TINJAUAN PUSTAKA
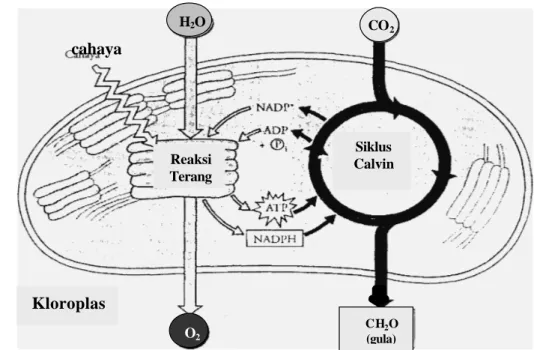
Karakteristik yang paling penting pada tumbuhan adalah kemampuan nya untuk memanen energi dari matahari yang digunakan untuk mengubah CO2 udara menjadi molekul organik yang lebih komplek. Proses ini dinamakan fotosintesis yang meliputi reaksi oksidasi dan reduksi. Proses keseluruhan adalah oksidasi air (pemindahan elektron disertai pelepasan O2 sebagai hasil samping) pada reaksi terang dan reduksi CO2 pada siklus Calvin untuk membentuk senyawa organik, misalnya karbohidrat (Gambar 1). Faktor-faktor yang berpengaruh terhadap fotosintesis antara lain adalah ketersediaan CO2, H2O, cahaya, hara, suhu, umur dan genetik (Salisbury dan Ross 1992, Campbell et al. 2002).
Gambar 1 Gambaran umum fotosintesis: kerjasama reaksi terang dan s i k l u s Calvin (Campbell et al. 2002).
Daun dan Kloroplas
Daun merupakan organ utama tempat berlangsungnya fotosintesis pada tumbuhan. Struktur daun dipengaruhi oleh cahaya yang akan berpengaruh pada luas dan tebal daun, kerapatan trikoma dan kandungan antosianin pada vakuola sel epidermis (Bolhar-Nordenkampf dan Draxler, 1993). Secara anatomi, daun biasanya terdiri dari lapisan epidermis atas dan bawah yang melapisi sel-sel mesofil (beberapa lapis sel palisade dan sel bunga karang) (Hopkins dan Huner 2004). Lapisan palisade dan bentuk sel-sel
Reaksi Terang Kloroplas Siklus Calvin cahaya CO2 H2O O2 CH2O (gula)
mesofil dapat berubah sesuai kondisi cahaya, yang menyebabkan tanaman menjadi efisien dalam menyimpan energi cahaya untuk perkembangannya. Peran yang kontras antara sel palisade dan sel bunga karang, yaitu sel palisade dapat menyebabkan cahaya lewat dan sel bunga karang menangkap cahaya sebanyak mungkin, menyebabkan absorbsi cahaya yang lebih seragam di dalam daun. (Taiz dan Zeiger 2002 ).
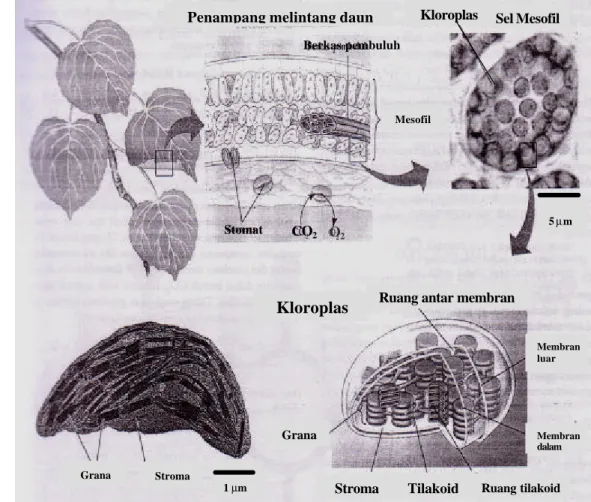
Gambar 2 Anatomi daun, stomata dan kloroplas (Campbell et al. 2002).
Pada daun juga terdapat organ -organ tambahan yang berperan dalam dua proses penting yaitu fotosintesis dan transpirasi, melalui pertukaran gas pada daun dengan lingkungannya (Woodward 1998, Croxdale 2000 ). Tanaman mengoptimalkan pertukaran gasnya dengan beradaptasi secara long-term (misalnya melalui kerapatan stomata) dan short-term (pembukaan dan penutupan stomata) (Schluter 2003).
Stomat a
Grana Stroma
Kloroplas
Kloroplas Sel Mesofil
Penampang melintang daun
Stroma Grana
Tilakoid Ruang tilakoid Membran dalam Membran luar Ruang antar membran
Mesofil Berkas pembuluh
CO2 O2
5 µm
Dalam sel-sel mesofil terdapat organel sel yang berfungsi untuk melakukan proses fotosintesis yang disebut kloroplas (Gambar 2). Aneka bentuk dan ukuran kloroplas ditemukan pada berbagai tumbuhan. Kloroplas berasal dari proplastid yang membelah pada saat embrio berkembang, dan menjadi kloroplas ketika daun dan batang terbentuk. Kloroplas muda juga aktif membelah, khususnya bila organ yang mengandung kloroplas terkena cahaya (Salisbury dan Ross 1992).
Sel mesofil umumnya memiliki kurang lebih 30-40 kloroplas , berukuran 2-4 x 4-7 µm (Campbell et al., 2002). Kloroplas diselubungi membran ganda untuk mengatur lalu lintas molekul keluar masuk kloroplas. Di dalam kloroplas dijumpai bahan amorf, lir -gel, dan kaya akan enzim yang disebut stroma. Di dalam stroma terdapat tilakoid (kantung) yang mengandung pigmen. Pada membran tilakoid ini energi cahaya digunakan untuk mengoksidasi H2O untuk membentuk ATP d an NADPH, energi yang diperlukan dalam Siklus Calvin untuk mengubah CO2 menjadi karbohidrat. Pada bagian tertentu terdapat tumpukan tilakoid yang disebut grana. Rongga yang ada dalam tilakoid dinamakan lumen, yang berisi air dan garam terlarut yang juga berperan dalam fotosintesis (Salisburry dan Ross 1992, Newcomb 1997, Becker et al. 2000, Campbell et al. 2002, Taiz dan Zeiger 2002).
Kloroplas berisi DNA dan RNA sehingga banyak protein kloroplas merupakan produk transkripsi dan translasi dalam kloroplas. Protein yang lain dikode oleh DNA inti, disintesis oleh ribosom dalam sitoplasma (Newcomb 1997, Campbell et al. 2002, Buchanan et al. 2000).
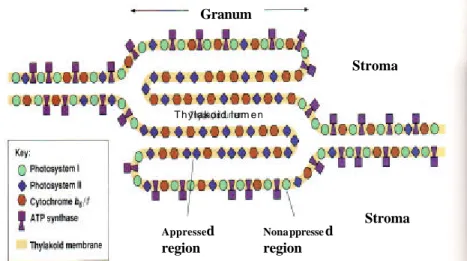
Pada membran tilakoid terdapat berbagai macam protein esensial untuk fotosintesis. Fotosistem II (PSII), Komple k cyt b6-f, Fotosistem I (PSI) dan ATP synthase yang merupakan protein integral dan tersisip dalam lipid bilayer dari membran. PS II dan protein transport elektron berlokasi terutama pada lamela grana. PS I dan protein transfer elektron serta enzim yang mengkatalisis pembentukan ATP (ATP Synthase) ditemukan dalam lamela stroma dan bagian pinggir dari lamela grana. Komplek cyt b6-f yang menghubungkan kedua fotosistem terdistribusi secara merata (Becker et al.
2000, Taiz dan Zeiger 2002, Hopkins dan Huner 2004). Organisasi komplek protein pada membran tilakoid disajikan pada Gambar 3.
Gambar 3 Organisasi komplek protein pada membran tilakoid (Becker et al. 2000). Cahaya
Cahaya sangat besar peranannya dalam proses fisiologis , seperti fotosintesis, respirasi, pertumbuhan dan perkembangan, penutupan dan pembukaan stomata, berbagai pergerakan tanaman, dan perkecambahan (Taiz dan Zeiger 2002) melalui sinyal transduki pada 4 tipe fotoreseptor, fotoreseptor cahaya biru dan 3 fotoreseptor cahaya merah (phytochrom, protochlorophyllide reductase dan chlorophyll) (Mullet 1997)
Cahaya merupakan bentuk energi yang dikenal sebagai energi elektromagnetik, yang juga disebut radiasi yang bergerak dalam bentuk gelombang. Jarak antara puncak -puncak gelombang disebut panjang gelombang dan yang paling penting bagi kehidupan berkisar antara panjang gelombang 380-750 nm, yang dikenal sebagai cahaya tampak . Cahaya ini dapat dideteksi mata manusia sebagai bermacam-macam warna (Campbell et al. 2002).
Cahaya tersusun atas partikel-partikel yang d isebut foton, memiliki jumlah energi tetap yang berbanding terbalik dengan panjang gelombang cahaya sehingga semakin pendek panjang gelombang, semakin tinggi energinya. Energi cahaya pada panjang gelombang 400-700 nm merupakan
Stroma Stroma Granum Appressed region Nonappressed region Thylakoid lumen
panjang gelombang paling aktif dalam fotosintesis (Becker et al. 2000, Taiz dan Zeiger 2002).
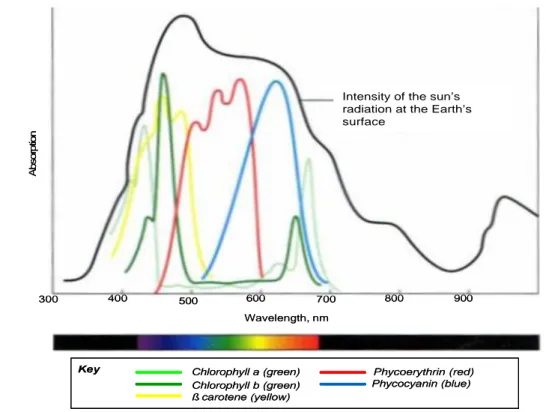
Bila cahaya mengenai materi, cahaya dapat dipantulkan, diteruskan (ditransmisi), atau diserap (diabsorpsi). Bahan -bahan yang menyerap cahaya-tamp ak pada tumbuhan disebut pigmen. Pigmen yang berbeda akan menyerap panjang gelombang yang berbeda, dan panjang gelombang yang diserap akan menghilang (Campbell et al. 2002; Hopkins dan Huner 2004 ). Spektrum cahaya yang diserap oleh berbagai pigmen disajikan pada Gambar 4.
Gambar 4 Spektrum cahaya yang diserap oleh berbagai pigmen (Becker et al. 2000).
Pigmen Fotosintetik
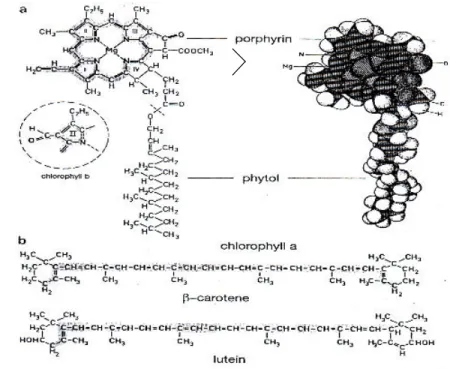
Klorofil dan karoten merupakan pigmen fotosintetik pada tumbuhan. Klorofil merupakan pigmen utama dalam fotosintesis, turunan δ-aminolevulinic acid (ALA) yang dibentuk di plastid (Sandmann dan Scherr 1998). Struktur klorofil terdiri dari cincin porfirin, dengan atom magnesium sebagai pusat dan ekor hidrokarbon (phytol) yang berinteraksi dengan daerah hidrofobik protein pada membran tilakoid (Gambar 5). Bagian cincin berisi elektron yang tidak terikat secara ketat dan merupakan bagian molekul yang terlibat dalam transisi
300 400 500 600 700 800 900
Wavelength, nm
Intensity of the sun’s radiation at the Earth’s surface Absorption Chlorophyll a (green) Chlorophyll b (green) ß carotene (yellow) Phycoerythrin (red) Phycocyanin (blue) Key 300 400 500 600 700 800 900 Wavelength, nm
Intensity of the sun’s radiation at the Earth’s surface Absorption Chlorophyll a (green) Chlorophyll b (green) ß carotene (yellow) Phycoerythrin (red) Phycocyanin (blue) Key
elektronik dan reaksi redoks (Campbell et al. 2002, Taiz dan Zeiger 2002, Hopkins dan Huner 2004).
Gambar 5 a. Struktur klorofil a b. rumus molekul β-carotene dan lutein (Campbell et al. 2002).
Klorofil a (C55H72O5N4Mg) berperan sebagai penangkap cahaya dan klorofil a tertentu (P680 dan P700) berperan-serta secara langsung dalam reaksi terang, mengubah energi matahari menjadi energi kimiawi. Klorofil b (C55H70O6N4Mg) hampir identik dengan klorofil a, tetapi perbedaan struktural yang kecil di antara keduanya cukup untuk membuat kedua pigmen tersebut mempunyai spektra absorpsi yang berbeda, sehingga warnanyapun juga berbeda. Klorofil a berwarna biru-hijau sementara klorofil b berwarna kuning-hijau. Klorofil b berfungsi sebagai pigmen pemanen cahaya yang d ibentuk dari klorofil a melalui oksidasi methyl (-CH3) pada tetraphyrrole cincin B menjadi bentuk formil (-CHO) setelah terjadi akumulasi klorofil a (Ito et al. 1996) oleh enzim oxigenase (Sandmann dan Scheer 1998). Bila klorofil b disintesa melebihi kebutuhan, klorofil b akan dikonversi ke klorofil a dan akan terikat ke komplek protein klorofil a (Ito et al. 1996). Bila klorofil b tidak dapat dikonversi ke klorofil a, klorofil b akan dilepaskan dari apoprotein LHCII (light-harvesting complexes II) dan ditimbun pada membran tilakoid sebagai klorofil bebas.
Klorofil bebas ini beracun bagi kloroplas karena membentuk oksigen radikal, yang merusak kloroplas (Ito et al. 1994).
Pigmen aksesoris selain klorofil b adalah karotenoid (karoten dan xanthofil) , merupakan senyawa hidrokarbon yang mempunyai warna berbagai campuran kuning dan jingga, berfungsi terutama dalam fotoproteksi dengan menyerap dan melepaskan energi cahaya yang berlebihan, yang jika tidak dilepas akan merusak klorofil. Karotenoid merupakan pigmen yang menangkap cahaya pada kisaran panjang gelombang 445 nm, energi yang diserap akan disalurkan ke pusat reaksi melalui klorofil b (Sandmann dan Scheer 1998). Pigmen-pigmen asesori ini sebagian besar ditemukan di kompleks pemanen cahaya (LHC) pada PSII (Buchanan et al. 2000, Hopkins dan Huner 2004).
Fotosistem sebagai Komplek Pemanen Cahaya
Pada membran tilakoid terdapat dua jenis fotosistem yang bekerja secara bersama pada reaksi terang fotosintesis yaitu fotosistem I (PSI) dan fotosistem II (PSII). Masing-masing memiliki pusat reaksi dan aseptor elektron primer tertentu yang berdekatan dengan molekul klorofil a yang menjadi pusat reaksi. Klorofil a pusat-reaksi fotosistem I adalah P700, menyerap cahaya pada panjang gelombang 700 nm (bagian spektrum yang sangat merah). Klorofil a pusat-reaksi fotosistem II adalah P680, spektrum absorpsinya memiliki puncak pada 680 nm (juga bagian spektrum merah).
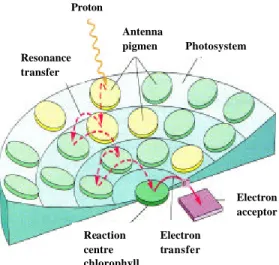
Fotosistem memiliki kompleks antena pengumpul-cahaya (LHC) yang berfungsi untuk mengirim energi secara efisien ke pusat reaksi dimana mereka berasosiasi. Ukuran sistem antena berbeda-bed a. Jumlah dan keragaman molekul pigmen membuat fotosistem dapat mengumpulkan cahaya pada permukaan yang lebih luas dan spektrum yang lebih lebar daripada yang dapat dikumpulkan molekul pigmen tunggal. Ketika setiap molekul antena menyerap foton, energinya disalurkan dari molekul pigmen ke molekul pigmen lain d idekatnya yang energinya lebih rendah hingga energi itu menemukan klorofil a yang berfungsi sebagai pusat reaksi (Gambar 6). Kira-kira 95-99% foton yang diserap pigmen antena ditransfer ke pusat reaksi dengan absorbsi maksimum ke arah warna merah. Transfer elektron yang digerakkan cahaya matahari dari klorofil ke
akseptor elektron primer merupakan langkah pertama reaksi terang (Becker et al. 2000, Taiz dan Zeiger 2002, Hopkins dan Huner 2004).
Gambar 6 Transfer energi ke pusat reaksi pada suatu fotosistem (Becker et al. 2000).
Mekanisme adaptasi tanaman terhadap naungan
Intensitas cahaya rendah menyebabkan proses metabolisme terganggu, yang diakibatkan oleh adanya penurunan laju fotosintesis dan sintesis karbohidrat. Pengaruh tercepat dari cekaman cahaya rendah ialah terhadap penurunan kandungan karbohidrat, terutama fruktosa dan sukrosa, yang diikuti dengan berbagai perubahan metabolisme pada tanaman. Pada kebanyakan tanaman, kemampuan tanaman dalam mengatasi cekaman naungan adalah tergantung pada kemampuannya dalam melanjutkan fotosintesis dalam kondisi defisit cahaya (Levitt 1980).
Mohr dan Schopfer (1995) menjelaskan bahwa adaptasi terhadap naungan pada dasarnya dapat melalui dua cara, yaitu melalui: 1) peningkatan luas daun sebagai cara mengurangi penggunaan metabolit, dan 2) mengurangi jumlah cahaya yang ditransmisikan dan direfleksikan. Sedang Levitt (1980) menyatakan bahwa adaptasi terhadap naungan dicapai melalui: 1) mekanisme penghindaran (avoidance) yang berkaitan dengan respon perubahan anatomi dan morfologi daun untuk fotosintesis yang efisien dan 2) mekanisme toleran (tolerance) yang berkaitan dengan penurunan titik kompensasi cahaya serta respirasi yang efisien.
Uji cepat ruang gelap merupakan cara yang efektif untuk menyeleksi plasma nutfah toleran pada intensitas cahaya rendah. Genotipe toleran yang
Resonance transfer Proton Antenna pigmen Reaction centre chlorophyll Electron transfer Electron acceptor Photosystem
efisien respirasinya mampu bertahan hidup sampai akhir penelitian. Nilai kesesuaian uji tersebut pada padi gogo 93,3% dengan percobaan naungan paranet dan 56% dengan hasil uji lapang di bawah tegakan karet (Sopandie et al. 2003b). Naungan 50% pada padi gogo menyebabkan perubahan morfologi, anatomi dan agronomi baik pada genotipe yang toleran maupun yang peka dan mekanisme efisiensi penangkapan cahaya dilakukan dengan penambahan tinggi tanaman, memperluas permukaan daun, penurunan ketebalan daun dan lapisan mesofil (Sopandie et al. 2003 a).
Pada kedelai, seleksi plasma nutfah pada 70 genotipe dengan menggunakan naungan paranet 50% menghasilkan 29 genotipe toleran, 25 moderat dan 16 peka. Hasil verifikasi selanjutnya diperoleh 13 genotipe yang konsisten toleran dari 16 genotipe yang diuji, antara lain Ceneng dan Pangrango. Dan satu genotipe yang konsisten peka dari 5 genotipe yang diuji, yaitu Godek A (Elfarisna 2000). Naungan pada kedelai menyebabkan kenaikkan kandungan klorofil daun, kadar nitrogen daun, bobot 100 biji, penurunan jumlah polong dan biji per tanaman dan produksi biji kering (Sunarlim, 1985). Reduksi cahaya menjadi 40% sejak perkecambahan pada kedelai menurunkan jumlah buku, cabang, diameter cabang, jumlah polong, dan hasil biji (Baharsjah 1980).
Perubahan pada Kloroplas
Cahaya mempengaruhi orientasi kloroplas dalam sel daun. Pada intensitas cahaya rendah, kloroplas akan mengumpul pada kedua sisi dinding sel terdekat dan terjauh dari cahaya. (Mohr dan Schopfer 1995, Taiz dan Zeiger 2002).
Intensitas cahaya rendah meningkatkan jumlah kloroplas per sel, volume kloroplas dan membran tylakoid serta grana (stack granum). Aktivitas fotokimia pada pusat reaksi II dan jumlah cytochrome f menurun, sedang jumlah LHC meningkat, berhubungan dengan keberadaan pigmen antena yang lebih banyak (Tanaka et al. 1998). Perbedaan antara kloroplas cahaya dan naungan adalah ukuran volume grana. Kloroplas cahaya hanya mempunyai grana 2-5 membran sedang kloroplas naungan dapat mempunyai 10-20 kali lipatnya yang berkorelasi dengan ratio klorofil a/b dan PSII/PSI, untuk meningkatkan penangkapan dan penyerapan cahaya (Critchley 1999). Penurunan nisbah klorofil a/b pada intensitas cahaya rendah disebabkan peningkatan klorofil b, yang berkaitan
dengan peningkatan protein klorofil a/b pada LHCII. Membesarnya antena untuk fotosistem II ini akan mempertinggi efisiensi pemanenan cahaya. Walaupun kandungan klorofil tinggi, rendahnya laju fotosintesis sering dihubungkan dengan tingginya resistensi stomata dan rendahnya aktivitas Ribulose bifosfat (RuBP). Selain itu, walaupun kandungan klorofil meningkat namun terjadi penurunan jumlah klorofil per luas area daun karena daun menjadi lebih tipis (Mohr dan Schopfer 1995).
Mekanisme adaptasi terhadap intensitas cahaya rendah pada kedelai menurut Khumaida (2002) melalui peningkatan kandungan klorofil yang terjadi pada semua genotipe kedelai yang diberi perlakuan naungan. Kandungan klorofil a dan b sama-sama meningkat, namun proporsi peningkatan terjadi lebih banyak pada klorofil b, sehingga menurunkan rasio klorofil a/b. Perlakuan gelap menyebabkan gangguan perkembangan membran tilakoid kedelai toleran maupun peka naungan. Sedang perlakuan naungan 50% tidak menghambat perkembangan membran tilakoid pada genotipe toleran seperti Pangrango dan B613, tetapi menghambat pada genotipe peka Godek. Pada kondisi naungan 50% kloroplas genotipe toleran mempunyai volume grana dan butir pati lebih banyak dibandingkan genotipe peka.
Antosianin
Selain klorofil yang terdapat dalam kloroplas, pada tanaman juga terdapat pigmen non kloroplas yaitu antosianin (anthos=bunga; kyanos=biru tua). Antosianin merupakan produk metabolisme sekunder yang termasuk golongan flavonoid dan dikenal sebagai pigmen yang menyebabkan warna merah jambu, ungu dan biru. Antosianin dibentuk dari asam amino Phenylalanine melalui lintasan sikimat di sitoplasma dan ditimbun dalam vakuola sel parenkim dewasa (Noh dan Spalding 1998, Buchanan et al. 2000). Lintasan pembentukan antosianin disajikan pada gambar 7. Antosianin merupakan bentuk glikosida (turunan gula) dari berbagai jenis antosianidin yang dibentuk dari dihidroflavonol. Keberadaan gula (aglikon) pada antosianin sangat meningkatkan kelarutannya dalam air (Salisbury dan Roos 1995). PAL (Phenylalanine Ammonia Lyase) merupakan enzim kunci dalam metabolisme, aktivitasnya meningkat seiring dengan umur daun dan berhubungan dengan proses penuaan (Buchanan et al.
2000). Pembentukan antosianin dipengaruhi cahaya (ultra violet dan cahaya tampak) juga oleh stress hara (terutama kekurangan nitrogen dan fosfor), kekeringan serta suhu rendah (Hoch et al. 2003).
Gambar 7 Lintasan pembentukan antosianin (modifikasi dari Noh dan Spalding 1998 serta Buchanan et al. 2000). Enzim yang terlibat adalah PAL (Phenylalanine Ammonia Lyase); CHS (Chalcone synthase); CHI (Chalcone Isomerase); F3H (Flavanone-3-hydroxylase); dan DR (Dihydroflavonol 4-reductase) .
Woodall dan Stewart (1998) menyimpulkan dari hasil beberapa penelitian bahwa antosianin mempunyai distribusi yang luas pada daun, dapat ditemukan pada sel epidermis bagian atas atau bawah, atau sel-sel mesofil dan biasanya mempunyai absorbsi pada panjang gelombang 270-290 nm (UV) dan 500-550 nm (visible spectrum). Acylasi dengan asam organik aromatik meningkatkan penyerapan pada daerah UV di panjang gelombang 310-320 nm. Hal ini menunjukkan fungsi antosianin yang mencegah kerusakan akibat radiasi UV. Hopkins dan Huner (2004) juga menjelaskan bahwa antosianin menyerap cahaya
Shikimat Siklus Calvin Phenylalanine Cinnamate Naringenin chalcone Flavan-3,4-diols Anthocyanin 4-Coumarate Metabolisme Primer Metabolisme Sekunder 4-Coumaryl-CoA 3 malonyl-CoA Naringenin 3-hydroxyflavones PAL1 PAL2 PAL3 CHS CHI F3H DFR
antara 475 nm dan 560 nm dan mentransmisikan cahaya biru dan merah. Sebagaimana flavonoid, antosianin menyerap UV-B sehingga diduga fungsinya untuk mencegah kerusakan jaringan daun yang disebabkan oleh radiasi ultra violet seperti kerusakan asam nukleat, protein dan apparatus fotosintesis yang akan mengurangi biomass.
Antosianin dapat diekstrak dalam larutan yang sedikit asam. Warna antosianin peka terhadap pH : merah (asam), violet (netral) dan biru (basa) dan umumnya bermuatan positif sehingga bergerak menuju katode saat elektroforesis. Perubahan warna ini terjadi akibat hidroksilasi atau metilasi pada aglikon dan reaksi tersebut dapat balik (Salisbury dan Roos 1995, Hopkins dan Huner 2004 ).