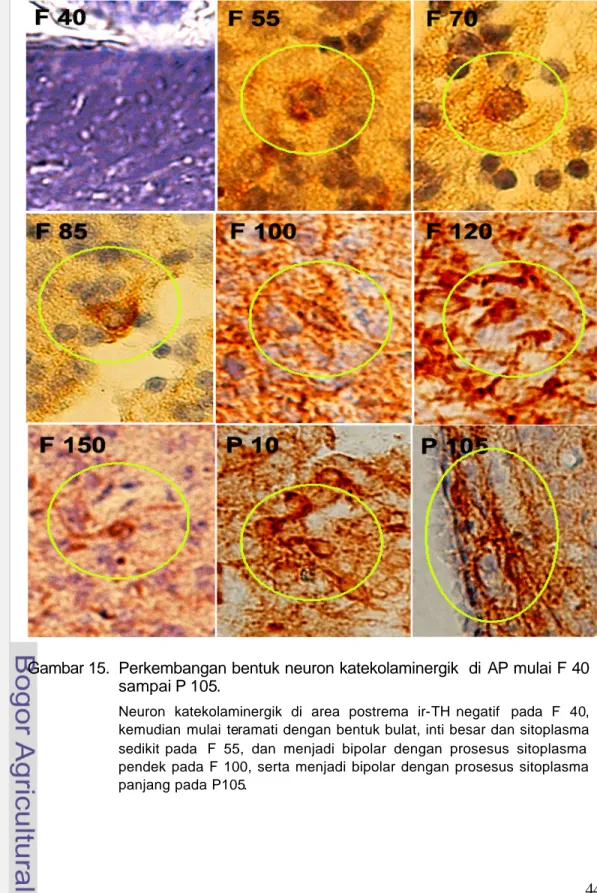
Neuron KA pada F 40, di daerah bakal medula oblongata belum ditemukan, tetapi dapat dilihat adanya aktivitas neurogenesis tinggi yang ditandai dengan tebalnya zona ventrikularis dan zona subventrikularis (Gambar 13). Neuron KA mulai ditemukan pada F 55 di daerah bakal AP. Bakal AP terletak berbatasan dengan ventrikel IV, berbentuk agak bulat dan menonjol ke arah ventrikel. Pada zona ventrikularis dan zona subventrikularis tampak sel-sel yang berproliferasi sehingga daerah ini tampak tebal dan dipenuhi sel-sel muda. Neuron KA dapat dilihat pada zona subventrikularis dan zona intermedia (Gambar 14). Neuron KA pada F 55 ini berbentuk bulat dengan inti besar dan memiliki sedikit sitoplasma (Gambar 15).
Pada F 70, neuron masih berbentuk bulat dengan prosesus sitoplasma yang sedikit dan pendek. Peningkatan sitoplasma dan prosesusnya mulai terjadi pada F 85. Stadium perkembangan menengah dimulai pada F 100, ditandai dengan neuron yang berbentuk bipolar serta prosesus sitoplasma lebih panjang.
Gambar 11. Sayatan koronal medula oblongata pada P 105 dengan perwarnaan imunohistokimia terhadap TH.
Neuron katekolaminergik (berwarna coklat kekuningan) terdistribusi pada 1 = Area postrema 2 = Nukleus traktus solitarius
3 = Nukleus motoris dorsal dari nervus vagus 4 = Nukleus hipoglosus 5 = Formasio retikularis sentralis 6 = Nukleus ambigus 7 = Nukleus retikularis lateralis 8 = Nukleus grasilis Vent IV = Ventrikel IV
Gambar 12. Gambaran mikroskopis area postrema pada P 10 dengan pewarnaan HE.
Daerah area postrema dibagi menjadi bagian dorsal, sentral, dan ventrolateral. Permukaan AP dilapisi oleh lapisan ependima yang tersusun atas sel ependima berbentuk kubus. Lapisan subependima terletak di profundal lapisan ependima dan banyak terdapat neuron, pembuluh darah, serta astrosit.
d = bagian dorsal s = bagian sentral vl = bagian ventrolateral E = lapisan ependima SE = subspendima 1 = sel ependima 2 = neuron 3 = pembuluh darah 4 = astrosit
Sampai pada P 105, neuron di AP didominasi oleh bentuk neuron bipolar dengan arah prosesus sitoplasma yang sejajar dengan permukaan AP, dan hanya sebagian kecil yang arahnya tegak lurus terhadap permukaan. Pada P 105 ini bentuk neuron bipolar tidak berubah, tetapi jumlahnya bertambah seiring dengan pertambahan umur.
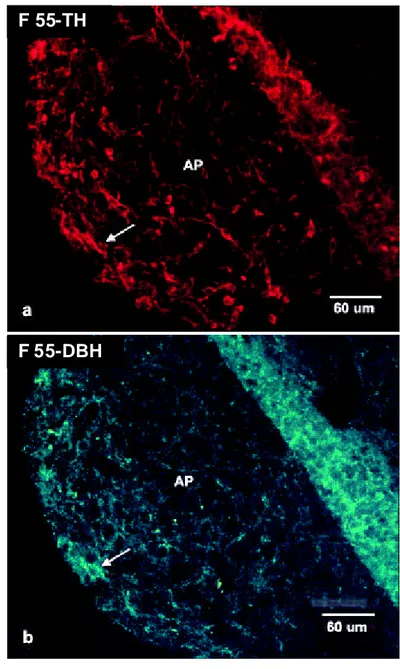
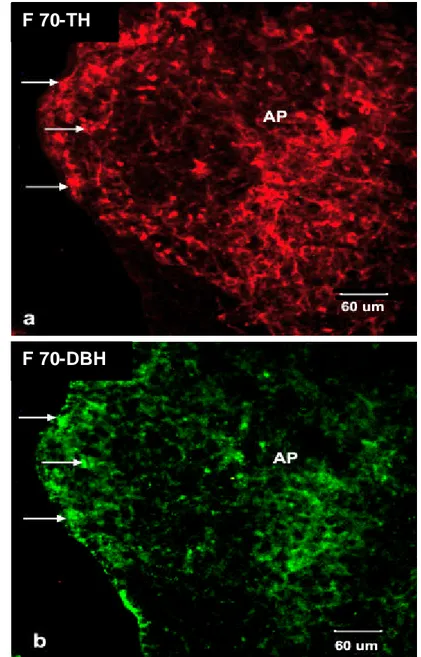
Berdasarkan pengamatan dengan pewarnaan fluoresen pada F 55 dapat diamati bahwa ukuran neuron KA berukuran kecil, kemudian bertambah besar pada F 70, dan semakin besar pada F 145 (Gambar 17).
Gambar 13. Sayatan koronal bakal medula oblongata pada F 40 dengan pewarnaan imunohistokimia terhadap TH.
Di daerah bakal area postrema tidak terdapat sel neuroepitelial yang imunoreaktif terhadap TH, namun dapat diamati aktivitas neurogenesis yang tinggi pada zona ventrikularis, zona subventrikularis, dan zona intermedia (Gambar insert).
ZV = zona ventrikularis ZSV = zona subventrikularis ZI = zona intermedia
Gambar 14. Sayatan koronal daerah bakal AP di medula oblongata pada F 55 dengan pewarnaan imunohistokimia terhadap TH.
Area postrema terletak berbatasan dengan ventrikel IV dan neuron katekolaminergik terdistribusi pada zona subventrikularis dan zona intermedia. Zona ventrikularis dipadati oleh sel yang sedang berproliferasi. Kepala panah putih menunjukkan pembuluh darah di area postrema. Neuron KA (warna coklat) dapat dilihat pada daerah SVZ dan IZ (anak panah hitam ).
ZSV = zona subventrikularis ZI = zona intermedia ZV = zona ventrikularis
Gambar 15. Perkembangan bentuk neuron katekolaminergik di AP mulai F 40 sampai P 105.
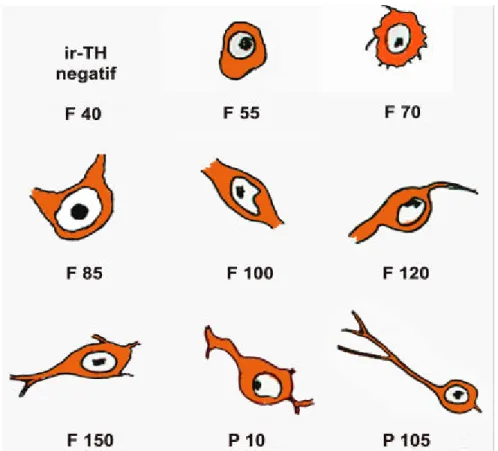
Neuron katekolaminergik di area postrema ir-TH negatif pada F 40, kemudian mulai teramati dengan bentuk bulat, inti besar dan sitoplasma sedikit pada F 55, dan menjadi bipolar dengan prosesus sitoplasma pendek pada F 100, serta menjadi bipolar dengan prosesus sitoplasma panjang pada P105.
Migrasi neuron KA pada AP dan distribusinya
Pada F 55, di bagian dorsomedial medula oblongata pada level nukleus olivaris dapat ditemukan banyak neuron yang ir-TH di zona subventrikularis dan zona intermedia, sedangkan di zona intermedia dan bagian ventrolateral hanya sedikit yang imunoreaktif. Zona ventrikularis, subventrikularis dan zona intermedia yang ir-TH ini diduga sebagai bakal AP (Gambar 18) mengingat letaknya berbatasan dengan ventrikel IV dan terdapat di bagian paling kaudal otak. Neuron KA dari daerah bakal AP kemudian mengalami migrasi mencapai bagian ventrolateral medula oblongata. Hasil ini menunjukkan bahwa pada F 55 neuron KA sudah bermigrasi mencapai daerah yang akan menjadi posisi definitif. Berdasarkan distribusi neuron KA tersebut, diduga arah perkembangan neuron KA di medula oblongata berjalan dari bagian dorsomedial menuju ke ventrolateral.
Gambar 16. Gambaran skematis perkembangan bentuk neuron KA di AP mulai F 40 sampai P105.
Gambar 17. Sayatan koronal daerah medula oblongata pada F 55 dengan pewarnaan imunohistokimia terhadap TH.
Daerah bakal AP (kotak biru), daerah tengah (kotak merah) dan ventrolateral (kotak hijau). Daerah AP terletak pada bagian dorsomedial medula oblongata dan berbatasan dengan ventrikel IV (vent IV). Tanda panah menunjukkan migrasi neuron katekolaminergik ke arah ventrolateral dari daerah bakal AP ke daerah tengah, sampai daerah ventrolateral. Insert menunjukkan adanya neuron katekolaminergik pada daerah tersebut (kepala panah).
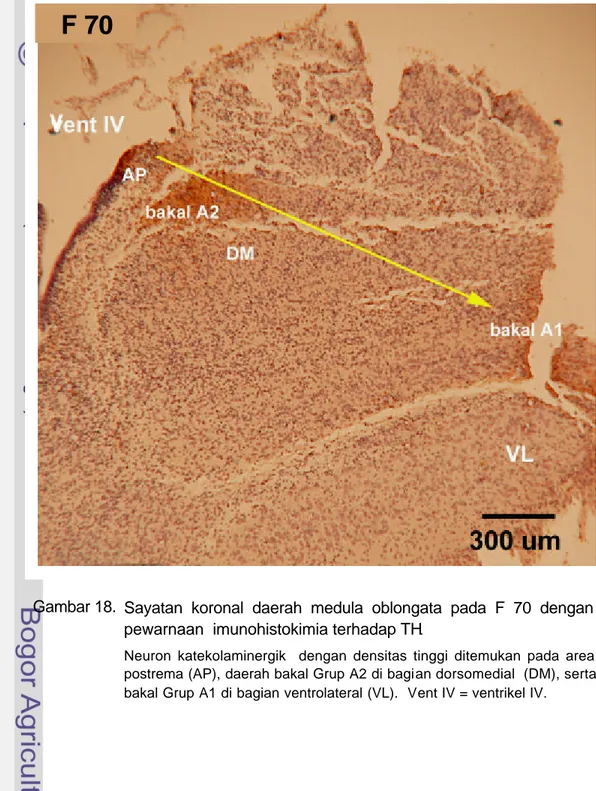
Gambar 18. Sayatan koronal daerah medula oblongata pada F 70 dengan pewarnaan imunohistokimia terhadap TH.
Neuron katekolaminergik dengan densitas tinggi ditemukan pada area postrema (AP), daerah bakal Grup A2 di bagian dorsomedial (DM), serta bakal Grup A1 di bagian ventrolateral (VL). Vent IV = ventrikel IV.
F 70
Pada F 70 dapat diamati daerah AP yang menonjol ke ventrikel IV dan mengandung neuron KA. Neuron KA di daerah dorsomedial dan ventrolateral medula oblongata densitasnya meningkat. Neuron KA selain terdistribusi di AP dijumpai juga di daerah calon nukleus traktus solitarius dan sekitarnya sebagai bakal Grup A2, serta di daerah bakal nukleus retikularis lateralis dan sekitarnya sebagai bakal Grup A1 (Gambar 19).
Pada perkembangan selanjutnya, pada F 100 neuron KA di daerah AP mulai terpisah dengan bagian ventral, yaitu kelompok neuron KA di daerah dorsomedial medula oblongata (Grup A2) yang terletak di daerah nukleus traktus solitarius, nukleus motor dorsalis nervus vagus, dan nukleus hipoglosus. Pada daerah ventrolateral medula oblongata, yaitu di nukleus retikularis lateralis dan nukleus ambigus juga terbentuk Grup A1. Gambaran ini menunjukkan bahwa di medula oblongata pada fetus MEP umur 100 hari terbentuk tiga kelompok neuron KA yaitu neuron di area postrema, neuron di dorso medial dan neuron di ventrolateral. Distribusi yang sama teramati juga pada sampel yang lebih tua sampai pada anak umur 105 hari.
Diferensiasi neuron KA di AP menjadi neuron dopaminergik atau noradrenergik
Neuron KA tampak padat pada zona subventrikuler dan zona intermedia daerah bakal AP pada F 55. Keberadaan neuron KA memperlihatkan bahwa pada tahap ini neuron KA sudah mengalami deferensiasi bersamaan dengan proses proliferasi dan migrasi dalam neurogenesis. Gambar 20, 21 dan 22 menampilkan peningkatan densitas neuron DA dan NA yang diwarnai secara imunohistokimia dengan teknik label ganda menggunakan antibodi terhadap TH dan DBH. Densitas neuron KA di AP yang bersifat ir-TH positif tetapi ir-DBH negatif lebih besar dibandingkan neuron yang ir-TH positf dan ir-DBH positif. Pada F 70, densitas neuron yang terlabel ganda (ir-TH positf dan ir-DBH positif) meningkat dibandingkan dengan F 55, demikian pula antara F 145 dengan F 70. Hasil menunjukkan bahwa pada neuron KA pada AP terutama tersusun atas neuron DA dengan sedikit neuron NA.
Gambar 19.Sayatan sagital AP pada F 55 dengan pewarnaan imunofloresen label ganda terhadap TH dan DBH.
Pada daerah AP, neuron ir-TH positif berwarna merah (a) dan neuron ir-DBH positif berwarna hijau (b). Tanda panah menunjukkan neuron yang terlabel ganda TH dan DBH.
F 55-TH
Gambar 20.Sayatan sagital AP pada F 70 dengan pewarnaan imunofloresen label ganda terhadap TH dan DBH.
Pada daerah AP, neuron ir-TH positif berwarna merah (a) dan neuron ir-DBH positif berwarna hijau (b). Tanda panah menunjukkan neuron yang terlabel ganda TH dan DBH densitasnya lebih tinggi dibandingk an dengan F 55.
F 70-TH
Gambar 21. Sayatan sagital AP pada F 145 dengan pewarnaan imunofloresen label ganda terhadap TH dan DBH.
Pada daerah AP, neuron TH positif berwarna merah (a) dan neuron ir-DBH positif berwarna hijau (b). Tanda panah menunjukkan neuron yang terlabel ganda TH dan DBH, dengan densitas yang lebih tinggi dibandingkan dengan F 55 dan F 70.
F 145-DBH
F 145-TH
b
a
Serabut ir-DBH positif tersebar sampai permukaan ventrikuler sejak F 55, sedangkan serabut ir-TH mulai menyebar sampai ke permukaan ventrikuler pada F 145. Serabut ir-DBH positif yang ditemukan pada AP memiliki densitas lebih besar dibandingkan dengan badan selnya.
Terbentuknya jalur akson katekolamin di medula oblongata
Jalur akson katekolamin di medula oblongata mulai tampak pada F 120 (Gambar 23) yang menghubungkan neuron KA Grup A1 di daerah ventrolateral dengan daerah intermedia yaitu pada formasio retikularis sentralis serta dengan Grup A2 di daerah dorsomedial. Akson dari neuron katekolaminergik bergabung membentuk jalur akson yang mengarah dorsomedial dari bagian ventromedial. Jalur akson ini disebut ventral ascending catecholamine pathway (acpv).
Intensitas pewarnaan neuron KA di AP
Pada AP bagian dorsal, baik badan sel maupun prosesus sitoplasma memiliki intensitas perwarnaan lebih kuat dibandingkan pada daerah sentral (Gambar 24). Hasil ini menunjukkan bahwa neuron pada daerah tersebut memiliki kandungan enzim TH lebih banyak atau neurotransmiter katekolamin lebih banyak pada daerah tersebut. Sebagai contoh Gambar 24 yang menampilkan area postrema F 100, terlihat jelas bahwa neuron KA di AP bagian dorsal lebih padat, terwarnai lebih kuat dengan enzim TH. Neuron pada AP berbentuk bipolar, sebagian besar sejajar dengan permukaan, namun pada bagian sentral beberapa neuron mengarah ke ventrolateral pada daerah yang berbatasan dengan nukleus traktus solitarius.
Pembuluh darah pada AP
Secara teoritis , AP kaya akan pembuluh darah berfenestrasi (Ganong 2000; Buller 2001). Namun, pengamatan histologi pada penelitian ini tidak menemukan adanya fenestra tersebut. Pada fetus yang masih muda, pembuluh darah di AP berukuran kecil dan masing-masing berdiri sendiri (panah putih
Gambar 22. Jalur akson katekolamin ventral ascenden catecholamine pathway (acpv) di medula oblongata pada F 120 dengan pewarnaan
imunohistokimia terhadap TH.
Jalur akson katekolamin menghubungkan neuron katekolaminergik Grup A1 yang terdapat di nukleus retikularis lateralis, nukleus ambigus, dan daerah intermedia dengan Grup A2 yang terdapat di nukleus traktus solitarius, nukleus motor dorsalis dari nervus vagus, dan nukleus hipoglosus. Tanda panah menunjukkan jalur katekolaminergik yang mengarah ke dorsomedial
NRL = nukleus retikularis lateralis NA = nukleus ambigus NTS = nukleus traktus solitarius NH = nukleus hipoglosus NMDNV = nukleus motor dorsalis nervus vagus
Gambar 23. Sayatan koronal daerah AP pada F 100 dengan pewarnaan imunohistokimia terhadap TH.
Densitas neuron dan serabut ir-TH positif di daerah dorsal lebih besar dibandingkan dengan daerah sentral.
D = dorsal E = lapisan ependimal S = sentral
pada Gambar 14). Pembuluh darah tersebut kemudian saling beranastomose sehingga pada fetus umur 145 hari tampak seperti jala-jala. Gambar 24 memperlihatkan pembuluh darah di area postrema saling beranastomosis pada anak umur 10 hari (P 10). Sebagian neuron KA terletak berdekatan dengan pembuluh darah dan banyak memiliki prosesus sitoplasma, terutama dendrit di ruang perivaskuler.
Gambar 24. Sayatan koronal area postrema dengan pewarnaan imunohistokimia terhadap TH pada P 10.
Pembuluh darah pada AP tampak saling beranastomosis, dengan neuron katekolaminergik dan serabut katekolaminergik di dekatnya.
pd = pembuluh darah a = neuron katekolaminergik b = serabut katekolaminergik
Pembahasan Anatomi area postrema
Secara anatomis AP tersusun atas lapisan ependimal, subependimal, dan bagian sentral. Berdasarkan pewarnaan imunohistokimia terhadap TH, intensitas pewarnaan tinggi dapat dilihat pada daerah dorsal dibandingkan daerah sentral. Hal ini menunjukkan adanya aktivitas sintesa katekolamin yang lebih tinggi pada daerah dorsal tersebut. Di medula oblongata, neuron KA juga ditemukan di luar AP, yakni di daerah dorsomedial dan ventrolateral. Menurut Felten dan Sladek (1983), serta Kitahama et al. (1994), neuron KA di daerah dorsomedial medula oblongata merupakan neuron KA Grup A2, dapat ditemukan di nukleus traktus solitarius, nukleus motor dorsal dari nervus vagus, dan nukleus hipoglosus. Selain itu, neuron KA juga ditemukan di daerah ventrolateral medula oblongata, yang merupakan Grup A1 dan ditemukan pada daerah nukleus ambiguus dan nukleus retikularis lateralis. Pada formasio retikularis sentral dapat dijumpai adanya neuron Grup A1 yang berdekatan dengan Grup A2.
Perkembangan morfologi neuron KA di AP
Pada F 40, neuron KA di daerah kaudal otak belum ditemukan (Gambar 13). Diduga, neuron tersebut sudah terbentuk tetapi belum mengalami diferensiasi lebih lanjut, sehingga belum dapat mengekspresikan TH. Dugaan ini didukung oleh laporan Levitt dan Rakic (1982) yang menyatakan bahwa neuron monoaminergik di batang otak bagian kaudal monyet Rhesus sudah terbentuk pada embrio umur 35-43 hari dan mencapai puncaknya pada umur antara 38-40 hari. Secara umum, tahap-tahap perkembangan neuron adalah terbentuknya (lahirnya) neuron, diikuti dengan proliferasi, migrasi, diferensiasi, dan terakhir proses maturasi. Proses diferensiasi dapat terjadi secara tumpang tindih bersama dengan proses proliferasi dan migrasi (Shepherd 1983; Insel 2000; Morilak et al. 2000).
Pada penelitian ini, neuron KA di daerah medula oblongata mulai teramati pada F 55 (setara dengan embrio manusia umur 12,5 minggu), sedangkan menurut Verney (1999), pada embrio manusia neuron tersebut sudah ditemukan pada umur 4,5 minggu. Foster (1994) menyatakan bahwa, neuron KA di daerah
medula oblongata pada tikus ditemukan lebih lambat dibanding MEP, yaitu pada umur 13 hari kebuntingan, yang setara dengan fetus manusia umur 23,5 minggu dan fetus MEP umur 102 hari. Hal ini menunjukkan bahwa berdasarkan lamanya periode kebuntingan, perkembangan neuron KA di medula oblongata manusia terjadi paling cepat, diikuti monyet, kemudian tikus. Menurut Verney (1999), perbedaan waktu awal keberadaan neuron KA tersebut diduga akibat perkembangan sistem katekolaminergik di medula oblongata mamalia sudah terpola dan dipertahankan secara filogenetik.
Neuron di daerah bakal AP pada F 55 berada pada zona subependima dan zona intermedia. Neuron tersebut berbentuk bulat dengan inti besar dan sitoplasma sedikit (Gambar 14). Neuron dengan morfologi seperti ini merupakan neuron yang masih muda, aktif bermitosis sehingga intinya besar. Menurut La Velle dan La Velle 1970 dalam Lorke et al. (2003), pada sitodiferensiasi terjadi tiga tahap perkembangan morfologi neuron secara umum yaitu selama awal perkembangan neuron bersifat apolar, berbentuk bulat dan tidak mempunyai prosesus. Pada stadium perkembangan tengah neuron berbentuk bipolar memiliki satu atau dua prosesus sitoplasma yang pendek. Pada stadium perkembangan akhir, sebagian besar neuron berbentuk multipolar dengan beberapa prosesus sitoplasma dan ujung prosesus yang sudah mencapai tempat inervasi.
Neuron KA di AP pada F 100 berada dalam stadium perkembangan menengah dengan teramatinya neuron bipolar yang memiliki prosesus sitoplasma yang pendek. Pada P 105 dijumpai neuron KA di AP yang menciri sebagai neuron pada stadium perkembangan bentuk akhir yaitu berbentuk bipolar dengan prosesus sitoplasma yang panjang. Hal ini didukung oleh penemuan Fuxe dan Owman (1965), neuron KA di AP pada mamalia dewasa berbentuk oval (bipolar) atau bulat dengan satu atau dua prosesus sitoplasma.
Berdasarkan pengamatan dengan mempergunakan teknik fluoresen, pada F 145, ukuran neuron KA di AP sudah lebih besar dibanding F 70 dan F 55 (Gambar 19a, 20a, 21a). Hal ini menunjukkan bahwa bertambahnya volume otak dalam proses perkembangan terjadi karena proliferasi sel dan bertambahnya ukuran sel tersebut.
Migrasi neuron KA pada medula oblongata dan distribusinya
Distribusi neuron KA di daerah bakal AP serta pada bagian sentral dan ventrolateral rombensefalon bagian kaudal pada F 55 menununjukkan bahwa neuron KA bermigrasi ke arah ventrolateral. Migrasi neuron KA dimulai dari daerah bakal AP, ke bagian sentral rombensefalon dan kemudian mencapai daerah yang akan menjadi posisi definitifnya di ventrolateral (Gambar 17). Hal ini juga terjadi pada embrio manusia umur 4,5 minggu yaitu terdistribusinya neuron KA pada rombomer ke 6 dan 7 di bagian basal plate untuk calon Grup A2/C2 dan alar plate untuk calon A1/C1 (Verney 1999).
Menurut Levitt dan Rakic (1982) gradien perkembangan neuron monoaminergik pada monyet Rhesus di bagian medula oblongata berjalan ke arah rostrokaudal, sedangkan menurut Verney (1999), pada manusia arah perkembangannya ke dorsoventral.
Pada F 70, AP tampak menonjol ke ventrikel IV dan mengandung neuron KA. Neuron KA di daerah dorsomedial dan ventrolateral medula oblongata bertambah banyak yang menunjukkan bertambah pesatnya proliferasi dan migrasi neuron (Gambar 18). Verney (1999) juga menemukan gambaran bertambahnya neuron KA di medula oblongata pada embrio manusia umur 6 minggu dibandingkan pada embrio umur 4,5 minggu.
Keberadaan neuron KA di AP sejak fetus MEP umur 55 hari sampai anak umur 105 hari pada penelitian ini merupakan suatu informasi yang baru, mengingat belum adanya data tentang perkembangan neuron KA di AP sejak masa prenatal sampai postnatal. Felten dan Sladek (1983) tidak menyebutkan tentang keberada an neuron KA di AP pada monyet Rhesus dan squirrel monkey, juga didukung oleh informasi dari Kitahama et al. (1994) tentang tidak adanya neuron KA tersebut di AP monyet Jepang. Namun Schreihofer et al. (1997) menyatakan adanya neuron KA di AP pada monyet Rhesus dan MEP, tetapi karena neuronnya berukuran kecil sehingga sulit diamati.
Verney (1999) melaporkan keberadaan neuron KA di AP pada fetus manusia umur 4 bulan (setara dengan fetus MEP umur 74 hari) tetapi tidak menjelaskan tentang keberadaan neuron tersebut pada sampel yang lebih muda
umur 24 minggu). Keberadaan neuron KA di AP pada manusia dilaporkan ada pada fetus umur 14,5 sampai 25 minggu, yang setara dengan fetus umur 63–109 hari pada MEP (Lorke et al . 2003), tetapi tidak pada manusia dewasa (Pearson et al. 1983; Arango et al. 1988; Kitahama et al. 1994), sehingga keberadaannya pada masa prenatal diduga hanya bersifat sementara, selanjutnya neuron menghilang melalui proses apoptosis. Belum ada pustaka yang menyatakan keberadaan neuron KA di AP pada fetus manusia sebelum 14,5 minggu. Pada mamalia lain seperti kelinci, marmut dan anjing (Fuxe dan Owman 1965), tikus (Armstrong et al. 1982; Micheli et al. 1987), kucing (Beleslin
et al. 1989; Maqbool et al. 1993), monyet Rhesus dan MEP (Schreihofer et al.
1997), ditemukan keberadaan neuron KA tersebut di AP, tetapi informasi lebih lanjut tentang perkembangan prenatal dan postnatal neuron tersebut belum ada. Untuk itu diperlukan penelitian lebih lanjut tentang keberadaan kelompok neuron KA di AP dari masa anak sampai dewasa, agar proses perkembangan dan menghilangnya neuron tersebut pada manusia dewasa dapat lebih dipahami.
Berdasarkan pengamatan pola migrasi dan distribusi neuron KA di medula oblongata, diduga neuron KA di AP pada MEP merupakan sel bakalan bagi neuron KA yang ada di medula oblongata, yaitu neuron noradrenergik Grup A1 dan A2 serta neuron adrenergik Grup C1 dan C2.
Diferensiasi menjadi neuron dopaminergik dan noradrenergik
Diferensiasi yang terjadi pada stadium awal perkembangan menunjukkan adanya kemampuan neuron tersebut dalam menghasilkan enzim TH, yang merupakan rate limiting enzyme dalam sintesa katekolamin. Bereaksinya neuron tersebut dengan antibodi terhadap enzim TH menunjukkan bahwa neuron tersebut merupakan kelompok neuron katekolaminergik.
Verney (1999) menyatakan bahwa setelah terbentuk neuron yang ir-TH, neuron akan cepat mengalami diferensiasi. Munculnya neuron KA memperlihatkan, bahwa pada tahap ini neuron KA sudah mengalami diferensiasi bersamaan dengan proses proliferasi dan migrasi dalam neurogenesis. Menurut Morilak et al. (2000) neurogenesis tidak harus diawali dari terbentuknya sel
(lahirnya sel), proliferasi kemudian migrasi, diferensiasi, maturasi dan kematian sel, namun proses diferensiasi sel dapat berlangsung secara bersamaan dengan proses migrasi.
Dengan teknik pewarnaan label ganda terhadap TH dan DBH, dapat ditemukan bahwa pada F 55 neuron NA di AP sudah ada walaupun dalam jumlah sedikit. Bersamaan dengan bertambahnya umur kebuntingan terjadi peningkatan proporsi neuron NA di AP. Pada F 70, neuron NA tampak lebih banyak dibandingkan dengan F 55, dan semakin meningkat pada F 145, tetapi gambaran AP secara keseluruhan menunjukkan bahwa neuron DA lebih dominan. Hal ini sesuai dengan pendapat Kalia et al. (1985) dan Maqbool et al. (1993) bahwa pada AP neuron dopaminergik merupakan komponen utama neuron katekolaminergik. Pengaturan pembentukan neuron menjadi neuron dopaminergik atau neuron noradrenergik dilakukan oleh materi genetik yang ada pada sel tersebut. Diferensiasi suatu neuron diduga sudah ditentukan takdirnya (cell fate), akan menjadi neuron DA atau neuron NA oleh faktor transkripsi AP-2, seperti hasil penelitian Kim et al. (2001) yang menunjukkan adanya faktor transkripsi gen AP-2 yang mengatur ekspresi dua macam enzim yang mensintesa noradrenalin, yaitu TH dan DBH pada neuron NA. Tetapi, pada neuron dopaminergik tidak ditemukan faktor transkripsi AP-2 ini.
Berdasarkan hasil pewarnaan dengan antibodi terhadap DBH seperti yang ditampilkan pada Gambar 19, 20, dan 21, tampak bahwa meskipun badan neuron DBH sedikit, tetapi serabut yang DBH banyak ditemukan. Serabut ir-DBH adalah serabut noradrenergik atau adrenergik, tetapi bukan dopaminergik. Hal ini menunjukkan bahwa AP juga mendapat masukan noradrenergik dan adrenergik dari daerah lain, seperti yang dinyatakan oleh Armstrong et al. (1981) bahwa pada AP ditemukan adanya input adrenergik dan mungkin juga noradrenergik.
Dalam kaitannya dengan proses muntah, neuron dopaminergik yang ada di AP diduga ikut berperan untuk memacu terjadinya muntah. Hal ini didukung oleh adanya reseptor dopamin D2 di AP yang peka terhadap bahan kimia emetik dan merangsang aktivasi reseptor tersebut yang menyebabkan terjadinya
Terbentuknya jalur akson katekolamin di medula oblongata
Jalur katekolamin mulai ditemukan pada F 120. Jalur akson ini menghubungkan neuron KA Grup A2 dengan A1, dan diduga berasal dari daerah ventrolateral menuju ke bagian dorsomedial medula untuk bergabung dengan akson dari Grup A2. Jalur ini menurut Felten dan Sladek (1983) disebut dengan
ventral ascending catecholamine pathway (acpv), menuju ke hipotalamus,
trunkus ensefalikus, dan basal telensefalon. Terbentuknya jalur katekolaminergik merupakan tanda bahwa neuron KA di medula oblongata sudah mengalami diferensiasi lebih lanjut membentuk konektivitas antar neuron dan jalur akson, sehingga dapat saling berkomunikasi. Terbentuknya jalur akson ini juga menandakan bahwa neuron KA di AP sudah siap berfungsi.
Mengingat fungsi neuron KA di medula oblongata yang sangat penting dalam mengatur fungsi sistem otonom seperti sistem respirasi (Gozal dan Gaultier 2001) dan sistem kardiovaskular (Yamamoto et al. 2003), maka kesiapan neuron KA untuk berfungsi pada periode ini (kebuntingan 120 hari setara dengan umur 27,5 minggu pada manusia), merupakan faktor yang sangat mendukung kehidupan janin jika terjadi kelahiran prematur. Perlu penelitian lebih lanjut tentang perkembangan sistem katekolaminergik di daerah pons dan
midbrain, yang berperan juga dalam pengaturan sistem otonom. Dengan
penelitian lanjutan ini akan dapat diketahui, apakah kesiapan daerah tersebut sejalan dengan pembentukan jalur akson di medula oblongata.
Jalur katekolaminergik terus mengalami peningkatan pembentukan sampai pada masa postnatal. Mulai F 120 sampai P 105 terjadi proses maturasi neuron yang ditandai dengan terbentuknya konektivitas antar neuron, yang ditandai dengan banyaknya komponen serabut syaraf. Pada saat ini, diduga terjadi juga proses pembentukan sinaps. Sebagai bagian dari masa perkembangan, periode ini merupakan periode kritis terhadap gangguan perkembangan sistem katekolaminergik yang dapat berakibat fatal, karena menyebabkan tidak berfungsinya sistem respirasi dan kardiovaskular yang sangat penting untuk kelangsungan hidup setelah lahir. Neuron KA di AP, Grup A2 dan A1 berperan besar dalam proses respirasi untuk mengatasi kondisi hipoksia (Gozal dan Gaultier 2001) dengan cara meningkatkan frekuensi
pernafasan. Adanya inhalasi gas CO atau nikotin yang merupakan komponen besar dalam asap rokok pada ibu hamil terbukti menimbulkan angguan perkembangan pada sistem respirasi dan kardiovaskuler pada janin (Ruggiero et
al. 1999; Talcos et al. 2000; Gozal dan Gaultier 2001). Terganggunya
perkembangan neuron KA di AP ini diduga menjadi salah satu penyebab terjadinya sudden infant death syndrome (SIDS), yaitu kematian mendadak pada bayi yang biasanya terjadi pada saat bayi tidur. Pada pengamatan histologis medula oblongata pada bayi yang meninggal akibat SIDS, ditemukan adanya penurunan secara signifikan jumlah serabut ir-TH di daerah yang mengontrol sistem respirasi dan kardiovaskular, yaitu di AP, nukleus traktus solitarius, nukleus retikularis sentralis (Talcos et al. 2000), serta di nukleus motor dorsalis dari nervus vagus, dan area ventrolateralis (Ozawa et al. 1999; Talcos et al. 2000). Janin yang terpapar nikotin selama masa prenatal, secara signifikan akan mengalami gangguan maturasi neuron dan pembentukan sirkuitnya, sehingga menyebabkan terjadinya respon abnormal pada kondisi hipoksia (Gozal dan Gaultier 2001) yang mengarah pada terjadinya SIDS.
Terbentuknya jalur akson katekolaminergik di medula oblongata pada hari ke- 120 kebuntingan menunjukkan bahwa sistem homeostasis berkaitan dengan respirasi, kardiovaskuler sudah siap ketika memasuki trimester ke tiga kebuntingan.
Intensitas pewarnaan neuron KA di AP
Bagian dorsal AP pada penelitian memperlihatkan intensitas pewarnaan terhadap TH yang lebih besar dibanding dengan daerah sentral. Intensitas yang lebih kuat pada neuron dan daerah antar neuron ini menunjukkan bahwa daerah tersebut memiliki kandungan enzim TH lebih banyak. Intensitas yang lebih kuat di bagian dorsal jika dibandingkan dengan daerah sentral menunjukkan bahwa AP bagian dorsal lebih aktif mensintesis katekolamin dibanding daerah sentral. Padatnya serabut yang ir-TH di bagian dorsal AP ini diduga karena adanya masukan akson dari daerah lain, sesuai dengan pendapat Gozal dan Gaultier (2001), bahwa padatnya TH pada bagian antar neuron di AP diduga karena
maupun dari sistem syaraf perifer. Akson terminal katekolaminergik yang padat di bagian dorsal AP ini menurut Armstrong et al. (1982) diduga akibat adanya proses pelepasan neurotransmiter ke neurofil untuk mempengaruhi kerja sel ependima maupun sel lain yang ada didekatnya.
Pembuluh darah pada AP
Pada fetus yang masih muda, pembuluh darah di AP memiliki ukuran kecil dan saling terpisah satu dengan lainnya. Pembuluh darah tersebut kemudian saling beranastomosis, sehingga pada F 145 terbentuk bangunan seperti jala-jala (unpublished data). Bentukan ini ditunjukkan oleh Gambar 24 yang memperlihatkan pembuluh darah di AP yang saling beranastomose. Secara teoritis, pembuluh darah yang mensuplai AP berfenestrasi, namun dalam penelitian ini tidak ditemukan bentuk fenestra tersebut. Pembuluh darah tersebar sama banyak antara daerah dorsal dengan daerah sentral AP.
Menurut pengamatan Armstrong et al. (1982), serabut syaraf yang letaknya berdekatan dengan pembuluh darah adalah dendrit. Dendrit tersebut menjulur sampai ruang perivaskular dan diduga berfungsi untuk mengatur permeabilitas pembuluh darah serta mengatur aliran darah di AP. Hal yang sama juga dijumpai pada beberapa neuron KA yang memiliki prosesus sitoplasma yang letaknya berdekatan dengan pembuluh darah di medula oblongata, terutama dendrit di ruang perivaskuler. Neuron KA tersebut diduga kuat berperan dalam pengaturan permeabilitas pembuluh darah atau mempengaruhi aliran darah pada AP yang mirip dengan peran noradrenalin pada batang otak. Kedekatan posisi serabut katekolaminergik dengan pembuluh darah diduga terkait erat dengan fungsinya sebagai kemoreseptor (Arango et al. 1988).