ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN1175-5334(online edition) Copyright © 2010 · Magnolia Press
Zootaxa 2576: 59–68 (2010)
www.mapress.com/zootaxa/
Article
A new endemic rock Gecko
Cnemaspis
Strauch 1887 (Squamata: Gekkonidae)
from Gunung Jerai, Kedah, northwestern Peninsular Malaysia
NORHAYATI AHMAD7, ANA-CAROLINE REMIGIO2, & LEE F. GREER21
Institute for Environment and Development (LESTARI), Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor Darul Ehsan, Malaysia. E-mail: [email protected]
2Department of Biology, La Sierra University, 4500 Riverwalk Parkway, Riverside, California, 92515-8247 USA 3School of Biological Sciences, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia
4Centre for Drug Research, Universiti Sains Malaysia, 11800 Minden, Penang, Malaysia 5
Department of Ecology and Evolutionary Biology, E145 Corson Hall, Cornell University, Ithaca, New York, 14853 USA
6 Department of Biology, Villanova University, 800 Lancaster Avenue, Villanova, Pennsylvania, 92108 USA
7School of Environment and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia 43600
Bangi, Selangor Darul Ehsan, Malaysia
8
Corresponding author
Abstract
A new species of endemic Cnemaspis is described from Gunung Jerai (also known as Kedah Peak) in the northwestern state of Kedah, Peninsular Malaysia. Cnemaspis harimau sp. nov. differs from all other Sundaland congeners except C. affinis, C. biocellata, C. kumpoli, C. mcguirei, C. pseudomcguirei, and C. shahruli in having a black shoulder patch with a white or yellow ocellus anteriorly located. It is most similar in appearance to its sister species, C. affinis, but differs by having a smaller maximum SVL of 40.7 mm vs. 50.8 mm; three vs. five postmentals; caudal tubercles encircling vs. not encircling tail; lateral caudal tubercles on anterior 25% of tail highly spinose and protruding vs. slightly spinose; and an overall higher degree of scale keeling (most prominent on the tail). The discovery of another montane endemic once again highlights the understudied nature of Peninsular Malaysia’s extensive mountain ranges.
Key words: Cnemaspis harimau, Gunung Jerai, herpetofauna, Kedah Peak, new species, taxonomy
Introduction
The Southeast Asian gekkonid genus Cnemaspis has seen a significant upsurge in diversity over the last seven years due to new discoveries and taxonomic revision. As a result, five new species have been described from Vietnam (Grismer & Ngo 2007; Grismer et al. 2010b), one from Cambodia (J. Grismer et al. 2010); one from Borneo (Grismer & Chan 2009), seven from Thailand (Grismer et al. 2010c, one from Laos (Grismer 2010); and 12 from Peninsular Malaysia and its associated archipelagos (Chan & Grismer, 2008; Das & Grismer 2003; Grismer & Chan, 2008; Grismer & Chan 2010; Grismer & Das, 2006; Grismer et al. 2008a,b, 2010a; Grismer et al. 2009). Prior to 2003, Peninsular Malaysia only had four known species of Cnemaspis [C. affinis
Grismer & Das, C. perhentianensis Grismer & Chan, C. baueri Das & Grismer, and C. limi Das & Grismer; lowland species: C. kendallii, C. flavigaster Chan & Grismer,C. kumpoli Taylor, and C. shahruli Grismer, Chan, Quah, Muin, Savage, Grismer, Ahmad, Greer & Remigio; upland species: C. mcguirei Grismer, Grismer, Wood & Chan, C. pseudomcguirei Grismer, Ahmad, Chan, Belabut, Muin, Wood & Grismer, known only from the Banjaran Bintang; C. affinis, known only from Penang Island; C. argus, known only from Gunung Lawit; and C. flavolineata, known only from the Banjaran Titiwangsa (see above references).
Gunung Jerai (=Jerai Mountain or Kedah Peak) is an isolated coastal massif reaching 1200 m a.s.l. in elevation located in the state of Kedah, northwestern Peninsular Malaysia. It is bordered by the Indian Ocean in the west and surrounded by notably flat plains which do not exceed 15 m a.s.l. in elevation with the nearest mountains lying some 32 km to the east or 35 km southwest on the island of Penang (WWF Malaysia 1977). Geologically, Gunung Jerai has been isolated from other mountain systems for its entire existence and existed as an island, numerous times in the past (Bradford 1972; WWF Malaysia 1977). Only four notable herpetological studies have ever been conducted on Gunung Jerai. The earliest collection came from Robinson and Kloss (1916) which reported three species of mammals, 13 reptiles and eight amphibians from Gunung Jerai and the adjacent towns of Alor Star and Gurun. Subsequently, anuran assemblages were studied by Sukumaran (2005) who reported a total of 25 species of frogs and discovered an undescribed lineage of
Odorrana from the upper elevations of the mountain which was later described as O. monjerai (Matsui & Jaafar 2006). Shahrul et al. (2006) then reported nine species of lizards, nine snakes and one turtle in a brief reptile survey.
During March 2010, field work on Gunung Jerai resulted in the discovery of a species of Cnemaspis from the granite boulder outcrops of Sungai Badak at 601 m elevation. This population possessed a unique suite of characters which could not be ascribed to any other known Southeast Asian Cnemaspis and is herein described as a new species based on its morphological diagnosibility under the general lineage concept of species (de Queiroz 1998, 1999).
Material and methods
The following measurements on the type series were taken with Mitutoyo dial calipers to the nearest 0.1 mm under a Nikon SMZ 1500 dissecting microscope on the left side of the body where appropriate: snout-vent length (SVL), taken from the tip of snout to the vent; tail length (TL), taken from the vent to the tip of the tail, original or regenerated; tail width (TW), taken at the base of the tail immediately posterior to the postcloacal swelling; forearm length (FL), taken on the dorsal surface from the posterior margin of the elbow while flexed 90º to the inflection of the flexed wrist; tibia length (TBL), taken on the ventral surface from the posterior surface of the knee while flexed 90º to the base of the heel; axilla to groin length (AG), taken from the posterior margin of the forelimb at its insertion point on the body to the anterior margin of the hind limb at its insertion point on the body; head length (HL), the distance from the posterior margin of the retroarticular process of the lower jaw to the tip of the snout; head width (HW), measured at the angle of the jaws; head depth (HD), the maximum height of head from the occiput to the throat; eye diameter (ED), the greatest horizontal diameter of the eye-ball; eye to ear distance (EE), measured from the anterior edge of the ear opening to the posterior edge of the eye-ball; eye to snout distance (ES), measured from anteriormost margin of the ball to the tip of snout; eye to nostril distance (EN), measured from the anterior margin of the eye-ball to the posterior margin of the external nares; inner orbital distance (IO), measured between the anterior edges of the orbit; ear length (EL), the greatest horizontal distance of the ear opening; and internarial distance (IN), measured between the nares across the rostrum.
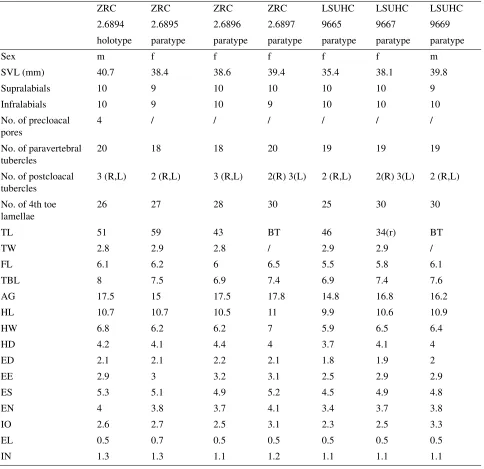
lamellae beneath the fourth toe counted from the base of the first phalanx to the claw; the total number of precloacal pores, their orientation, shape, and degree of separation; the degree and arrangement of body and tail tuberculation; the relative size and morphology of the subcaudal scales, subtibial scales, and submetatarsal scales beneath the first metatarsal; and the number of postcloacal tubercles on each side of the tail base. Longitudinal rows of caudal tubercles on the non-regenerated portion of the tail are quite variable between species and useful in differentiating several taxa. Up to five pairs of the following rows may be present in varying combinations: paravertebral row — the dorsal row adjacent to the middorsal, caudal furrow; dorsolateral row — the row between the paravertebral row and the lateral, caudal furrow on the dorsolateral margin of the tail; lateral row — the row immediately below the lateral, caudal furrow; and ventrolateral row — the row below the lateral row on the ventrolateral margin of the tail. When present, this row is usually restricted to the anterior 25% (or less) of the tail. Rarely there may be a row of tubercles within the lateral, caudal furrow. Measurements and character evaluations of the type series are presented in Table 1. Comparison of diagnostic characters of Cnemaspis within the shoulder-patch group is presented in Table 2. For comparisons with other Sundaland species, see Grismer et al. (2010b:Table 1).
Partial sequences of the mitochondrial gene encoding NADH dehydrogenase subunit 2 (ND2) were amplified and sequenced (Grismer et al. in prep.). Gene- specific pairwise distances between taxa were calculated with FastDist plus BioNJ / Neighbor (PHYLYP) 3.66 (Felsenstein 1989; Gascuel 1997).
The type series was deposited at the Zoological Reference Collection at the Raffles Museum of Biodiversity Research, National University of Singapore (ZRC) and La Sierra University Herpetological Collection (LSUHC).
Results
Systematics
Cnemaspis harimau sp. nov.
Tiger Rock Gecko Figure 2A, B
Holotype. Adult male (ZRC 2.6894) collected on 17 March 2010 by Chan Kin Onn, Lee Grismer, Jesse Grismer, Anna Savage, Shahrul Anuar, Mohd. Abdul Muin, and Evan Quah at 2030 hrs at 601 m a.s.l. from Sungai Badak (=Badak river), Gunung Jerai, Kedah, Peninsular Malaysia (N 05°48′59″, E 100°23′53″).
Paratypes. Collection locality and collector of the paratypes is the same as that of the holotype. The paratypes were collected between 2030 and 2130 hrs. ZRC 2.6897 (female; 16 March 2010); LSUHC 9669 (male; 17 March 2010); and ZRC 2.6895, 2.6896, LSUHC 9665, 9667 (female; 17 March 2010).
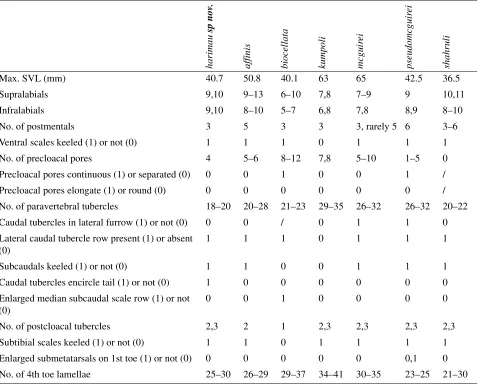
Diagnosis. Cnemaspis harimau differs from all other Southeast Asian congeners except C. affinis, C. biocellata, C. kumpoli, C. mcguirei, C. pseudomcguirei, and C. shahruli in having a black shoulder patch with a white or yellow ocellus anteriorly. It can be differentiated from the above-mentioned species in the unique combination of adult males reaching 40.7 mm SVL, females 39.4 mm SVL; nine or 10 supralabials, 9–10 infralabials; three postmentals; ventral scales keeled; four precloacal pores in males, arranged in a chevron, medially separated by one non-pore bearing scale; 18–20 paravertebral tubercles; 25–30 subdigital lamellae under the fourth toe; keeled subcaudals; enlarged, spinose lateral caudal tubercles on anterior 1/3 of tail; caudal tubercles encircling the tail, and males having an orange-colored gular region in life.
Description of holotype. Adult male; SVL 40.7 mm; head oblong in dorsal profile, moderate in size (HL/ SVL 0.26), somewhat narrow (HW/SVL 0.17), flat (HD/HL 0.39), distinct from neck; snout short (ES/HL 0.50), concave in lateral profile; postnasal region constricted medially; scales of rostrum low, rounded, juxtaposed, weakly keeled, larger than similarly shaped scales on occiput; weak, supraorbital ridges; frontonasal sulcus present, moderate; canthus rostralis smoothly rounded; eye large (ED/HL 0.20); extra-brillar fringe scales small in general but slightly larger anteriorly; pupil round; ear opening oval, taller than wide; rostral concave dorsally, dorsal 80% divided by longitudinal groove; rostral bordered posteriorly by two supranasals, a smaller, azygous postrostral, and nostrils; bordered laterally by first supralabials; 10 (R,L) raised supralabials of similar size, but smallest posteriorly; 11 (R,L) infralabials, decreasing gradually in size posteriorly; nostrils small, oblong, oriented dorsolaterally; bordered posteriorly by small, granular, postnasal scales; mental large, triangular, flat, extending to level of second infralabials, bordered postero-laterally by two enlarged, elongate postmentals of similar size which are separated by a single, small, azygous scale; gular scales smooth, rounded, juxtaposed; throat scales keeled.
webbing absent; toes increase in length from first to fourth with fourth and fifth nearly equal in length; 26 subdigital lamellae on fourth toe; dorsal, caudal scales arranged in segmented whorls, slightly larger than dorsal body scales; caudal scales low, keeled, juxtaposed anteriorly, raised and more keeled posteriorly; shallow, middorsal and lateral caudal furrow; subcaudal scales keeled, subimbricate anteriorly, degree of keeling and scale-overlap increase posteriorly; no median row of enlarged subcaudal scales; three longitudinal rows of tubercles on base of tail on either side of midline; single row of paravertebral tubercles throughout length of tail; single row of dorsolateral and lateral tubercles on anterior 25% of tail; no tubercles within lateral, caudal furrow; three postcloacal tubercles; tail approximately 125% of SVL.
FIGURE 2. A. Male holotype of Cnemaspis harimau sp. nov.(ZRC 2.6894). B. Female paratype of C. harimau sp. nov.
(LSUHC 9665). C. Adult male C. affinis from Penang Hill. D. Gunung Jerai (type locality of C. harimau sp. nov.), as viewed from Penang Hill (type locality of C. affinis).
Color in preservative. Dorsal base color including head grayish-brown; faint, post-orbital stripes are visible in dorsal profile; vertebral blotches indistinct; yellow bands faded to light gray; dorsal surfaces of limbs with irregular light and dark blotches; entire ventral surface whitish gray, orange coloration on gular region and throat undetectable; ventral surfaces of tail with dark stippling, corresponding to the caudal bands.
Variation and sexual dimorphism. Adult male, LSUHC 9669 has a darker base color and a distinct pre- and post-orbital stripe which connects to the occipital streaks with which it forms a continuous stripe; medial occipital spot present; paired paravertebral streaks are dark and more distinct than the holotype; precloacal pores were not assessable due to torn skin. Sexual dimorphism is distinct with females lacking precloacal pores, shoulder patches and orange coloration on the gular region and throat; other body markings are similar to males; the paratype females, ZRC 2.6896, 2.6897 and LSUHC 9667 have a darker base color; ZRC 2.6897 has a wide, distinct, continuous vertebral stripe extending from the base of the occiput to the tail base. It should be noted that base color of specimens, be it in life or in preservative, should not be taken into account when diagnosing this species because of their ability to substrate match. Other morphological variations are presented in Table 1. Values that were identical throughout the type series are not indicated.
TABLE 1. Morphological data taken for the type series. BT=broken tail; (r)= regenerated tail; /=unable to assess. See
Materials and methods for character abbreviations.
Supralabials 10 9 10 10 10 10 9
Infralabials 10 9 10 9 10 10 10
Distribution. Gunung Jerai, in the state of Kedah, Peninsular Malaysia.
Natural history. Sungai Badak was mostly dry during our visit and was a rocky stream bed, lined with dense vegetation and medium sized (0.5–2 m diameter) granite boulders. Lizards were observed on the boulders and at the base of tree trunks near the boulders during the day and night, although many more were seen at night. They were wary and when provoked, fled downwards to seek refuge in porous matrices formed by the dried expansive soil at the base of boulders.
Etymology. The specific epithet, harimau means tiger in the Malay language and is in reference to the yellow banding on the flanks of this species which are arbitrarily analogous to the banding on a tiger. Additionally, the year 2010 coincides with the Chinese zodiac year of the tiger.
FIGURE 3. Differences in subcaudal squamation with Cnemaspis harimau sp. nov. (top; holotype, ZRC 2.6894) having
larger lateral tubercles and stronger scale keeling as compared to C. affinis (bottom; LSUHC 6758).
from C. shahruli in having a maximum SVL of 40.7 mm vs. 36.5 mm; four vs. zero precloacal pores; and caudal tubercles encircle vs. do not encircle tail. Cnemaspis harimau is most similar in appearance to its sister species C. affinis, but differs by having a maximum SVL of 40.7 mm vs. 50.8 mm; 18–20 vs. 20–28 paravertebral tubercles; lateral postmentals separated by one vs. more than one (usually three) smaller, azygous scales; caudal tubercles encircle vs. do not encircle tail; lateral caudal tubercles on anterior 25% of tail highly spinose and protruding vs. slightly spinose; and an overall higher degree of scale keeling (most prominent on the tail). Differences between species within the shoulder patch group are summarized across Table 2.
TABLE 2. Diagnostic characters of species within the shoulder patch group.
Discussion
Preliminary molecular analysis show that Cnemaspis harimau (n=3) and C. affinis (n= 5) are sister species with a pairwise sequence divergences of 2.9–3.6% based on 749 base pairs on ND2 (Grismer et al. in prep.). This suggests that a common ancestor once ranged throughout lowland areas from at least Penang Island to Gunung Jerai at a time when Penang Island was connected to Peninsular Malaysia (Inger & Voris, 2001). A subsequent rise in sea levels separated Penang Island from the mainland and for a brief time, rendered Gunung Jerai as an island as well. This is likely to have resulted in the allopatric speciation event that gave rise to each species. Despite the relatively recent separation, both species have diverged enough to attain reciprocal
ha
Supralabials 9,10 9–13 6–10 7,8 7–9 9 10,11
Infralabials 9,10 8–10 5–7 6,8 7,8 8,9 8–10
No. of postmentals 3 5 3 3 3, rarely 5 6 3–6
Ventral scales keeled (1) or not (0) 1 1 1 0 1 1 1
No. of precloacal pores 4 5–6 8–12 7,8 5–10 1–5 0
Precloacal pores continuous (1) or separated (0) 0 0 1 0 0 1 /
Precloacal pores elongate (1) or round (0) 0 0 0 0 0 0 /
No. of paravertebral tubercles 18–20 20–28 21–23 29–35 26–32 26–32 20–22
Caudal tubercles in lateral furrow (1) or not (0) 0 0 / 0 1 1 0
Lateral caudal tubercle row present (1) or absent (0)
1 1 1 0 1 1 1
Subcaudals keeled (1) or not (0) 1 1 0 0 1 1 1
Caudal tubercles encircle tail (1) or not (0) 1 0 0 0 0 0 0
Enlarged median subcaudal scale row (1) or not (0)
0 0 1 0 0 0 0
No. of postcloacal tubercles 2,3 2 1 2,3 2,3 2,3 2,3
Subtibial scales keeled (1) or not (0) 1 1 0 1 1 1 1
Enlarged submetatarsals on 1st toe (1) or not (0) 0 0 0 0 0 0,1 0
it may range much lower, following a microhabitat of rocky stream corridors down the mountain. Cnemaspis harimau is likely to be endemic to Gunung Jerai being that Jerai is bordered by the Indian Ocean in west and paddy fields to the east.
Remarkably, Cnemaspis harimau is the first endemic reptile reported from this isolated mountain range. The ranid frog, Odorrana monjerai (Matsui & Jaafar 2006), which was previously thought to be endemic to Gunung Jerai, has also been found in Bukit Hijau, Kedah (Chan et al. 2010). This discovery adds to the growing number of upland endemics that have been found in the vastly unexplored mountain ranges of Peninsular Malaysia (Chan et al. 2009; Grismer 2007; Grismer et al. 2008b, 2009; Wood et al. 2009).
Acknowledgements
We thank the Kedah State Forestry Department for their permission to conduct research in Gunung Jerai Permanent Forest Reserve. This research was supported in part by USM Grant 815019. For assistance in the laboratory of Lee F. Greer, we thank Jennifer A. Burns, J. Rachel Salvador, Tzu Yi Liao, Keng Ti Liao, Neal Christopher, Caitlin Miller, Jennifer Gonzales, Valitsinee Pattanaprommas, Brittany Pick, Ramona Bahnam, Caitlin Hutchinson, Nathan Daramola, Fawaz Tarzi, Abraham Castillo, Kevin Bolanos, William Lee, William Angkadjaja, Whitney Ware, Wesley Marquis, and Rajiv Johnson. This research was supported in part by grants to LLG and LFG from the College of Arts and Sciences, La Sierra University, Riverside, California.
References
Bradford, E.F. (1972) The Geology and Mineral Resources of the Gunong Jerai Area, Kedah. Geological Survey Headquarters, Ipoh.
Chan, K.O., Daicus, B. & Norhayati, A. (2010) A Revised Checklist of the Amphibians of Peninsular Malaysia. Russian Journal of Herpetology, in press.
Chan, K.O. & Grismer, L.L. (2008) A new species of Cnemaspis Strauch 1887 (Squamata: Gekkonidae) from Selangor, Peninsular Malaysia.Zootaxa, 1877, 49–57.
Chan, K.O., Grismer, L.L., Norhayati, A. & Daicus, B. (2009) A new species of Gastrophrynoides (Anura: Microhylidae): an addition to a previously monotypic genus and a new genus for Peninsular Malaysia. Zootaxa, 2124, 63–68.
Das, I. & Grismer, L.L. (2003) Two new species of Cnemaspis Strauch, 1887 (Squamata: Gekkonidae) from the Seribuat Archipelago, Pahang and Johor States, West Malaysia. Herpetologica, 59, 544–552.
De Queiroz, K. (1998) The general lineage concept of species, species criteria, and the process of speciation: A conceptual unification and terminological recommendations. In: Howard, D.J. & Berlocher, S.H. (eds). Endless forms: Species and Speciation. Oxford: Oxford University Press, 57–75.
De Queiroz, K. (1999) The general lineage concept of species and the defining properties of the species category. In: Wilson, R.A. (ed.) Species: New Interdisciplinary Essays. Cambridge: The MIT Press, 49–89.
Felsenstein, J. (1989) PHYLIP - Phylogeny Inference Package (Version 3.2). Cladistics, 5, 164–166.
Gascuel, O. (1997) BIONJ: An improved version of the NJ algorithm based on a simple model of sequence data. Molecular Biology Evolution, 14(7), 685–95.
Grismer, J.L., Grismer, L.L. & Thou, C. (2010) A new species of Cnemaspis Strauch 1887 (Squamata: Gekkonidae) from southwestern Cambodia. Journal of Herpetology, 44(1), 28–36.
Grismer, L.L. (2007) A new species of small montane forest floor skink (Genus Spenomorphus Fitzinger 1843) from southern peninsular Malaysia. Herpetologica, 63, 544–551.
Grismer, L.L. (2010) The first record of the genus Cnemaspis Strauch (Squamata: Gekkonidae) from Laos with the description of a new species. Zootaxa, in press.
Grismer, L.L. & Chan, K.O. (2008) A new species of Cnemaspis Strauch 1887 (Squamata: Gekkonidae) from Pulau Perhentian Besar, Terengganu, Peninsular Malaysia. Zootaxa, 1771, 1–15.
Grismer, L.L. & Chan, K.O. (2009) A new species of karst dwelling Cnemaspis Strauch 1887 (Squamata: Gekkonidae) from Sarawak, Borneo. Zootaxa, 2246, 21–31.
Grismer, L.L. & Chan, K.O. (2010) Another new Rock Gecko (genus Cnemaspis Strauch 1887) from Pulau Langkawi, Kedah, Peninsular Malaysia. Zootaxa, 2419, 51–62.
Cnemaspis Strauch 1887) from the border region of Thailand and Peninsular Malaysia.Zootaxa, 1875, 51–68. Grismer, L.L., Chan, K.O., Quah, E., Muin, M.A., Savage, A.E., Grismer, J.L., Norhayati, A., Greer, L.F. III. &
Regemio, A-C. (2010a) Another new, diminutive Rock Gecko (Cnemaspis Strauch) from Peninsular Malaysia and a discussion of resource partitioning in sympatric, species pairs. Zootaxa, 2569, 55–66.
Grismer, L.L. & Das, I. (2006) A new species of gekkonid lizard of the genus Cnemaspis Strauch 1887 from Pulau Pemanggil, Johor, West Malaysia. Herpetological Natural History, 10, 1–7.
Grismer, L.L., Grismer, J.L., Wood, Jr., P.L. & Chan, K.O. (2008b) The distribution, taxonomy, and redescription of the geckos Cnemaspis affinis (Stoliczka 1887) and C. flavolineata (Nicholls 1949) with descriptions of a new montane species and two new lowland, karst-dwelling species from Peninsular Malaysia. Zootaxa, 1931, 1–24.
Grismer, L.L. & Ngo, V.T. (2007) Four new species of the gekkonid genus Cnemaspis Strauch 1887 (Reptilia: Squamata) from southern Vietnam. Herpetologica, 63,482–500.
Grismer, L.L., Ngo, V.T. & Grismer, J.L. (2010b) A colorful new species of insular rock gecko (Cnemaspis Strauch 1887) from southern Vietnam. Zootaxa, 2352, 46–58.
Grismer, L.L., Norhayati, A., Chan, K.O., Belabut, D., Muin, M.A., Wood, P.W. Jr. & Grismer, J.L. (2009) Two new diminutive species of Cnemaspis Strauch 1887 (Squamata: Gekkonidae) from Peninsular Malaysia. Zootaxa, 2019, 40–56.
Grismer, L.L., Sumontha, M., Cota, M., Grismer, J.L., Wood, P.L.Jr., Pauwels, O.S.G. & Kunya, K. (2010c) A revision and redescription of the rock gecko Cnemaspis siamensis (Taylor 1925) (Squamata: Gekkonidae) from Peninsular Thailand with descriptions of seven new species. Zootaxa, 2576, 1–55.
Inger, R.F. & Voris, H.K. (2001) The biogeographical relations of the frogs and snakes of Sundaland. Journal of Biogeography, 28, 863–891.
Matsui, M. & Jaafar, I. (2006) A New Cascade Frog of the Subgenus Odorrana from Peninsular Malaysia. Zoological Science, 23, 647–651.
Robinson, H.C. & Kloss, C.B. (1916) The Natural History of Kedah Peak. Journal of the Federated Malay States Museums, 7(2), 219–244.
Shahrul, A.M.S., Rayan, M., Nurul ‘Ain, D., Nor Zalipah, E., Mohd Yusof, M. & Ganesan, M.O. (2006) Brief notes on the reptilian fauna of Gunung Jerai. In: Shaharuddin, M.I., Wan-Yusoff, W.A., Jalil, M.S., Yusoff, M. & Latiff, A. (Eds), Hutan Simpan Gunung Jerai, Kedah: Pengurusan, persekitaran fizikal dan kepelbagaian biologi. Forestry Department of Peninsular Malaysia, Kuala Lumpur.
Sukumaran, J. (2005) Anuran Assemblages of Gunung Jerai, Kedah, Peninsular Malaysia. Unpublished M.Sc. Thesis, Faculty of Science, University of Malaya.
Wood, P.L.Jr., Grismer, J.L., Grismer, L.L., Norhayati, A., Chan, K.O. & Bauer, A. (2009) Two new montane species of Acanthosaura Gray, 1831 (Squamata: Agamidae) from Peninsular Malaysia. Zootaxa, 2012, 28–46.
WWF, Malaysia. (1977) Gunung Jerai State Park, Kedah: Its Natural Resources and its Future Role. World Wildlife Fund Malaysia, Kuala Lumpur.
Appendix. Species examined.
Cnemaspis affinis (Stoliczka 1870): LSUHC 6758, 6788, 6759, Penang Island.
Cnemaspis biocellata Grismer, Chan, Nurolhuda & Sumontha 2008: LSUHC 8787–8793, 8802–8805, Gua Kelam, Perlis.
Cnemaspis kumpoli Taylor 1963: LSUHC 8846–49, Wang Kelian, Perlis.