PENGARUH
PEJANTAN
TERHADAP
KERAGAMAN
DNA
MIKROSATELIT
DARI
LOKUS
CSN-3,
BM
143,
BM
415
DI
KROMOSOM
BTA-6
C. SUMANTRI1, A. ANGGRAENI2. dan A. FARAJALLAH3
1Departemen Ilmu Produksi Ternak, Fakultas Peternakan, IPB, Bogor 2Balai Penelitian Ternak, Jl. Veteran III PO Box 221 Ciawi, Bogor 16002
3Departemen Biologi, Fakultas Matematika dan IPA, IPB, Bogor
ABSTRAK
Tiga mikrosatelit DNA meliputi lokus CSN-3, BM 143 dan BM 415 yang terletak pada kromosom 6 (BTA-6) digunakan untuk mendeteksi pengaruh pejantan baik pada keragaman DNA mikrosatelit dan nilai pemuliaan (NP) produksi susu sapi Friesian Holstein (FH) di Balai Pembibitan Ternak Unggul (BPTU) Baturraden. Sejumlah kegiatan penelitian mencakup pengambilan sampel darah sapi laktasi, ekstraksi DNA (SAMBROOK et al., 1989) yang dimodifikasi, amplifikasi fragment DNA dengan PCR (polymerase chain reaction), genotiping dan identifikasi alel. Penghitungan frekuensi genotipe dan alel berdasarkan NEI. Proporsi genotipe dan alel dari ketiga lokus CSN-3, BM 143 dan BM 415 pada BTA-6 dipengaruhi oleh pejantan. Keragaman DNA mikrosatellit pada tiga lokus yang diamati memperlihatkan bahwa: lokus CSN 3 mempunyai genotipe AC pada klasifikasi NP tinggi (21,43%) lebih tinggi dari NP sedang (10,53%), lokus BM 143 mempunyai genotipe BB hanya pada NP tinggi (6,67%), dan lokus BM 415 mempunyai genotipe BB (20%) hanya pada NP rendah.
Kata kunci: DNA mikrosatelit, sapi FH, nilai pemuliaan
PENDAHULUAN
Pendekatan yang lebih sistematis untuk menemukan gen pengontrol produksi susu dapat dilakukan antara lain dengan membuat peta QTL (Quantitative Traits Loci) yakni dengan mempelajari keterkaitan gen (linkage gen) dan mengkoleksi penciri genetik pada keseluruhan kromosom (genom). Studi menggunakan desain grant-daughter dilapor-kan telah berhasil menemudilapor-kan lima kromosom yang mempengaruhi produksi susu, lemak, dan protein (GEORGESet al., 1995). Kemudian satu
QTL berhasil dipetakan pada kromosom 6 termasuk haplotipa kasein. Penelitian selanjutnya (ASHWELLet al., 1998) telah pula berhasil mengidentifikasi penciri genetik BM 415 dan BP 7 pada kromosom 6 dan BM 6425 pada kromosom 14 yang diidentifikasi memiliki pengaruh langsung pada persentase protein, sedangkan BM 719 pada kromosom 16 memiliki pengaruh langsung dengan lama sapi berproduksi susu. Mikrosatelit CSN3 kelompok gen kasein dilaporkan berhubungan dengan produksi susu, persentase lemak dan kg lemak (VELMALA et al., 1995). BM 143
berjarak 31 cM dari CSN3 (BISHOP et al., 1994). Lokus TGLA37 pada posisi 9 cM dari BM143 dan 22 cM dari CSN3 berpengaruh terhadap produksi susu persentase lemak dan protein (GEORGESet al., 1995 dan KUHNet al.,
1996).
Sejumlah penelitian awal yang ditujukan untuk mengetahui keragaman pada tingkat molekuler genetik telah dilakukan pada sapi FH di BPTU Baturraden. Sebagai contoh,
RAHMANIet al. (2004) melakukan pengamatan
terhadap keragaman satu gen (lokus) hormon pertumbuhan dan melaporkan ditemukan 4 alel dengan frekuensi A(0,21), B(0,09), C (0,04) dan D (0,66). SUMANTRI et al., 2004 yang mempelajari keragaman gen κ-kasein pada sapi FH di BPTU Baturraden melaporkan polimorfisme gen κ-kasein mempunyai frekuensi alel A (0,47) dan B (0,53) dengan komposisi genotipe AA (0,21), AB(0,53) dan BB (0,26). Dilaporkan lebih jauh bahwa genotipe BB sangat berpengaruh (P<0.01) terhadap kadar protein susu (SUMANTRIet al., 2005). Keragaman DNA mikrosatelit pada delapan lokus dari populasi sapi yang sama dilaporkan dalam studi selanjutnya (SUMANTRI
kedelapan lokus yang diamati. Dari 33 alel yang terdeteksi menunjukkan lima alel pada BM 143 dan BM 4208; empat alel pada BM415; CSN3; ETH225; BM1237 dan BM888 dan tiga alel pada BP 31. Ada 14 alel dari 33 alel terdeteksi kemungkinan alel spesifik sapi perah di Baturaden. Heterozigositas per lokus (h) paling rendah 0.6151 untuk lokus BM415 dan tertinggi 0.7301 untuk lokus BM888. Rataan heterozigositas (H) untuk kedelapan lokus sebesar 0.6768. Genotipe BB pada lokus BM 143 dan genotipe AC pada lokus CSN-3 berpengaruh nyata terhadap nilai pemuliaan produksi susu.
Salah satu penyebab tingginya keragaman genetik tersebut diperkirakan akibat dari pengaruh pajantan yang dipakai sebagai sumber genetik dalam perkawinan IB dari populasi sapi FH betina di BPTU Baturraden. Dengan demikian diperlukan penelitian lanjutan dengan memfokuskan pengamatan kontribusi pejantan (FH impor) terhadap proporsi genotipe dan nilai pemuliaan (NP) untuk produksi susu sapi FH betina laktasi di BPTU Baturraden.
METODE PENELITIAN Sapi FH pengamatan
Sejumlah 99 ekor sapi betina FH yang sedang berlaktasi di BPTU Baturaden diambil sampel darahnya untuk dianalisa genotipe DNA mikrosatelitnya. Sapi tersebut sudah dievaluasi NP produksi susunya selama pengamatan produksi 10 tahun (1988-1998) oleh Tim Peneliti Pusat Penelitian Peternakan
(ANGGRAENIet al., 2000).
Ekstrasi DNA
Setiap sampel darah dimasukkan ke dalam tabung falcon, disentrifugasi 3500 rpm selama 10 menit sehingga terbentuk tiga lapisan yaitu plasma darah, buffy coat (lapisan sel darah putih berinti) dan sel darah merah. Buffy coat dipindahkan ke tabung 1,5 ml yang akan digunakan sebagai sumber DNA. Ekstrasi
DNA dilakukan menurut SAMBROOK et al.,
(1989) yang di modifikasi.
Analisa PCR
Analisa PCR dilakukan dengan cara sebagai berikut: 2 μl 50 ng sampel DNA, 0,25
μl primer mikrosatelit dan kondisi suhu annealing PCR untuk setiap primer disajikan dalam (Tabel 1). 1,25 μl mM MgCl2, 1 μl 2 mM dNTPs dan 0,25 μl 4 unit Ampli Taq gold DNA polimerase dan ditambah 7,75 μl millique water steril sampai total volume 12,75
μl. Tabung tersebut dimasukkan ke dalam mesin PCR TaKaRa thermocycler MP4. Genotiping dilakukan dengan memisahkan produk PCR menggunakan teknik elektro-foresis gel poliakrilamid 5 - 8% dalam bufer 1xTBE (Tris 0.5 M, asam borat 0.65 M, EDTA 0.02M) yang kemudian diikuti dengan pewarnaan sensitif perak.
Analisa data
Frekuensi masing-masing alel setiap lokus mikrosatelit dihitung berdasarkan rumus NEI
(1987):
Keterangan:
Xi = frekuensi alel lokus ke-i
nij = jumlah individu untuk genotipe AiAj
nii = jumlah individu untuk genotip AiAi
N = julah sampel
HASIL DAN PEMBAHASAN Pengaruh pejantan terhadap proporsi genotipe dan alel
Selama kurun waktu 12 tahun (1988-2000) ada 21 pejantan yang semen bekunya dipakai untuk inseminasi buatan (IB) di BPTU Baturraden dan 10 diantaranya mempunyai anak lebih dari lima ekor (Tabel 1).
(
2
N
)
i
n
n
2
X
j ij ii i∑
≠
+
=
Tabel 1. Semen beku yang dipergunakan di BPTU Baturraden pada periode 1988-2000
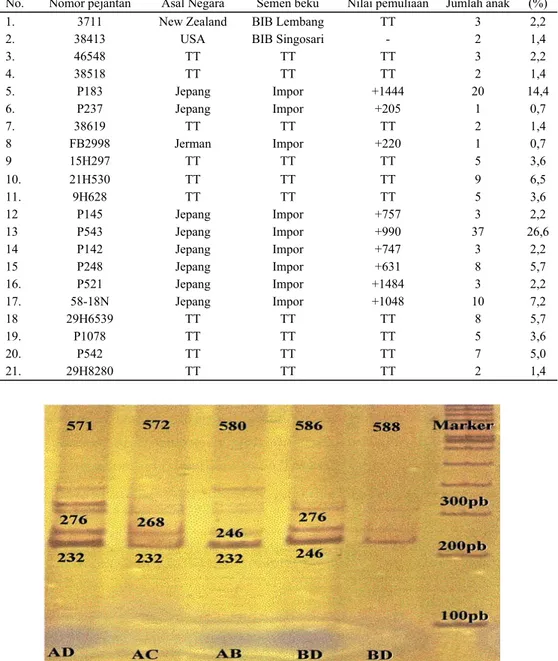
No. Nomor pejantan Asal Negara Semen beku Nilai pemuliaan Jumlah anak (%)
1. 3711 New Zealand BIB Lembang TT 3 2,2
2. 38413 USA BIB Singosari - 2 1,4
3. 46548 TT TT TT 3 2,2 4. 38518 TT TT TT 2 1,4 5. P183 Jepang Impor +1444 20 14,4 6. P237 Jepang Impor +205 1 0,7 7. 38619 TT TT TT 2 1,4 8 FB2998 Jerman Impor +220 1 0,7 9 15H297 TT TT TT 5 3,6 10. 21H530 TT TT TT 9 6,5 11. 9H628 TT TT TT 5 3,6 12 P145 Jepang Impor +757 3 2,2 13 P543 Jepang Impor +990 37 26,6 14 P142 Jepang Impor +747 3 2,2 15 P248 Jepang Impor +631 8 5,7 16. P521 Jepang Impor +1484 3 2,2 17. 58-18N Jepang Impor +1048 10 7,2 18 29H6539 TT TT TT 8 5,7 19. P1078 TT TT TT 5 3,6 20. P542 TT TT TT 7 5,0 21. 29H8280 TT TT TT 2 1,4
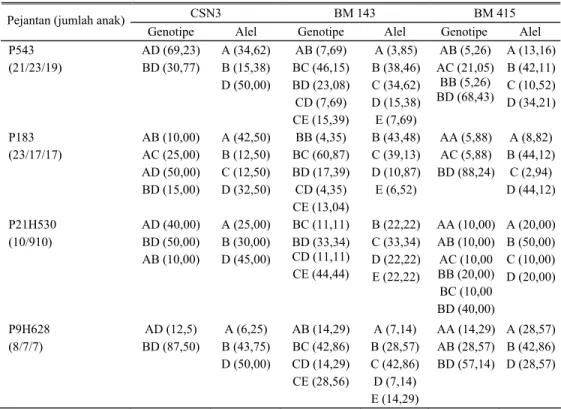
Gambar 1. Pola elektroforesis lokus CSN3 individu No 571 (genotipe AD), No 572 (genotipe AC), No 580 (genotipe AB), No 586 dan No 588 (genotipe BD)
Jumlah pejantan yang dipakai pada penelitian ini ada empat ekor meliputi: P.543 dengan nilai pemuliaan (NP +990) dan proporsi pemakaian 26,6% (mempunyai 37
14,4% (mempunyai anak 20 ekor), tetapi yang 2 ekor lainnya P21H530 dan P9H628 tidak tersedia informasi baik nilai pemuliaan maupun proporsi pemakaiannya (3,6-6,5%).
AD, No 572 bergenotipe AC, No 580 bergenotipe AB, No 586 dan No 588 bergenotipe BD. Untuk lokus BM 143 dan BM 415 dapat dilihat pada SUMANTRI et al. (2006).
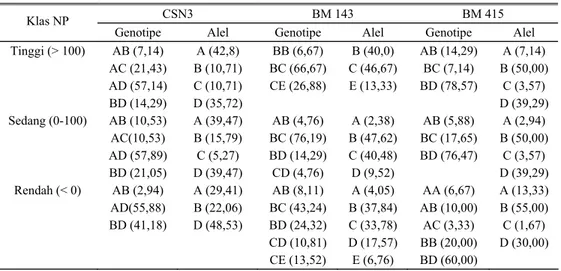
Proporsi genotipe dan alel dari setiap lokus pada kelompok tetua jantan yang berbeda diperlihatkan pada Tabel 2. Pada lokus CSN 3 genotipe AD dan BD umum ditemukan pada semua pejantan dengan proporsi yang bervariasi. Genotipe AC (25%) dan AB (10%) hanya ditemukan pada pejantan P 183. Alel A, B dan D umum ditemukan pada semua pejantan, kecuali alel C (12,5%) hanya ditemukan pada pejantan P 183. Pada lokus BM 143 genotipe BC, CD dan CE umum ditemukan pada semua pejantan dengan proporsi bervariasi. Genotipe BD ditemukan di tiga pejantan lainnya kecuali pada pejantan P9H628. Genotipe AB hanya ditemukan pada pejantan P543 (7,69%) dan pejantan P9H628 (14,29%), genotipe BB (4,35%) hanya ditemukan pada pejantan P183. Alel B,C,D, dan E umum ditemukan di semua pejantan,
kecuali alel A hanya ditemukan pada pejantan P543 (3,85%) dan pejantan P9H628 (7,14%). Pada lokus BM 415 genotipe BD umum ditemukan di semua pejantan dengan proporsi bervariasi, genotipe AA ditemukan di tiga pejantan kecuali P 543, AB umum ditemukan kecuali di pejantan P 183 dan AC umum ditemukan kecuali pada pejantan P9H628 dan BB ditemukan hanya pada pejantan P21H530 (20,00%) dan P543 (5,26%). Alel A, B, dan D umum ditemukan pada semua pejantan, alel C ditemukan pada tiga pejantan kecuali pada P9H628.
Hasil tersebut menunjukkan terdapat variasi yang diakibatkan oleh penggunaan pejantan.
SUMANTRI et al. (2006) melaporkan nilai
heterozigositas per lokus (ĥ) untuk BM 143 (0,6756) lebih tinggi dari CSN3 (0,6572) dan BM 415 (0,6151). Nilai heterozigositas merupakan cara yang paling akurat untuk mengukur variasi genetik suatu populasi (NEI dan KUMAR 2000).
Tabel 2. Proporsi genotipe dan frekuensi alel dari DNA mikrosatelit lokus CSN3, BM 143 dan BM 415 (BTA-6) berdasarkan pejantan berbeda
CSN3 BM 143 BM 415
Pejantan (jumlah anak)
Genotipe Alel Genotipe Alel Genotipe Alel
P543 (21/23/19) AD (69,23) BD (30,77) A (34,62) B (15,38) D (50,00) AB (7,69) BC (46,15) BD (23,08) CD (7,69) CE (15,39) A (3,85) B (38,46) C (34,62) D (15,38) E (7,69) AB (5,26) AC (21,05) BB (5,26) BD (68,43) A (13,16) B (42,11) C (10,52) D (34,21) P183 (23/17/17) AB (10,00) AC (25,00) AD (50,00) BD (15,00) A (42,50) B (12,50) C (12,50) D (32,50) BB (4,35) BC (60,87) BD (17,39) CD (4,35) CE (13,04) B (43,48) C (39,13) D (10,87) E (6,52) AA (5,88) AC (5,88) BD (88,24) A (8,82) B (44,12) C (2,94) D (44,12) P21H530 (10/910) AD (40,00) BD (50,00) AB (10,00) A (25,00) B (30,00) D (45,00) BC (11,11) BD (33,34) CD (11,11) CE (44,44) B (22,22) C (33,34) D (22,22) E (22,22) AA (10,00) AB (10,00) AC (10,00 BB (20,00) BC (10,00 BD (40,00) A (20,00) B (50,00) C (10,00) D (20,00) P9H628 (8/7/7) AD (12,5) BD (87,50) A (6,25) B (43,75) D (50,00) AB (14,29) BC (42,86) CD (14,29) CE (28,56) A (7,14) B (28,57) C (42,86) D (7,14) E (14,29) AA (14,29) AB (28,57) BD (57,14) A (28,57) B (42,86) D (28,57)
Hubungan genotipe mikrosatelit DNA dengan produksi susu
Laporan sejumlah penelitian yang banyak dilakukan pada populasi sapi perah di daerah iklim temperate telah mengindikasikan adanya pengaruh yang jelas dari genotipe mikrosatelit DNA terhadap produksi dan kualitas susu yang dihasilkan. Sebagai contoh RON et al.
(1994) yang melakukan pengamatan pada Sapi FH Israel dengan menggunakan 10 mikrosatelit DNA melaporkan mikrosatelit D21S4 berpengaruh sangat nyata (P<0,01) terhadap produksi susu dan protein. Dengan menggunakan 159 mikrosatelit DNA yang tersebar pada genom sapi perah, GEORGES et al. (1995) mengidentifikasi bahwasanya QTL produksi susu terletak menyebar pada lima
kromosom yaitu kromosom no 1, 6, 9, 10 dan 20, namun hasil pengamatan QTL memberikan pengaruh sangat bervariasi pada produksi susu, persentase protein dan lemak. Studi lain
(ASWHELL et al., 1997) membuktikan QTL
untuk produksi susu terletak pada kromosom nomor 21 dengan mikrosatelit BM103 berpengaruh nyata (P<0,01) pada persentase lemak dan protein, sedangkan mikrosatelit BM 3413 berpengaruh nyata (P<0,01) pada produksi lemak dan protein. Selanjutnya studi lain pada mikrosatelit TGLA 153, AGLA 29, dan BM 5004 yang terletak pada kromosom nomor 20 menunjukkan adanya korelasi yang erat dengan persentase protein dan persentase lemak (ARRANZ et al. (1998).
Tabel 3. Proporsi genotipe dan frekuensi alel lokus BTA 6 berdasarkan klasifikasi nilai pemuliaan tertaksir EBV berbeda
CSN3 BM 143 BM 415
Klas NP
Genotipe Alel Genotipe Alel Genotipe Alel Tinggi (> 100) AB (7,14) AC (21,43) AD (57,14) BD (14,29) A (42,8) B (10,71) C (10,71) D (35,72) BB (6,67) BC (66,67) CE (26,88) B (40,0) C (46,67) E (13,33) AB (14,29) BC (7,14) BD (78,57) A (7,14) B (50,00) C (3,57) D (39,29) Sedang (0-100) AB (10,53) AC(10,53) AD (57,89) BD (21,05) A (39,47) B (15,79) C (5,27) D (39,47) AB (4,76) BC (76,19) BD (14,29) CD (4,76) A (2,38) B (47,62) C (40,48) D (9,52) AB (5,88) BC (17,65) BD (76,47) A (2,94) B (50,00) C (3,57) D (39,29) Rendah (< 0) AB (2,94) AD(55,88) BD (41,18) A (29,41) B (22,06) D (48,53) AB (8,11) BC (43,24) BD (24,32) CD (10,81) CE (13,52) A (4,05) B (37,84) C (33,78) D (17,57) E (6,76) AA (6,67) AB (10,00) AC (3,33) BB (20,00) BD (60,00) A (13,33) B (55,00) C (1,67) D (30,00)
Tabel 3 memperlihatkan genotipe AC pada lokus CSN 3 klasifikasi NP tinggi (21,43%) lebih tinggi dari NP sedang (10,53%) dan tidak ditemukan pada NP rendah. Genotipe AB, AD dan BD umum ditemukan pada semua klasifikasi dengan proporsi bervariasi. Alel A,B dan D umum ditemukan dengan proporsi bervariasi, kecuali alel C hanya ditemukan pada klasifikasi NP tinggi (10,71%) dan klasifikasi NP sedang (5,27%). Genotipe BC pada lokus BM 143 umum ditemukan pada semua klasifikasi dengan proporsi bervariasi,
bervariasi dan genotipe CE ditemukan pada klasifikasi EBV tinggi (26,88%) dan klasifkasi NP rendah (13,52%) dan BB hanya muncul pada klasifikasi NP tinggi (6,67%).
Alel B dan C umum ditemukan pada semua klasifikasi dengan proporsi bervariasi, alel A dan D hanya ditemukan pada klasifikasi sedang dan rendah dan alel E hanya ditemukan pada klasifikasi tinggi (13,33%) dan klasifikasi rendah (6,76%). Hal ini memperkuat hasil penelitian sebelumnya yang dilaporkan oleh
antara genotipe BB pada lokus BM 143 dan genotipe AC pada lokus CSN-3 dengan nilai pemuliaan produksi susu. Genotipe AB dan BD umum ditemukan pada semua klasifikasi dengan proporsi bervariasi, genotipe BC hanya ditemukan pada klasifikasi tinggi (7,14%) dan klasifikasi sedang (17,65%) dan genotipe AC (3,33%) dan BB (20%) hanya ditemukan pada klasifikasi NP rendah. Alel A,B, C dan D umum ditemukan pada semua klasifikasi dengan proporsi yang bervariasi.
KESIMPULAN
Keragamnan genotipe dan alel dari ketiga lokus CSN-3, BM 143 dan BM 415 pada BTA-6 dari polupasi sapi FH BPTU Baturraden dipengaruhi oleh pejantan IB dengan proporsi keragaman dari ketiga lokus tersebut ditemuka bervariasi antara pejantan. Keragaman DNA mikrosatelit pada tiga lokus yang diamati memperlihatkan bahwa: lokus CSN 3 mempunyai genotipe AC dengan proporsi relatif tinggi untuk NP produksi susu tinggi, sedangkan lokus BM 143 mempunyai genotipe BB dengan proporsi relatif tinggi untuk NP produksi susu tinggi (6,67%), sebaliknya lokus BM 415 mempunyai genotipe BB hanya pada NP rendah.
DAFTAR PUSTAKA
ANGGRAENI, A. 2000. Identifikasi Keunggulan Genetik Produksi Susu Sapi Perah Fries Holland sebagai Penghasil Sapi Perah Bibit. Laporan Balitnak Tahun 1999 dan 2000. Bogor.
ARRANZ, J.J. COPPIETERS, W. BERZI, P. CAMBISANO, N. GRISART, B. KARIM,L. MARCQ, F. MOREAU, L. MEZER, C., RIQUET, J. SIMON, P., VANMANSHOVEN, P. WAGENAAR, D. and GEORGES, M. 1998. A QTL Affecting Milk Yield and Composition Maps to Bovine Chromosome 20: a Confirmation. Animal Genetics. 28:107-115.
ASWELL, M.S., REXROAD. JR, C.E., MILLER, R.H., and VANRADEN, P.M. 1997. Detection of Loci Affecting Production and Health Traits in an Elite US Holstein Population Using Microsatellite Markers. Animal Genetics. 28:216-222.
BAWDEN, W.S. and NICHOLAS, K.R. 1999. Molecular Genetics of Milk Production: In The Genetics of Cattle Edited: R. FRIES and A. RUVINSKY. CAB International. New York. USA.
BISHOP, MD., KAPPES, SM., KEELE JW, STONE RT, SUNDEN SLF, HAWKINS GA, TOLDO, SS, FRIES, R., GROSZ, MD., YOO J and BEATTIE CW. 1994. A Genetic Linkage Map for Cattle. Genetics 136:619-639.
GEORGES, M., NIELSON, D., and MACKINNON M. 1995. Mapping Quantitative Trait Loci Controlling Milk Production in Dairy Cattle by Exploiting Progeny Testing. Genetics. 139. 907-920.
KUN, CH., FREYER, G., WEIKARD, R. GOLDAMMER, T and SCHWERIN, M. 1999. Detection of QTL for Milk Production Traits in Cattle by Application of Specifically Developed Marker Map of BTA6. Animal Genetics. 30:333-340. NEI. M. 1987. Molecular Evolutionary Genetics.
Columbia University Press. New York. NEI, M, and KUMAR. S. 2000. Molecular Evolution
and Phylogenetics. Oxford University Press. Inc. New York.
RAHMANI, N., MULADNO, dan SUMANTRI, C. 2004. Analisis Polimorfisme Gen Bovine growth hormone (bGH) pada Sapi Perah Friesian Holstein di BPTU Baturraden. Prosiding Seminar Nasional Teknologi Peternakan dan Veteriner. Bogor, 4-5 Agustus. 2004.
RON, M., BAND, M., YANAI, A. and WELLER J.J. 1994. Mapping Quantitative Trait Loci with DNA Microsatellite in a Commercial Dairy Cattle Population. Animal Genetics 25: 259-294.
SAMBROOK, J., FRITSCH, E.F., and MANIATIS T. 1989. Molecular Cloning Laboratory Manual 3rd Ed. Cold Spring Harbour Lab. Press. New
York.
SUMANTRI, C., ANGGRAENI, A., MAHESWARI, R.R.A., DIWYANTO, K., FARAJALLAH, A. dan BRAHMANTIYO, B. 2004. Frekuensi Gen Kappa Kasein (κ-kasein) pada Sapi Perah FH berdasarkan Produksi Susu di BPTU Baturraden.Prosiding Seminar Nasional Teknologi Peternakan dan Veteriner. Bogor, 4-5 Agustus. 2004.
SUMANTRI, C., MAHESWARI, R.R.A., ANGGRAENI, A., DIWYANTO, K., dan FARAJALLAH, A. 2005. Pengaruh Genotipe Kappa Kasein (κ-kasein) terhadap Kualitas Susu pada Sapi Perah FH di BPTU Baturraden.Prosiding Seminar Nasional
Teknologi Peternakan dan Veteriner. Bogor, 12-13 September. 2005.
SUMNATRI, C., PERWITASARI, D., FARAJALLAH, A. dan A. ANGGRAENI. 2006. Keragaman DNA Mikrosatelit Sapi Fries Holstein (FH) di Balai Pembibitan Ternak Unggul (BPTU) Sapi Perah Baturraden. Prosiding Seminar Nasional Teknologi Peternakan dan Veteriner. Bogor, 5-6 September 2006.
VELMALA, R., VILKKI,J., ELO,K. DEKONING, D.J. and MAKI-TANILA, A. V. 1999. A Search for Quantitative Trait Loci for Milk Production Traits on Chromosome 6 in Finnish Ayrshire Cattle. Animal Genetics. 30:136-143.