PET Imaging of Transgene Expression
Duncan C. MacLaren, Tatsushi Toyokuni, Simon R. Cherry, Jorge R. Barrio,
Michael E. Phelps, Harvey R. Herschman, and Sanjiv S. Gambhir
A vital step in transgenic animal study and gene therapy is the ability to assay the extent of transgene expression. Unfortunately, classic methods of assaying transgene expression require biopsies or death of the subject. We are developing techniques to noninvasively and repetitively determine the location, duration, and magnitude of trans-gene expression in living animals. This will allow inves-tigators and clinicians to assay the effectiveness of their particular experimental and therapeutic paradigms. Of radionuclide (single photon emission computed tomogra-phy, positron emission tomography [PET]), optical (green fluorescent protein, luciferase), and magnetic (magnetic resonance imaging) approaches, only the radionuclide approach has sufficient sensitivity and quantitation to measure the expression of genes in vivo. We describe the instrumentation involved in high resolution PET scanning. We also describe the principles of PET reporter gene/ reporter probe in vivo imaging, the development of two in vivo reporter gene imaging systems, and the validation of our ability to noninvasively, quantitatively, and repeti-tively image gene expression in murine viral gene transfer and transgenic models. We compare the two reporter gene systems and discuss their utility for the study of transgenic animals and gene therapies. Finally, we mention alterna-tive approaches to image gene expression by using radio-labeled antibody fragments to image specific proteins and radiolabeled oligonucleotides to image RNA messages directly. Biol Psychiatry 2000;48:337–348 © 2000 So-ciety of Biological Psychiatry
Key Words: Transgene imaging, PET, gene expression, herpes simplex virus thymidine kinase, dopamine 2 recep-tor, reporter gene
Introduction
I
n this review we describe the technology involved in monitoring reporter gene expression in living animals, using positron emission tomography (PET). First we address the biological and clinical significance of studying gene expression in mouse model systems and how it applies to human diseases. Then we cover the technology involved in imaging gene expression using classic (e.g.,b-galactosidase [bgal], alkaline phosphatase [AP], lucif-erase, fluorescent proteins) and radionuclide (e.g., PET) methods. We also describe and compare current PET reporter gene imaging methods presently in use to quan-titatively assay gene expression in living animals. We then comment on potential new technologies to image gene expression and, finally, discuss the general state, utility, and future of using PET to image gene expression.
Human Diseases and Mouse Model Systems
Advances in molecular biology have allowed us to inves-tigate the function of genes and the role of various regulatory/promoter regions in human diseases by study-ing their role in mouse model systems. For example, mutations in the p53 gene can lead to many cancers in both human and murine subjects (Culver and Blaese 1994; Dasika et al 1999; Lewin 1994). Studies of the promoter and regulatory regions of genes in mouse models have also led to an understanding of human diseases where the expression of the gene under consideration is either attenuated or accentuated. Following gene therapy in animal models of Parkinson’s and Alzheimer’s diseases could lead to treatments for human patients.
Reporter Genes
The role and function of the promoter and regulatory regions of genes are often measured by their regulated expression of a reporter gene (e.g.,bgal and AP; Forss-Petter et al 1990; Lewin 1994; Naciff et al 1999). The reporter gene can either be fused to the gene of interest to make a chimeric protein of questionable function or be expressed as a separate protein. In the case where the reporter gene is expressed as a separate protein, its expression can be controlled by the same but separate promoter, or the reporter gene can be expressed as part of a bicistronic message through the use of an internal From The Crump Institute for Biological Imaging (DCM, TT, SRC, MEP, HRH,
SSG), the UCLA/DOE Laboratory of Structural Biology & Molecular Medi-cine (DCM, TT, SRC, JRB, MEP, HRH, SSG), the Department of Molecular & Medical Pharmacology (DCM, TT, SRC, JRB, MEP, HRH, SSG), the Molecular Biology Institute (DCM, HRH), the UCLA–Jonsson Comprehensive Cancer Center (HRH, SSG), and the Department of Biomathematics (MEP, SSG), UCLA School of Medicine, Los Angeles, California.
Address reprint requests to Duncan C. MacLaren, University of California at Los Angeles, Dept. of Pharmacology, 611 Charles E. Young Dr. East, 341 Boyer Hall, Los Angeles CA 90095-1570.
Received February 3, 2000; revised May 31, 2000; accepted June 14, 2000.
© 2000 Society of Biological Psychiatry 0006-3223/00/$20.00
ribosome entry site (IRES; Levenson et al 1998). Unfortu-nately, assaying these classic reporter gene methods requires biopsies or even death of the subject, and thus leaves out the possibility of true noninvasive longitudinal studies. Methods to image gene expression in animals that are small or largely transparent to visible light include the use of fluorescent proteins (e.g., green fluorescent protein; Misteli and Spector 1997) and luciferases (firefly and renilla luciferases; Nish-iyama et al 1985). Technologies have also been developed to qualitatively image luciferase gene expression in small ani-mals such as mice by using systemic delivery of an enzyme
substrate (e.g., luciferin) and extremely sensitive cameras (Contag et al 1998; Sweeney et al 1999). A method applied more recently to noninvasively, repetitively, and quantita-tively image gene expression uses PET reporter genes
Figure 1. (A) Schematic for imaging probe-modifying positron emission tomography (PET) reporter gene (PRG) expression. The enzymatic reporter gene complex is transfected into target cells. Inside the transfected cell, the transgene is transcribed to messenger RNA (mRNA) and then translated to the enzyme. The radiolabeled probe is modified by the enzyme and “trapped” within the cell. Thus, after a period of time where the unbound label is “washed” from the system, the magnitude of reporter probe accumulation in the cell reflects the level of enzyme activity and level of PRG expression. (B) Schematic for imaging receptor/ligand-based PRG expression. The reporter gene once delivered to a cell by a vector of choice is transcribed to mRNA and then translated to a ligand-binding protein or receptor. Accumulation of radiolabeled probe by the protein in or on the cell reflects the reporter gene expression level.
Figure 2. Bicistronic vector expressing two genes by a common promoter. Two different processes translate the single messenger RNA (mRNA) transcript into two separate proteins. Cap-depen-dent translation produces the initial (therapeutic) protein, and the internal ribosome entry site (IRES) allows the ribosome to translate the following gene and produce the reporter protein. By imaging the reporter protein one can infer therapeutic gene
expression. Figure 3. Comparison of positron emission tomography (PET)
imaging systems. A nude (NU/NU) mouse was imaged with a radiolabeled probe, 3-(29-[18
and radio-labeled PET reporter probes (for reviews, see Gambhir et al 1999a; Herschman et al 2000).
PET Reporter Genes
Positron emission tomography reporter genes encode receptors that bind positron-emitting ligand probes or enzymes that modify the positron-emitting substrate probes to produce sequestered positron-emitting prod-ucts. Cells expressing the PET reporter gene will sequester the radiolabel of the PET reporter probe 1) as a ligand bound to the PET reporter receptor or 2) as a “trapped” product of the enzymatic reaction of the PET reporter enzyme (Figure 1; also reviewed in Gambhir et al 1999a; Herschman et al 2000). Ideally, those cells not expressing the PET reporter genes will not retain the PET reporter probe. Since positron-emitting radionu-clides result in the creation of high-energy gamma rays (511 keV) by positron– electron annihilation, the animal is largely transparent to the wavelength of the radiation produced, and visualization of radiolabeled probe/li-gand accumulation is readily obtained in even deep and visually opaque tissues. Emission computed tomogra-phy then allows quantitative imaging of the accumula-tion of the PET reporter probe and, in turn, the expression levels of the PET reporter gene. Since PET imaging does not require obtaining tissue samples from the subject, this system is noninvasive and can be used to repetitively measure reporter gene expression in vivo.
It is also important to note that radionuclide-based methods offer significant advantages over optical- (Contag et al 1998) and magnetic resonance imaging– based (Bog-danov and Weissleder 1998) approaches for imaging reporter gene expression. Radionuclide-based methods
offer the highest level of sensitivity for imaging relatively low levels of reporter gene expression—as low as 10212 mol/L of radiolabeled substrate (Phelps 1991; Phelps et al 1986). This high degree of sensitivity may allow the use of relatively weak promoters and the imaging of relatively low levels of gene expression. Furthermore, radionuclide-based methods are highly quantitative. Since PET allows for the quantitation of absolute levels of radionuclide probes, dynamic imaging and kinetic modeling allow us to obtain the rate constants of the underlying biochemical processes (Green et al 1998; Huang and Phelps 1986).
One of the most basic considerations is whether the reporter gene is endogenous or exogenous for the organism under scrutiny. Expression of endogenous genes has the advantage of not inducing an immune response and thus allowing for repeated studies. A possible complication in using an endogenous gene as a PET reporter transgene is a high background or erroneous signal due to its inherent expression. Exogenous genes, especially in gene therapy use, have the disadvantage of inciting an immune response that might limit their repeated application, unless, of course, one wishes to image the immune response itself. Exogenous PET reporter genes, with the appropriate probe, have the advan-tage of only producing signal in the tissues in which they are expressed. An ideal reporter gene for longitudinal studies should therefore produce no immune response and not be normally expressed in the organism— or at least in the organ(s) under consideration.
Direct and Indirect Reporter Transgene Strategies
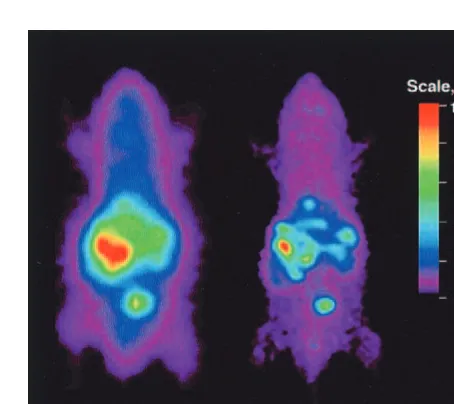
Direct imaging strategies are based on imaging the trans-gene product directly by binding of a radiolabeled ligand or probe directly to the gene product (e.g., messenger RNA [mRNA], dopamine 2 receptor [D2R]; MacLaren et Figure 4. 8-[18
F]Fluoroganciclovir (FGCV) can be used as a positron emission tomography (PET) reporter probe to image herpes simplex virus 1 thymidine kinase (HSV1-TK) expression. For the microPET images, mice were injected with 13109
al 1999). Another “direct” approach involves an enzymatic strategy where a radiolabeled probe is modified and trapped in cells expressing the PET reporter gene (e.g., HSV1-tk; Gambhir et al 1998a, 1999d, 2000a; Tjuvajev et al 1995, 1998).
Most therapeutic transgenes do not lend themselves to direct imaging of the transgene product. This is because most therapeutic transgene products lack appropriate ra-diolabeled probes. In addition, it would be very time consuming and inefficient to develop and validate new probes for each therapeutic transgene. Alternatively, it is both feasible and reasonable to develop and validate “indirect” imaging strategies using a reporter gene in combination with a therapeutic gene. The advantage of this paradigm is that a given reporter gene can be coupled with any therapeutic gene. Two strategies have been discussed: one uses a fusion gene containing complemen-tary DNA from both reporter and therapeutic genes, and the second strategy has a separate reporter gene and therapeutic gene on the same mRNA transcript (cis-linked). Both strategies are based on demonstrating a proportional and constant relationship in the coexpression of two transgenes over a wide range of expression levels. Researchers have described and validated the propor-tional expression of two cis-linked genes, using an IRES element within a single bicistronic transcription unit (Tju-vajev et al 1999; Yu et al 2000). The IRES element enables translation initiation at a downstream start codon within the bicistronic mRNA, thus permitting gene coex-pression by cap-dependent translation of the first cistron and cap-independent, IRES-mediated translation of the second cistron (Figure 2; Ghattas et al 1991; Jackson and Kaminski 1995; Pelletier 1988; Sachs et al 1997). These studies show that coexpression of the genes is proportional and quantitative and also demonstrate the potential for monitoring therapeutic gene transfer and expression by noninvasive imaging of cis-linked PET reporter genes (Tjuvajev et al 1999; Yu et al 2000).
PET Instrumentation
Because of their short breeding span, extensively charac-terized genetics, and readily manipulatable genome, mice have become the primary platform for most whole-animal research in gene expression, gene transfer, and models of human disease. The desire to image mouse models of human disease has led to rapidly increasing interest and efforts in developing imaging technologies that can mea-sure the distribution of radiolabeled tracers in vivo in the mouse (Weber and Ivanovic 1999). Autoradiography is a well-established technique that requires killing the animal of interest (Lear 1986, 197–235) and placing tissue slices in direct contact with analog or digital film. Although
autoradiography has and continues to play a key role, noninvasive approaches are highly desirable for applica-tions in which the same animal needs to be repeatedly evaluated, or for applications in which it is too expensive to study large sets of animals at various time points. Furthermore, for human applications, although tissue bi-opsies can be performed, whole-body noninvasive imag-ing is much more compatible with the patient’s comfort and safety. In essence, the goal for radionuclide-based imaging instrumentation has been to develop a noninva-sive in vivo analogue of autoradiography, with sufficient spatial resolution to resolve the structures of interest in a mouse and with sufficient sensitivity that high signal-to-noise images can be obtained.
Radiotracer imaging technologies that can measure the distribution of radiolabeled tracers in the human body are widely available and have a wide range of clinical and research applications. Two classes of clinical nuclear imaging systems exist—those designed to image single gamma– emitting radionuclides (e.g., technetium-99m, io-dine-131) and those designed to image positron-emitting radionuclides (e.g., fluorine-18, carbon-11, nitrogen 13, oxygen-15, copper-64, iodine-124). The former is known as single photon imaging or, when performed tomographi-cally, single photon emission computed tomography (SPECT). The latter is known as PET. In general, PET has greater spatial resolution and higher sensitivity and is easier to quantify than SPECT. For the sake of brevity, this review focuses on PET, though most of the discussion is also applicable to other radionuclide systems.
microPET scanner has a reconstructed image resolution of 1.8 mm in all three axes and has been shown to be fully quantitative (Chatziioannou et al 1999). The volumetric resolution is more than an order of magnitude better than state-of-the-art clinical PET systems, as illustrated by the images in Figure 3.
Gene Therapy and Reporter Genes
Our understanding of molecular biology in the last de-cades allowed us to create transgenic animals as well as to introduce genes into living animals (i.e., gene therapy). A vital step in either of these processes is the ability to assay for the expression of the transgene. Thus, the use of PET reporter genes can play critical roles in developing gene therapies by allowing researchers to determine the loca-tion, duraloca-tion, and expression level of the transferred DNA and, specifically, 1) develop vector modifications to prove delivery, 2) control expression levels, and 3) im-prove treatments to control duration of expression. Appli-cations of somatic gene transfer technology to treat diseases are at the forefront of gene therapy applications; as a result, these issues are of great interest to life scientists and clinicians. The repeatability, quantifiability, and high sensitivity of PET reporter gene systems should lead to rapid advancements in science and medicine.
Herpes Simplex Virus 1 Thymidine
Kinase (HSV1-tk), an Enzymatic PET
Reporter Gene
Herpes simplex virus 1 thymidine kinase, like mammalian TKs, phosphorylates thymidine, but unlike mammalian TKs, HSV1-TK has relaxed substrate specificity and so phosphorylates thymidine analogues (e.g., 5-iodo-29 -fluoro-29deoxy-1-b-D-arabino-furanosyl-uracil [FIAU]) as well as acycloguanosine analogues (e.g., acyclovir, ganci-clovir [GCV], penciganci-clovir [PCV]; Namavari et al 2000). Cellular enzymes then convert acycloguanosine mono-phosphates and the monophosphate of FIAU to di- and triphosphates, which have been shown to kill cells by incorporation as chain-terminating derivatives and/or by inhibition of DNA polymerase; however, at the concen-trations of tracer used for imaging by PET, the derivatives have no discernable effect upon the cells or the health of patients (for review, see Gambhir et al 2000b). Herpes simplex virus 1 thymidine kinase has been extensively studied; it is nontoxic in humans and is currently being used as a “susceptibility” gene (in combination with GCV) in clinical gene therapy protocols. Herpes simplex virus 1 thymidine kinase can be used as a reporter gene as well as a therapeutic gene (Borrelli et al 1988; Culver et al 1992; Moolten 1997; Moolten and Wells 1990). In gene therapy
protocols using HSV1-tk as a susceptibility gene, identi-fying the location and magnitude of HSV1-TK expression by noninvasive imaging would provide a highly desirable measure of expression (following successful gene trans-fection) and a basis from which the timing of GCV treatment can be optimized. This represents an ideal situation where the therapeutic and reporter genes are the same, and is an example of a direct imaging approach.
Two main categories of substrates have been investi-gated as reporter probes for imaging HSV1-tk reporter gene expression: derivatives of uracil nucleoside (e.g., FIAU radiolabeled with iodine; Morin et al 1997; Tjuvajev et al 1995) and derivatives of guanosine radiolabeled with fluorine-18 or carbon-11 (Alauddin et al 1996; Barrio et al 1996a, 1996b, 1996c, 1997; Monclus et al 1995). These two major classes of reporter probes share the ability to be phosphorylated by HSV1-TK, leading to their accumula-tion in cells by DNA polymerase.
Thymidine Derivative Reporter Probes
Using a cell culture model, researchers investigated three compounds (FIAU, iododeoxyuridine, and GCV) as po-tential HSV1-tk reporter probes and found FIAU to have the best imaging potential, based upon its in vitro charac-teristics for HSV1-TK and its ability to be labeled with several different nuclides (Tjuvajev et al 1995). Radiola-beled FIAU has been used in cell culture and in vivo as an agent for imaging gene expression by both SPECT and PET systems (Tachizawa et al 1981; Tjuvajev et al 1995, 1996). Imaging HSV1-TK expression in cancer patients undergoing combined HSV1-tk–GCV gene therapy with PET or SPECT has also been evaluated and shown to be feasible (Blasberg and Tjuvajev 1997).
Guanosine Derivative Reporter Probes
accumu-lates on hepatocytes in large part due to the presence of coxsackie and adenoviral receptors (Haisma et al 1999), the majority of the injected adenovirus (.95%) infects the liver. Administration of radiolabeled reporter probes showed accumulation of PET signal only in livers express-ing HSV1-TK (Figure 4), thus validatexpress-ing the applicability of using radiolabeled acycloguanosine derivatives to im-age HSV1-tk as a PET reporter gene (Gambhir et al 1998a,
1998b, 1999c, 1999e). Panels B and C of Figure 4 also show the linear relationship between the amount of FGCV signal to the HSV1-tk activity and mRNA levels.
Reporter probes using fluorine-18 in the side chain of GCV (FHPG; Alauddin et al 1996; Bading et al 1997; Monclus et al 1997) and in the side chain of PCV (FHBG; Alauddin and Conti 1998) are also being studied and have shown that high specific activity (250 –500 Ci/mmol)
Figure 5. Fluoropenciclovir (FPCV) and the side chain of PCV (FHBG) positron emission tomography (PET) images of a trans-genic mouse expressing herpes simplex virus 1 thymidine kinase (HSV1-tk) in the liver. We studied a transgenic mouse in which the albumin promoter drives the HSV1-tk reporter gene. The mouse was imaged on day 0 with a microPET 1 hour after administration of FPCV and on day 1 with FHBG. Both images are displayed using the same common global maximum and illustrate the higher percent injected dose retained per gram of liver tissue (%ID/g) when utilizing FHBG. There is significantly greater hepatic accumulation when using FHBG (8 –11% ID/g), as compared with FPCV (3– 6% ID/g).
Figure 6. 3-(29-[18
F]Fluoroethyl)spiperone (FESP) can be used as a reporter probe to image dopamine 2 receptor (D2R) positron emission tomography (PET) reporter gene expression in living mice. Nude mice were injected via the tail vein with 13 109 plaque-forming units of adenovirus expressing eitherb-galactosidase (Ad-bGal) (A) or the dopamine 2 receptor (Ad-D2R) (B). Two days later, both mice were injected with FESP and imaged using microPET. For each mouse, a whole-body coronal projection image of the [18
[18F]fluorinated acycloguanosines can be synthesized in relatively high yields (5–15 mCi). The side chain of GCV and FHBG have been studied in cell culture models and in vivo, and are also well suited for imaging HSV1-tk gene expression (Gambhir et al 2000b).
Comparison of HSV1-tk Reporter Probes
Direct comparison of all the radiolabeled probes for in vivo imaging of HSV1-tk reporter gene expression is a necessary step for optimizing this PET reporter system. The UCLA group has compared uptake of FGCV, FPCV, FHBG, FHPG, and [14C]-FIAU in C6 cells expressing HSV1-tk. These preliminary data show that FIAU and FHBG are the better candidates for imaging HSV1-tk reporter gene expression because of their 1) long half-life in vivo, 2) low nonspecific retention, and 3) high specific retention. The true utility of alternative HSV-TK probes must ultimately be evaluated in whole animals, as issues such as stability, substrate competi-tion, routes of clearance, and rates of cellular transport all come into play. A transgenic mouse model in which the albumin promoter drives the HSV1-tk reporter gene is being studied at UCLA (Gambhir et al 2000b; Herschman et al 2000). These transgenic mice have been imaged by PET and clearly demonstrate accumu-lation of the FPCV (3– 6% injected dose retained per gram of tissue [ID/g]) and FHBG (8 –11% ID/g) re-porter probes in the mouse liver (Figure 5). Future studies that directly compare all prospective acy-cloguanosines and thymidine analogues in whole ani-mals will help to better define the advantages and disadvantages for each probe.
Mutant HSV1-tk Reporter Gene Approaches
Researchers at UCLA are also investigating mutant HSV1-tk reporter genes to further enhance the sensitivity of the HSV1-tk reporter assays (Gambhir et al 2000a). We are using a mutated HSV1-TK enzyme (HSV1-sr39TK) that utilizes GCV and PCV substrates more effectively and thymidine less effectively than the wild-type HSV1-TK enzyme (Black et al 1996). Mouse models with an adenovirus expressing this mutant HSV1-TK enzyme demonstrated improved imaging sensitivity (equivalent to two- to threefold) with both FGCV and FPCV as PET reporter probes. Additional studies with FHBG show a further enhancement by a factor of ;2 as well. Taken together, the mutant HSV1-tk with FHBG should offer a greater than fourfold improvement in sensitivity and place it on par with the sensitivity of the D2R/FESP PET
reporter system.
D
2R, a Receptor PET Reporter Gene
Another PET reporter gene system investigated uses D2R
and the radiolabeled ligand 3-(29-[18 F]fluoroethyl)spiper-one (FESP), a positron-emitting analogue of the dopamine antagonist spiperone. The latter was originally developed as a probe to image, by PET, the D2Rs of the striatum
(Barrio et al 1989), where concentrations of D2R as low as
2–20 nmol/L can be detected. This PET reporter gene system was also investigated because of the high binding affinities of D2R to FESP and, unlike substrates for
HSV1-tk, the ability of FESP to diffuse into every tissue in the body (Barrio et al 1989).
As for the HSV1-tk system, we used adenovirus delivery of the D2R reporter gene to demonstrate the
noninvasive, repetitive, and quantitative ability of the D2R/FESP PET reporter gene/PET reporter probe in
vivo imaging system. To determine whether PET can quantitatively monitor hepatic D2R expression in
ade-novirus-infected mice, animals were injected with vary-ing amounts of D2R-expressing or bgal-expressing
(control) virus, then injected with FESP and imaged by PET (Figure 6; MacLaren et al 1999). After scanning, the mice were killed and liver samples were analyzed for 1) FESP retention by ether extraction and [fluorine-18] well counting, 2) functional D2R protein levels by
[3H]spiperone binding using a conventional receptor binding assay, and 3) D2R mRNA levels by quantitative
Northern blots. The fluorine-18 signal retained in liver, as determined by region of interest measurements of PET images of living mice, is proportional to both the amount of hepatic FESP present and functional D2R
protein levels (Figure 7). In vivo PET analysis of hepatic D2R reporter gene expression accurately reflects
in vitro determinations of D2R levels and validates the
use of this PET reporter gene/probe system.
Comparison of HSV1-tk and D
2R Reporter
Gene Systems
The reporter gene assays developed to date fall into the two main categories enzyme based (e.g., HSV1-tk) and receptor based (e.g., D2R). Each of these assays has
some distinct features that deserve special comment. An enzyme-based approach has the theoretical advantage of signal amplification, since one molecule of reporter enzyme is capable of acting on many molecules of reporter probes. Most receptor-based assays, such as the D2R system, are capable of only a one-to-one
gene expression, thereby making it more difficult to quantitate the signal observed. Receptor-based ap-proaches in which the receptor is primarily limited to the cell surface have the unique advantage of not requiring reporter probe transport.
Our current mutant HSV1-tk/FHBG and D2R/FESP PET
reporter gene imaging systems now have essentially equiva-lent sensitivities,;20% ID/g in the liver (unpublished data); however, each of these systems has distinct advantages and disadvantages. The HSV1-tk reporter gene does not meet all the requirements of an ideal system. Expression of HSV1-tk may lead to an immune response, which is not optimal from the perspective of gene therapy. An additional limitation of the HSV1-tk system is that reporter probes for this system do not significantly penetrate the intact blood– brain barrier, thus preventing the easy utilization of the HSV1-tk reporter gene imaging system for in vivo imaging of the brain. The probe used for imaging the D2R gene, FESP, has no such barrier
limitations and penetrates into all tissues. Because the D2R
gene is normally present in the mammalian genome there are no issues of an immune response, but it also does not meet all the requirements of an ideal system. Being an endogenous gene leads to a problem of background signal in the striatum where D2R is normally expressed; however, this may be
viewed as an advantage, as the signal in the striatum may serve as an internal control for measuring D2R reporter gene
expression in other tissues. Herpes simplex virus 1 thymidine kinase expression, in the absence of acycloguanosines, has little or no effect on cells and tissues. In contrast, occupancy by endogenous agonists of the ectopic wild-type D2R, either
in transgenic animals or in gene therapy delivery vehicles, might have physiologic consequences because ligand-activated D2R regulates intracellular cyclic adenosine
monophosphate levels. Fortunately, D2R structure–
function studies have identified amino acids that uncou-ple receptor occupancy from intracellular signaling (Cox et al 1992; Neve et al 1991; Woodward et al 1996), and adenovirus strains with these D2R mutations
are being developed to test the efficacy of these mutant receptors as PET reporter genes.
Alternative PET Gene Imaging Methods
There are other potential methods to image gene expres-sion that warrant mention. One is by using radiolabeled modified oligonucleotides or small molecules to bind to the mRNA transcripts directly (Charlton et al 1997; Gambhir et al 1999b; Kobori et al 1999; Mannironi et al 1997; Pan et al 1998; Tavitian et al 1998). The mRNA transcripts themselves can also be engineered or selected so that sections of them bind to specific radiolabeled small molecules (e.g., RNA aptamers; Werstuck and Green 1998). A second method is to radiolabel antibodies or antibodylike constructs that target specific cell surface– expressed proteins (Govindan et al 1996; Hu et al 1996).
Discussion and Future Directions
There are a number of scenarios in which the use of two in vivo reporter gene imaging systems will be of consider-Figure 7. Positron emission tomography (PET) image analysis using the 3-(29-[18
F]fluoroethyl)spiperone (FESP) reporter probe measures dopamine 2 receptor (D2R) reporter gene protein levels. Nude mice were injected via tail vein with adenovirus (Ad-D2R) to produce varying levels of D2R reporter gene expression. Two to 60 days after viral injection, FESP was injected via the tail vein and 3 hours later the mice were imaged by PET. The mice were then killed and their livers were removed and analyzed for FESP, FESP metabolites, D2R protein levels (by [3
able utility in research protocols. For example, the ability to image two distinct reporter genes in vivo will allow direct comparisons, in the same individual, of alternative gene delivery vehicles (e.g., viruses, liposomes) in somatic gene transfer protocols. Many combinatorial applications of these in vivo PET reporter gene imaging systems are likely to emerge as their availability and utility become apparent. The advantages of high sensitivity, quantitative capability, and direct ability to translate the developed assays from animal to human studies will keep radionu-clide-based approaches at the forefront of imaging gene expression for transgenic and gene therapy studies.
Continued development of reporter genes and corre-sponding reporter probes will be needed to produce more optimal assays; however, for many applications the sys-tems developed to date appear sufficiently robust. The major applications of PET reporter gene imaging in vivo are likely to be 1) repetitive, quantitative monitoring of the location, duration, and extent of gene expression from gene therapy vehicles, using bicistronic vectors that ex-press therapeutic and reporter gene products from a common transcript, and 2) the evaluation of gene function and the role of promoter/regulatory elements by reporter gene expression in transgenic animals during longitudinal experiments.
There are also improvements in PET instrumenta-tion—a “second-generation” microPET instrument, cur-rently being developed, will provide a much higher reso-lution of 1 mm for each axis and be able to scan an entire mouse in 10 min. Positron emission tomography technol-ogy is still a long way from reaching the resolution and sensitivity limits imposed by geometric considerations and the physics of the positron annihilation process (Chatziio-annou et al 1999; Cherry et al 1997). The performance of dedicated animal PET scanners is likely to improve substantially in the next few years and will be accompa-nied by a reduction in both their size and their cost.
The combination of improved PET reporter gene imag-ing systems and PET instrumentation will allow investi-gators to readily image and quantitatively evaluate gene expression in transgenic animals and gene therapy sub-jects. This will ultimately allow doctors to follow the effectiveness of gene therapy for many disorders, includ-ing neurodegenerative diseases such as Parkinson’s and Alzheimer’s.
These studies were supported by Department of Energy contract DE-FC03-87ER60615, National Institutes of Health Grant No. RO1 CA82214-01 (SSG), National Institutes of Health Grant No. RO1 CA84572-01 (HRH), the University of California Biotechnology Pro-gram, the UCLA Gene Medicine ProPro-gram, the Dana Foundation, and the UCLA–Jonsson Comprehensive Cancer Center.
References
Alauddin MM, Conti PS (1998): Synthesis and preliminary evaluation of 9-(4-[18F]-fluoro-3-hydroxymethylbutyl)gua-nine ([18F]FHBG): A new potential imaging agent for viral infection and gene therapy using PET. Nucl Med Biol 25:175–180.
Alauddin MM, Conti PS, Mazza SM, Hamzeh FM, Lever JR (1996): Synthesis of 9-[(3-[18F]fluoro-1-hydroxy-2-propoxy) methyl]guanine ([18F]FHPG): A potential imaging agent of viral infection and gene therapy using PET. Nucl Med Biol 23:787–792.
Bading JR, Alauddin MM, Fissekis JD, Kirkman E, Raman RK, Conti PS (1997): Pharmacokinetics of F-18 fluorohydroxy-propoxymethylguanine (FHPG). J Nucl Med 38:43P. Barrio JR, Namavari M, Phelps ME, Satyamurthy N (1996a):
Elemental fluorine to 8-fluorpurines in one step. J Am Chem
Soc 118:10408 –10411.
Barrio JR, Namavari M, Phelps ME, Satyamurthy N (1996b): Regioselective fluorination of substituted guanines with di-lute F2: A facile entry of 8-fluoroguanine derivatives. J
Organic Chem 61:6084 – 6085.
Barrio JR, Namavari M, Satyamurthy N, Srinivasan A, Her-schman H, Gambhir S (1996c): 8-[F-18]fluoroacyclovir: An in vivo probe for gene expression with PET. J Nucl Med 37:193P.
Barrio JR, Namavari M, Srinivasan A, Gambhir S, Cherry S, Herschman H, et al (1997): Carbon-8 radiofluorination of purines: A general approach to probe design for gene therapy in humans. J Labelled Compounds Radiopharmaceuticals 40:348.
Barrio JR, Satyamurthy N, Huang SC, Keen RE, Nissenson CH, Hoffman JM, et al (1989): 3-(29-[18
F]fluoroethyl)spiperone: In vivo biochemical and kinetic characterization in rodents, nonhuman primates, and humans. J Cereb Blood Flow Metab 9:830 – 839.
Black ME, Newcomb TG, Wilson H-MP, Loeb LA (1996): Creation of drug-specific hepes simplex virus type 1 thymi-dine kinase mutant for gene therapy. Proc Natl Acad Sci
U S A 93:3525–3529.
Blasberg R, Tjuvajev J (1997): In vivo monitoring of gene therapy by radiotracer imaging. In: Semmler W, Schwaiger W, editors. Ernst Shering Research Foundation Workshop
22: Impact of Molecular Biology and New Technical Devel-opments on Diagnostic Imaging. Berlin: Springer Verlag,
161–189.
Bloomfield PM, Rajeswaran S, Spinks TJ, Hume SP, Myers R, Ashworth S, et al (1995): The design and physical character-istics of a small animal positronemission tomograph. Phys
Med Biol 40:1105–1126.
Bogdanov A, Weissleder R (1998): The development of in vivo imaging systems to study gene expression. Trends Biotechnol 16:5–10.
Borrelli E, Heyman R, Hsi M, Evans RM (1988): Targeting of an inducible toxic phenotype in animal cells. Proc Natl Acad Sci
U S A 85:7572–7576.
Charlton J, Sennello J, Smith D (1997): In vivo imaging of inflammation using an aptamer inhibitor of human neutrophil elastase. Chem Biol 4:809 – 816.
K, Farquhar TH, et al (1999): Performance evaluation of microPET: A high resolution LSO PET scanner for animal imaging. J Nucl Med 40:1164 –1175.
Cherry SR, Shao Y, Silverman RW, Meadors K, Siegel S, Chatziioannou AF, et al (1997): MicroPET: A high resolution PET scanner for imaging small animals. IEEE Trans Nucl Sci 44:1161–1166.
Contag PR, Olomu IN, Stevenson DK, Contag CH (1998): Bioluminescent indicators in living mammals. Nat Med 4:245–247.
Cox BA, Henningsen RA, Spanoyannis A, Neve RL, Neve KA (1992): Contributions of conserved serine residues to the interactions of ligands with dopamine D2 receptors. J
Neu-rochem 59:627– 635.
Culver KW, Blaese RM (1994): Gene therapy for cancer. Trends
Genet 10:174 –178.
Culver KW, Ram Z, Walbridge S, Ishii H, Oldfield EH, Blaise RM (1992): In vivo gene transfer with retroviral vector-producer cells for treatment of experimental brain tumors.
Science 256:1550 –1552.
Dasika GK, Lin SC, Zhao S, Sung P, Tomkinson A, Lee EY (1999): DNA damage-induced cell cycle checkpoints and DNA strand break repair in development and tumorigenesis.
Oncogene 20:7883–7899.
Forss-Petter S, Danielson PE, Catsicas S, Battenberg E, Price J, Nerenberg M, Sutcliffe JG (1990): Transgenic mice expressing beta-galactosidase in mature neurons under neuron-specific enolase promoter control. Neuron 5:187– 197.
Fricker RA, Torres EM, Hume SP, Myers R, Opacka-Juffrey J, Ashworth S, et al (1997): The effects of donor stage on the survival and function of embryonic striatal grafts in the adult rat brain. II. Correlation between positron emission tomography and reaching behaviour. Neuroscience 79: 711–721.
Fries O, Bradbury SM, Gebauer J, Holl I, Lorenz E, Renker D, et al (1997): A small animal PET prototype based on LSO crystals read out by avalanche photodiodes. Nucl Inst
Meth-ods A387:220 –224.
Gambhir SS, Barrio J, Wu L, Iyer M, Namavari M, Satyamur-thy N, et al (1998a): Imaging of adenoviral directed herpes simplex virus type 1 thymidine kinase gene expression in mice with ganciclovir. J Nucl Med 39:2003–2011. Gambhir SS, Barrio JR, Bauer E, Iyer M, Namavari M,
Satyamurthy N, et al (1998b): Radiolabeled penciclovir: A new reporter probe with improved imaging properties over ganciclovir for imaging herpes-simplex virus type 1 thy-midine kinase reporter gene expression. J Nucl Med 39:53P.
Gambhir SS, Barrio JR, Herschman HR, Phelps ME (1999a): Assays for noninvasive imaging of reporter gene expression.
Nucl Med Biol 26:481– 490.
Gambhir SS, Barrio JR, Herschman HR, Phelps ME (1999b): Imaging gene expression: Principles and assays. J Nucl
Cardiol 6:219 –233.
Gambhir SS, Barrio JR, Iyer M, Namavari M, Satyamurthy N, Toyokuni T, et al (1999c): In vivo validation of PET reporter gene/reporter probe assay for herpes simplex virus type 1 thymidine kinase with 8-[F-18]-fluoropenciclovir. J Nucl
Med 40:25–26.
Gambhir SS, Barrio JR, Phelps ME, Iyer M, Namavari M, Satyamurthy N, et al (1999d): Imaging adenoviral-directed reporter gene expression in living animals with positron emission tomography. Proc Natl Acad Sci U S A 96:2333– 2338.
Gambhir SS, Bauer E, Black M, Liang Q, Kokoris MS, Barrio JR, et al (2000a): A mutant herpes simplex virus type 1 thymidine kinase reporter gene shows improved sensitivity for imaging reporter gene expression with positron emis-sion tomography. Proc Natl Acad Sci U S A 97:2785–2790. Gambhir SS, Herschman HR, Cherry SR, Barrio JR, Satyamur-thy N, Toyokuni T, et al (2000b): Imaging transgene expres-sion with radionuclide imaging technologies. Neoplasia 2:118 –138.
Gambhir SS, MacLaren DC, Barrio JR, Toyokuni T, Satyamur-thy N, Nguyen K, et al (1999e, May): Noninvasive and repeated imaging of reporter gene expression in living mice utilizing positron emission tomography. Poster presented at the annual meeting of the American Society of Gene Therapy, Washington, DC.
Ghattas IR, Sanes JR, Majors JE (1991): The encephalomyo-carditis virus internal ribosomal entry site allows efficient coexpression of two genes from a recombinant provirus in cultured cells and in embryos. Mol Cell Biol 11:5848 – 5859.
Govindan SV, Goldenberg DM, Grebenau RC, Hansen HJ, Griffiths GL (1996): Thiolations, 99mTc labelings, and ani-mal in vivo biodistributions of divalent monoclonal antibody fragments. Bioconjug Chem 7:290 –297.
Green LA, Gambhir SS, Barrio JR, Bauer E, Nguyen K, Namavari M, et al (1998): Tracer kinetic modeling of 8-(F18)-fluoroganciclovir PET data: A new tracer for measuring reporter gene expression. J Nucl Med 39(suppl): 10P.
Haisma H, Pinedo H, Rijswijk A, der Meulen-Muileman I, Sosnowski BA, Ying W, et al (1999): Tumor-specific gene transfer via an adenoviral vector targeted to the pan-carci-noma antigen EpCAM. Gene Ther 6:1469 –1474.
Herschman HR, MacLaren DC, Iyer M, Namavari M, Bobinski K, Green LA, et al (2000): Seeing is believing: Non-invasive, quantitative and repetitive imaging of reporter gene expres-sion in living animals, using positron emisexpres-sion tomography.
J Neurosci Res 59:699 –705.
Hu S, Shively L, Raubitschek A, Sherman M, Williams LE, Wong JY, et al (1996): Minibody: A novel engineered anti-carcinoembryonic antigen antibody fragment (single-chain Fv-CH3) which exhibits rapid, high-level targeting of xenografts. Cancer Res 56:3055–3061.
Huang SC, Phelps ME (1986): Chapter 7. In: Phelps ME, Mazziota J, Schelbert HR, editors. Positron Emission
Tomography and Autoradiography, Principles and Appli-cations for the Brain and Heart. New York: Raven.
Hume SP, Lammertsma AA, Myers R, Rajeswaran S, Bloom-field PM, Ashworth S, et al (1996): The potential of high-resolution positron emission tomography to monitor striatal dopaminergic function in rat models of disease. J Neurosci
Methods 67:103–112.
Jeavons AP, Chandler RA, Dettmmar CAR (1999): A 3D HIDAC-PET camera with sub-millimetre resolution for imaging small animals. IEEE Trans Nucl Sci 46:468 – 473. Kobori N, Imahori Y, Mineura K, Ueda S, Fujii R (1999): Visualization of mRNA expression in CNS using 11C-labeled phosphorothioate oligodeoxynucleotide. Neuroreport 10:2971– 2974.
Lear JL (1986): Principles of Single and Multiple Radionuclide
Autoradiography. New York: Raven.
Levenson VV, Transue ED, Roninson IB (1998): Internal ribo-somal entry site-containing retroviral vectors with green fluorescent protein and drug resistance markers. Hum Gene
Ther 9:1233–1236.
Lewin B (1994): Genes V. New York: Oxford University Press. MacLaren DC, Gambhir SS, Satyamurthy N, Barrio JR, Sharf-stein S, Toyokuni T, et al (1999): Repetitive, non-invasive imaging of the dopamine D2 receptor as a reporter gene in living animals. Gene Ther 6:785–791.
Mannironi C, Di NA, Fruscoloni P, Tocchini VG (1997): In vitro selection of dopamine RNA ligands. Biochemistry 36:9726 – 9734.
Marriott CJ, Cadorette JE, Lecomete R, Scasnar V, Rousseau J, van Lier JE (1994): High-resolution PET imaging and quan-titation of pharmaceutical biodistributions in a small animal using avalance photodiode detectors. J Nucl Med 35:1390 – 1396.
Misteli T, Spector D (1997): Applications of the green fluores-cent protein in cell biology and biotechnology. Nat
Biotech-nol 15:961–964.
Monclus M, Luxen A, Cool V, Damhaut P, Velu T, Goldman S (1997): Development of a positron emission tomography radiopharmaceutical for imaging thymidine kinase gene ex-pression: Synthesis and in vitro evaluation of 9-{3-[18F]fluoro-1-hydroxy-2-propoxy)methyl}guanine. Bioorg Med Chem Lett 7:1879 –1882.
Monclus M, Luxen A, Van Naemen J, et al (1995): Develop-ment of PET radiopharmaceuticals for gene therapy: Syn-thesis of 9-((1-(18
F)fluoro-3-hydroxy-2-propoxy)methyl) guanine. J Labelled Compounds Radiopharmaceuticals 37:193–195.
Moolten FL (1997): Suicide genes for cancer therapy. Sci Med 4:16 –25.
Moolten FL, Wells JM (1990): Curability of tumors bearing herpes thymidine kinase genes transfected by retroviral vec-tors. J Natl Cancer Inst 82:297–300.
Morin KW, Atrazheva ED, Knaus EE, Wiebe LI (1997): Syn-thesis and celluar uptake of 29-substituted analogues of (E)-5-(2-[125I]iodovinyl-29-deoxyuridine in tumor cells transduced with the herpes simplex type-1 thymidine kinase gene. Evaluation as probes for monitoring gene therapy.
J Med Chem 40:2184 –2190.
Naciff JM, Behbehani MM, Misawa H, Dedman JR (1999): Identification and transgenic analysis of a murine promoter that targets cholinergic neuron expression. J Neurochem 72:17–28.
Namavari M, Barrio JR, Toyokuni T, Gambhir SS, Cherry SR, Herschman HR, et al (2000): Synthesis of 8-[18F] fluorogua-nine derivatives: In vivo probes imaging gene expression with PET. Nucl Med Biol 27:157–162.
Neve KA, Cox BA, Henningsen RA, Spanoyannis A, Neve RL (1991): Pivotal role for aspartate-80 in the regulation of dopamine D2 receptor affinity for drugs and inhibition of adenylyl cyclase. Mol Pharmacol 39:733–739.
Nishiyama T, Kawamura Y, Kawamoto K, Matsumura H, Yamamoto N, Ito T, et al (1985): Antineoplastic effects of 5-fluorocytosine in combination with cytosine deaminase capsules. Cancer Res 45:1753–1761.
Pan D, Gambhir SS, Toyokuni T, Iyer MR, Acharya N, Phelps ME, Barrio JR (1998): Rapid synthesis of a 59-fluorinated oligodeoxy-nucleotide: A model antisense probe for use in imaging with positron emission tomography (PET). Bioorg
Med Chem Lett 8:1317–1320.
Pelletier S (1988): Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA.
Nature 334:320.
Phelps ME (1991): PET: A biological imaging technique.
Neu-rochem Res 16:929 –994.
Phelps ME, Schelbert HR, Mazziotta J, editors (1986):
Positron Emission Tomography and Autoradiography, Principles and Applications for the Brain and Heart. New
York: Raven.
Sachs AB, Sarnow P, Hentze MW (1997): Starting at the beginning, middle, and end: Translation initiation in eu-karyotes. Cell 89:831– 838.
Srinivasan A, Gambhir SS, Green LA, Cherry SR, Sharfstein S, Barrio JR, et al (1996): A PET reporter gene (PRG)/PET reporter probe (PRP) technology for repeatedly imaging gene expression in living animals. J Nucl Med 37:107P.
Sweeney TJ, Maila¨nder V, Tucker AA, Olomu AB, Zhang W, Cao Y, et al (1999): Visualizing the kinetics of tumor-cell clearance in living animals. Proc Natl Acad Sci U S A 96:12044 –12049.
Tachizawa H, Sudo K, Sasano H, Sano M (1981): Disposition and metabolism of timiperone in the rat, dog and monkey.
Drug Metab Dispos 9:442– 448.
Tavitian B, Terrazzino S, Kuhnast B, Marzabal S, Stettler O, Dolle F, et al (1998): In vivo imaging of oligonucleotides with positron emission tomography. Nat Med 4:467– 471. Tjuvajev JG, Avril N, Oku T, Sasajima T, Miyagawa T, Joshi R, et al (1998): Imaging herpes virus thymidine kinase gene transfer and expression by positron emission tomography.
Cancer Res 58:4333– 4341.
Tjuvajev JG, Callegari A, Lindsley L, Joshi R, Balatoni J, Finn R, et al (1999): A general approach to the non-invasive imaging of transgenes using cis-linked herpes simplex virus thymidine kinase. Neoplasia 1:315–320.
Tjuvajev JG, Finn R, Watanabe K, Joshi R, Oku T, Kennedy J, et al (1996): Noninvasive imaging of herpes virus thymidine kinase gene transfer and expression: A potential method for monitoring clinical gene therapy. Cancer Res 56:4087– 4095.
Tjuvajev JG, Stockhammer G, Desai R, Uehara H, Watanabe K, Gansbacher B, Blasberg RG (1995): Imaging the expression of transfected genes in vivo. Cancer Res 55: 6126 – 6132.
Weber S, Terstegge A, Herzon H, Reinartz R, Reinhart P, Rongen F, et al (1997): The design of an animal PET: Flexible geometry for achieving optimal spatial resolution or high sensitivity. IEEE Trans Med Imaging 16:684 – 689. Werstuck G, Green MR (1998): Controlling gene expression in living cells through small molecule-RNA interactions.
Sci-ence 282:296 –298.
Woodward R, Roley C, Daniell S, Naylor LH, Strange PG
(1996): Investigation of the role of conserved serine residues in the long form of the rat D2 dopamine receptor using site-directed mutagenesis. J Neurochem 66:394 – 402.
![Figure 4. 8-[18F]Fluoroganciclovir (FGCV) can be used as a positron emission tomography (PET) reporter probe to image herpessimplex virus 1 thymidine kinase (HSV1-TK) expression](https://thumb-ap.123doks.com/thumbv2/123dok/3143954.1383640/3.612.119.488.82.202/fluoroganciclovir-positron-emission-tomography-reporter-herpessimplex-thymidine-expression.webp)
![Figure 6. 3-(2�-[18F]Fluoroethyl)spiperone (FESP) can be used as a reporter probe to image dopamine 2 receptor (D2R) positronemission tomography (PET) reporter gene expression in living mice](https://thumb-ap.123doks.com/thumbv2/123dok/3143954.1383640/6.612.61.288.80.294/fluoroethyl-spiperone-dopamine-receptor-positronemission-tomography-reporter-expression.webp)
![Figure 7. Positron emission tomography (PET) image analysis using the 3-(2�measures dopamine 2 receptor (Dproduce varying levels of D-[18F]fluoroethyl)spiperone (FESP) reporter probe2R) reporter gene protein levels](https://thumb-ap.123doks.com/thumbv2/123dok/3143954.1383640/8.612.65.537.81.251/positron-emission-tomography-analysis-dproduce-fluoroethyl-spiperone-reporter.webp)