Review
Novel components of the plant cytoskeleton: a beginning to plant
‘cytomics’
Eric Davies
a,*, Bratislav Stankovic
b, Kishu Azama
c, Koichi Shibata
c,
Shunnosuke Abe
caBotany Department,North Carolina State Uni6ersity,Raleigh,NC27695-7612,USA bWCSAR,Uni6ersity of Wisconsin,Madison,WI53706,USA
cLaboratory of Molecular Cell Biology,Faculty of Agriculture,Ehime Uni6ersity,Matsuyama, Japan
Received 16 May 2000; received in revised form 30 July 2000; accepted 1 August 2000
Abstract
The bulk of our knowledge concerning the plant cytoskeleton has come primarily from the use of techniques and probes derived from animal research. However, in comparison with animal tissues, relatively few plant cytoskeleton proteins have been identified. We presume this is not because the plant cytoskeleton is really made up of such few proteins, but rather that only rarely have attempts been made to identify plant-specific cytoskeleton proteins, using plant-specific methods. Here we outline methods that we have developed both for the isolation and identification of novel cytoskeleton proteins as well as for the visualization of novel filamentous structures in plant cells, and we describe several novel cytoskeleton proteins and two novel cytoskeleton structures, ‘nanofilaments’ and ‘nanotubules’. We postulate that use of such approaches will lead to a rapid expansion of our knowledge of the plant cytoskeleton. © 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Cytomics/proteomics; Cytoskeleton-associated proteins; Cytoskeleton-bound polysomes; 4 nm nanofilaments; 12 nm nanotubules; Plant-specific methods
www.elsevier.com/locate/plantsci
1. Prelude
Over the last 30 years, the concept of the cytosol has shifted from that of a watery bagful of sub-strates and enzymes undergoing reactions solely in solution, to a highly-ordered network of macro-molecular complexes entailing reactions based on solid-state physics [1 – 3]. This implies both cytoso-lic compartmentation [4] and a highly ordered milieu [5,6]. However, in order to understand cell function, this milieu must first be understood. Put another way, the role(s) of individual proteins, including enzymes [7] and macromolecular
com-plexes depend(s) on their subcellular location(s). The single most important macromolecular net-work in eukaryotic cells is the cytoskeleton, and more needs to be understood about both its indi-vidual components and its macromolecular archi-tecture. It orchestrates a large array of activities, providing spatial and temporal coherence for both materials and information, i.e. signaling [8 – 14]. We anticipate that the wealth of information aris-ing from the disciplines of: genomics, i.e. the entire nucleotide sequence of an organism; transcrip-tomics, i.e. the identification of all currently tran-scribed genes; and proteomics, i.e. the 3-D structure, function and (static) location of all cel-lular proteins [15] will lead to a new discipline, which we call ‘cytomics’. By ‘cytomics’ we mean the changes in sub-cellular location and function
* Corresponding author. Tel.:+1-919-5131901; fax:+ 1-919-515-3436.
E-mail address:eric –[email protected] (E. Davies).
E.Da6ies et al./Plant Science160 (2001) 185 – 196
186
of macromolecules (especially proteins) and
macromolecular complexes over space and time in individual cells within intact organisms. Since the plant cytoskeleton is crucial for dynamic cell func-tion, understanding it is a pre-requisite to under-standing plant cytomics.
2. Introduction
We are aware of three fundamentally different, yet complementary, approaches to deciphering the molecular components of the plant cytoskeleton. The first employs probes (proteins, antibodies and/
or nucleic acids), including actin and tubulin affinity chromatography [14] originally derived from animal research, and is in many ways the most successful, especially if the animal probes bind to or react with something in the plant. Further, since it is based on the already-known, this approach tends to furnish the most readily-ac-cepted results. It will not, however, identify proteins that are novel to plants and runs the risk that negative results can be taken to ‘prove’ that the plant homolog is really absent, while positive results ‘prove’ that a genuine plant homolog does exist. The bulk of plant cytoskeleton research find-ings has come from this approach and examples are too numerous to mention.
The second approach involves determining the location of a specific, known protein, generally by
workers already knowledgeable about that
protein. This approach tends to be slower, insofar as workers specializing on one particular protein need to learn techniques in a different field (cy-toskeleton isolation) or by collaborating with
col-leagues who already have the requisite
specialization in cytoskeleton techniques. How-ever, the success of this approach is bolstered by the sense of ownership that the workers feel for their pet protein. If the protein is novel to plants, this approach can (obviously) identify novel plant cytoskeleton proteins, and the results will tend to be accepted, at least in part because the workers are already recognized experts on that protein. However, if the cytoskeleton location is novel to plants (and especially if the findings contradict findings with animals), the results are less likely to be accepted. Examples of proteins found to be cytoskeleton-associated using this approach
in-clude enzymes involved in phosphoinositide
metabolism, earlier thought to be cytosolic and/or
membrane-associated [16], the NPA-binding
protein, earlier thought to be membrane-associ-ated [17] and sucrose synthase, earlier thought to be cytosolic [18].
The third approach involves isolating the cy-toskeleton in as intact and un-denatured state as possible, and identifying the associated proteins. However, the global nature of this approach and the frequent lack of expertise of the workers on a protein they happen to identify as cytoskeleton-as-sociated contributes to frequent rejection of these results (especially by experts who have a vested interest in that particular protein) as not being totally ‘proven’. Nevertheless, this is the approach we have adopted, since we feel it will furnish a wealth of novel plant cytoskeleton proteins, will lead to the greatest advances in plant cytomics, and is the one on which this review (personal perspective) concentrates.
As we have stated earlier [19,20], the biggest obstacles to the successful isolation of components of the plant cytosol are the tough cell wall and the vacuole. The tough cell wall demands the use of considerable force to disrupt it, while this, in turn, disrupts the vacuole, causing its contents to be released into the cytoplasm. The vacuole is often 10-fold the volume of the cytoplasm and usually contains deleterious materials such as acid pH and hydrolytic enzymes. These tend to destroy the delicate contents of the cytoplasm, rendering it difficult to isolate and identify them. There are, however, at least three different methods to cir-cumvent these problems: (a) employing cells de-void of a cell wall (or converting the plant cell to a protoplast by removing the cell wall), thereby permitting gentler homogenization; (b) using cells devoid of a dominant vacuole; and (c) designing buffers which protect the cytoplasmic contents from the deleterious contents of the vacuole.
3. Methodological approaches
3.1. Use of protoplasts
cause changes in cytoskeleton structure, hence they may not furnish a native cytoskeleton [19]. Nevertheless, these methods, and modifications thereof provided the basis for the isolation and identification of novel cytoskeleton proteins, in-cluding those involved in inositol phosphate sig-naling [22] and, most recently, the isolation of a 65-kDa microtubule-associated protein from car-rots, apparently responsible for the regular align-ment of cortical microtubules [23]. The protoplast approach has been modified recently by Anders-land et al. [24], who isolated the cytoskeleton from cotton fiber cytoplasts (cytoplasmic droplets de-void of a nucleus). These methods provide
sub-stantial amounts of cytoskeleton material
uncontaminated with nuclear material, but they furnish a cytoskeleton with relatively few proteins. Furthermore, the normal problems engendered by protoplast isolation are exacerbated by the cyto-plast isolation techniques, which cause additional disassembly of some cytoskeleton arrays (Anders-land et al., 1998). We are not aware of any work-ers who have used wall-less plant cells (such as liquid endosperm of tomato) for cytoskeleton isolation.
3.2. Use of non-6acuolate cells
We are also unaware of any workers who have adopted the second method, i.e. deliberately using non-vacuolate cells as starting material. Non-vac-uolated cells in plants generally occur in merister-matic regions, such as stem tips and root tips, but these regions are small and it would be time-con-suming to isolate amounts sufficient for
biochemi-cal analysis of the plant cytoskeleton.
Nevertheless, the endosperm tissue that we and others have used to study the role of the cytoskele-ton in protein synthesis and protein body forma-tion (reviewed in [25]) happens to be almost devoid of vacuoles and has relatively weak cell walls.
3.3. Use of specifically-designed cytoskeleton-stabilizing buffers
This method (using buffers which minimize the deleterious effects of the vacuole) is more likely to provide valuable results from a wide range of plant organs and tissues and is the method we originally adopted in our efforts to identify
puta-tive polyribosome-cytoskeleton interactions in
plants [26,27]. We designed a cytoskeleton stabiliz-ing buffer (CSB) by evaluatstabiliz-ing the protective role of each component and found that the most cru-cial were: ‘gentle’ buffers (such as HEPES) at low ionic strength buffers, the absence of Tris (a severe cytoskeleton-disrupting agent); low concentrations of monovalent cations (especially K+); the
pres-ence of at least 10 mM Mg2+; the absence of
Ca2+ (and the inclusion of EGTA) to inhibit a
calcium-activated protease; and the addition of protease inhibitors such as PMSF to inhibit other proteases [19,26].
We have used these methods to show that polysomes do, indeed, associate with the cy-toskeleton in pea stems [27] and pea roots [28],
and with a cytoskeleton-membrane complex
(protein body) in corn endosperm [29 – 32]; re-viewed in Stankovic et al. [25]. Larkins and co-workers have extended these studies on the role of the cytoskeleton in protein body biogenesis and function to show that the eukaryotic elongation factor (now called eEF1A) is in very close associa-tion with the cytoskeleton-protein body complex in corn endosperm [33,34]. Furthermore, Okita and co-workers have furthered our understanding of cytoskeleton-protein body interactions using rice endosperm to show that the legume-type protein bodies that store globulins are derived from the ER indirectly by fusion of Golgi vesicles into storage vacuoles, while cereal-type protein bodies that store prolamines are directly derived from the ER [35]. This group has also shown that localization of prolamine mRNA depends on a cytoskeleton-associated binding site [36].
4. Recent findings
4.1. No6el proteins of the plant cytoskeleton
In our protocol described in detail earlier [19], we isolate the cytoskeleton as a large, cytoskele-ton-membrane-polysome (CMP) complex, remove membranes with detergent to furnish a
cytoskele-ton/polysome complex, depolymerize the
cy-toskeleton in high ionic strength (Tris) buffers, remove the polysomes by centrifugation, isolate the solubilized (putative cytoskeleton) proteins by heparin affinity chromatography followed by 1D
E.Da6ies et al./Plant Science160 (2001) 185 – 196
188
spots, microsequence them and make degenerate primers to probe our cDNA expression library. Using either the expressed proteins or purified native proteins we generate antibodies to probe protein gel blots and perform in vitro and/or situ localization.
Working primarily with pea stems, we have used
these methods to examine the cytoskeleton
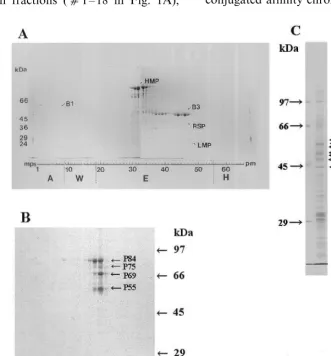
proteins released from the cytoskeleton-mem-brane-polysome complex, eluted from a heparin column and subjected to 1D-PAGE and several bands are labeled (Fig. 1A). Since this method did not furnish sufficient protein for analysis of the
unbound protein fractions (c1 – 18 in Fig. 1A),
these fractions were pooled, dialyzed against 1/10 CSB, re-applied to a heparin column, and eluted in KOAc (Fig. 1B). Further, since some
cytoskele-ton material was present in a lower speed (4000×
g) pellet, this was also analyzed (Fig. 1C). In
addition to the proteins labeled based on their location in a stained gel, we have also identified others using antibodies, including actin, a-tubulin,
b-tubulin, and eEF1A. In addition, we have
la-beled another protein, streptavidin-binding protein (SBP) based on its ability to bind streptavidin (and thus show up on protein gel blots probed with any antibody). This SBP was further purified by avidin conjugated affinity chromatography and eluted by
6 M guanidine. The results of these analyses are summarized in Table 1 which presents the protein’s label (as in Fig. 1), molecular mass, pI, number of spots, identity, and method of identity (e.g. cDNA or amino acid sequence).
We have identified at least two groups of proteins: those generally thought to be associated with mitochondria; and those involved in transla-tion (Table 1, Fig. 1). Proteins we have identified in the cytoskeletal fraction that are generally thought to be mitochondrial-associated proteins include: B1 (a hydrogen-transporting ATPase);
SBP (the biotin-containing b subunit of
methyl-crotonyl CoA: carbon dioxide ligase); and SBPa
(the a-subunit of the same enzyme). However,
these proteins may not be restricted to the mito-chondrion. There is at least one mitochondrial protein (citrate synthase) that can exist in the cytoplasm [37] and even form 14 nm (cytoskele-ton-like) filaments bearing eEF1a [37], while an-other (aconitase) changes its sub-cellular location from primarily mitochondrial to primarily cy-toskeletal during development [39].
Most interesting (at least from our perspective that the cytoskeleton plays a major role in
tether-ing, targeting, transporting and translating
mRNA) are those proteins involved in some
as-pect of RNA/protein metabolism. B2 (Fig. 1,
Table 1) is an omnipotent suppressor homolog, involved in translating non-sense codons and ap-parently containing a novel RNA binding domain [40]. B3 (Fig. 1, Table 1) is an apyrase or NTPase and might be involved in mRNA transport along the cytoskeleton [41]. RSP (ribosome-sedimenting protein) is a histone H1 homolog thought to be involved in ribosome attachment to the cytoskele-ton [42]. P55 (Fig. 1, Table 1) is a DnaK type molecular chaperone involved in transporting proteins into the chloroplast [43]. P84 (Fig. 1, Table 1) is an eEF2 homolog, involved in polypep-tide elongation, and has been shown to be a cytoskeleton-associated protein in animal cells [44]. Interestingly, DnaJ, another molecular chap-erone is also thought to be a cytoskeleton-associ-ated protein involved in gravity responses [45]. Since DnaK and DnaJ act together in protein import into the chloroplast [43] and BiP is impli-cated in transport of protein across the ER [46], we suspect that the cytoskeleton plays a major role not only in translation, but also post-translational modification and targeting.
The approach we have developed [19] has re-cently been adopted by Okita and coworkers [47] to analyze some of the proteins associated with the rice cytoskeleton-protein body. Partial peptide se-quences indicated that several ‘expected’ proteins, including those involved in translation (poly(A)-binding protein, ribosomal proteins S3, S4, S 17, L 13 and L27), and those associated with the animal cytoskeleton (titin) were present in the (biochemi-cal) cytoskeleton fraction. In addition, several ‘un-expected’ proteins, including those thought to be nuclear-associated (histone H3, transcription fac-tor Sox-19) and a phosphoglucomutase were also found in the cytoskeleton fraction. One significant difference between our results and those of Okita’s group is that we use more prolonged ultracentrifu-gation to remove polyribosomes, ribosomes, sub-units and sub-ribosomal particles, and thus find less integral ribosomal proteins as a ‘contaminant’ of the cytoskeleton fraction. Despite these differ-ences, it is increasingly apparent from our work as well as Okita’s that several proteins involved in translation are found closely associated with the plant cytoskeleton.
E
.
Da
6
ies
et
al
.
/
Plant
Science
160
(2001)
185
–
196
190
Table 1
The identities of several proteins isolated from the cytoskeleton derived from the cytoskeleton-membrane-polysome complex of pea stemsa
Heparin
Label given Mass (kDa) pI Spots (c) Identity Methodc Previous localization
affinityb
Hydrogen transporting ATPase
55 5.7 1 B150 Pa, C1 Mitochondria
B1
P Ribosome, cytoplasm
Omnipotent suppressor protein
B2 51 5.9 1 NA
Apyrase, NTPase, ectophosphatase
49 5.0–7.0 5 670 Pb, C2, E Nucleus, cell wall, ribosomes,
B3
CSK
NA MethylcrotonylCoA:CO2 ligase (beta subunit) P, C Mitochondria
6.5–6.7
SBP 78 3
P, C Mitochondria
(Alpha subunit) NA
NA NA
SBPa 60
Ribosome-sedimenting protein (atypical histone H1
42.5 NA NA 690 Pc, C3 Previously unknown
RSP
homolog)
P
P55 55 NA NA 20 DnaK type chaperone homolog Membranes, cytosol
P Ribosome, cytoskeleton
84 7.4–7.8 5 20 eEF2 homolog
P84
a 1GenBank No. AB003549;2GenBank No. AB022319; and3GenBank No. AB003549.aacc No. BAA20135;bacc No. BAA75506; andcacc No. BAA78535.
bmM KOAc needed to remove from column.
example is the mRNA binding protein mrnp 41 [50]. In addition to citrate synthase [37,38] men-tioned above, another mitochondrial-associated protein found in other locations is aconitase [51], whose relative distribution between the cytoskele-ton and other locations can change during devel-opment [39].
Recent examples from plants of proteins with multiple locations (including the cytoskeleton) have been shown for apyrase (NTPase, ectophos-phatase), which can locate to the nucleus and the cytoskeleton, where it might be involved in mRNA transport [41], as well as to the extra-cellu-lar matrix, where it might be involved in phos-phate reclamation [52] and xenobiotic resistance [53]; importin-a, which can localize to the cy-toskeleton and to the nucleus [54]; the NPA-bind-ing protein which localizes to the cytoskeleton and the plasma membrane [17], and sucrose synthase which can be cytosolic, membrane-associated or cytoskeleton-associated [18].
4.2. No6el macromolecular structures of the plant
cytoskeleton
Much of the previous discussion on novel protein components of the plant cytoskeleton is similar to that for novel macromolecular compo-nents, except that the task of getting results of the latter published is even more daunting. Despite this, we have adopted our approach to identifying novel protein components and used them to iden-tify novel macromolecular structures using the
cy-toskeleton-protective effects of the
cytoskeleton-stabilizing buffer (CSB). In addition to using HEPES (the basis of CSB) as the main buffer in tissue fixation, we have deliberately avoided the use of cacodylate, which is a herbicide [55] and deleterious to plant cell architecture. We also used a newly-designed TEM with medium-high (300 kV) acceleration voltage, which permits the visualization of sections as thick as 200 nm, without significant loss of resolution. Since we are interested in the association of ribosomes with filaments and membranes, and since individual ribosomes are 20 – 25 nm in diameter, conventional
(B50 nm) sections are not thick enough. We need
sections of sufficient thickness to decipher these three-dimensional relationships, and find that 100 – 200 nm is optimum. At voltages lower than 100 kV, resolution is impaired by chromatic
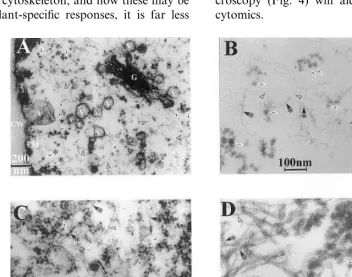
aber-ration in such thick sections, but elevating the voltage to 300 kV can cause specimen damage. Our microscope emits positive charges in numbers equal to the electrons, thereby neutralizing the electron damage, and so we can use 300 kV quite safely. Because of reduction in chromatic aberra-tion, this gives us resolution to about 0.5 nm and enables us to visualize filaments as thin as 4 nm. Using these techniques we have identified two quite different, yet novel structures. The first is a newly-identified tubular structure, about 11 – 13 nm in diameter, which is especially abundant in corn endosperm cells and provides a linkage be-tween the actin cytoskeleton, membranes, and protein bodies (Fig. 2), while the other is a small (4 nm) filament seen most abundantly in pea stems (Fig. 3). Two views of this new tubular structure are presented in the stereogram of the corn en-dosperm cytomatrix (Fig. 2A, B), which shows that abundant tubular filamentous structures of about 13 nm in external diameter are closely asso-ciated with polysomes, protein bodies, and vesi-cles. Since these structure are considerably smaller than microtubules, we refer to them as nan-otubules (NT). Pea stem tissue contains essentially identical nanotubules to those seen in corn en-dosperm and here they are seen in close conjunc-tion with ribosomes (Fig. 2C). We speculate that the NT might be another component of the cy-toskeleton involved in ribosome/polysome attach-ment, and thus implicated in protein synthesis. These structures are different from the 10 nm filaments reported in algae, which seem to be similar to intermediate filaments [56], and from the tubules in a maize mutant, which appear to reside within the endoplasmic reticulum [57]. Since these structures survive intact when isolated in vitro in a detergent-treated cytoskeleton fraction, they, like
the ‘nanofilaments’ below, cannot be
membranous.
E.Da6ies et al./Plant Science160 (2001) 185 – 196
192
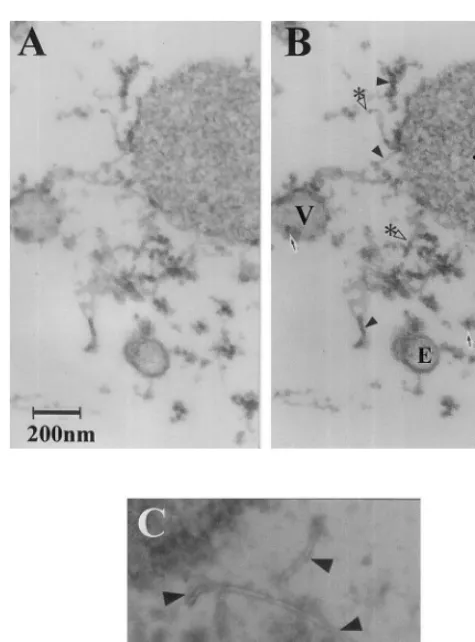
Fig. 2. Novel tubular structures visualized in situ in sweet corn endosperm and in pea stem tissue. Sweet corn en-dosperm (A, B) harvested 21 DAP was fixed in 100 mM HEPES-Na (pH 7.0) plus 5 mM MgC12 containing 2.5%
glutaraldehyde for 2 h, and post-fixed in the same buffer containing 1% OsO4. The samples were dehydrated in a 10 – 100% ethanol series and embedded in resin of 48.5 ml Epon 812, 18.5 ml DDSA (dodecenyl succinic anhydride), 33 ml MNA (methyl nadic anhydride) and 1.6 ml DMP-30 (2,4,6-tris[dimethylamino methyl] phenol), all from TAAB Labs, Berks, England. The embedded samples were then sectioned using an UltraCut microtorne (Leica) and stained in 7% uranyl acetate for 1 h, washed with dH20, and further
stained in 0.4% basic lead citrate for 10 min and then viewed at 300 kV with a JEM-3010HC microscope (JEOL, Tokyo, Japan). Etiolated pea stem tissue (C) was fixed, sectioned, embedded, stained and viewed as with corn endosperm, and observed under 300 kV at 100 000× magnification. (A, B) Stereogram of corn endosperm nanotubule (to be observed under a stereo pair viewer). Sample viewed at a magnification of 51 000×and viewed from two angles: A,+10°; B,−10°. V corresponds to a vesicle, E to endoplasmic reticulum, the asterisk (*) to a protein body, solid arrowheads to nan-otubules and open arrowheads with an asterisk (*) to cross sections of nanotubules. (C) Pea tissue viewed at higher magnification (100 000×). Solid arrowheads correspond to nanotubules, arrows to ribosomes. Pea tissue contains essentially identical nanotubules to those seen in corn en-dosperm and here they are seen in close conjunction with ribosomes.
to exist as 4 nm filaments. We are currently trying to identify both of these structures (NT and NF) using several antibodies.
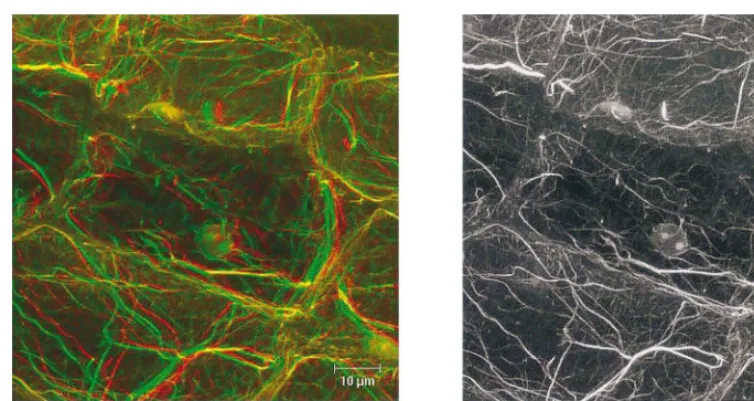
We anticipate that both magnetic resonance imaging [7] and laser scanning confocal mi-croscopy will furnish extremely powerful tools for visualizing the dynamic, spatio-temporal changes in protein location and function (i.e. cytomics). Again, the use of non-damaging techniques is imperative and, in this context, Fig. 4 shows the actin cytoskeleton as viewed in a sample ‘fixed’ in CSB plus detergent and stained with rhodamine-phalloidin. The stereogram or anaglyph (Fig. 4A) gives a clear view of the 3-dimensional nature of cytomatrix with the nucleus cradled in the actin cytoskeleton, while the corresponding black and white image shows very fine actin filaments similar to those shown earlier [26] and a surprisingly low non-specific fluorescence from the cell wall.
5. Future directions
We anticipate that plant-specific processes/ struc-tures involving the cytoskeleton will involve plant-specific cytoskeleton proteins [58]. Such processes (or structures) include gravitropism [45]; viral movement out of the nucleus [59] from cell-to-cell [60 – 62], through plasmodesmata [59] and in the phloem [63,64]; chloroplast movement and orien-tation [65]; host-parasite and host-symbiont rela-tionships [66], the endomembrane sheath (Reuzeau et al., [67,68]), and calcium channels [69].
cytoskeleton-associ-ated protein in animal tissues [70] and which we have found (data not shown) that it is cytoskele-ton-associated in plants, which will stimulate wa-ter uptake on the lower side. In addition, DnaK (Table 1) and DnaJ [45] might enhance protein translocation on (or to) the lower side. We wonder
whether the plasma membrane-associated
polysomes (Fig. 3A) might also play a role in gravitropism. Furthermore, since enzymes of the phosphoinositide pathway are cytoskeleton-associ-ated [16] and both ultra-rapid as well as long-term changes in IP3 levels occur after gravity tion [71], we speculate that there will be a stimula-tion of cytoskeleton-membrane interacstimula-tions and signaling [8] between the extra-cellular matrix and cytoplasm after gravity stimulation, and that IP3 will play a major role in this process.
While there is some basis for speculation as to what novel proteins may be found to be associated with the plant cytoskeleton, and how these may be involved in plant-specific responses, it is far less
easy to be prescient about the discovery of novel
macromolecular complexes making up novel
filamentous or tubular systems. Since the
condi-tions needed to stabilize/protect (known)
components of the cytoskeleton may be totally different from those needed to stabilize other
(novel) components, this mitigates against
the discovery of novel components, since reviewers
will likely refuse to accept images lacking
known components, and consider the novel com-ponents to be artifacts. Nevertheless, such efforts to identify novel components are essential to a full understanding of the cytoskeleton, and acceptance of the novel structures will be rendered easier if their protein components can be identified bio-chemically, as we are currently trying to do with both nanotubules (Fig. 2) and nanofilaments (Fig. 3). In this context, immuno-electron croscopy, and especially immuno-fluorescence mi-croscopy (Fig. 4) will aid in the study of plant cytomics.
E.Da6ies et al./Plant Science160 (2001) 185 – 196
194
Fig. 4. Laser scanning confocal fluorescence microscope image of pea stem tissue stained with rhodamine-phalloidin. A hand section of dark-grown pea stem tissue was permeabilized for 5 min in CSB+0.5% PTE and stained with rhodamine-phalloidin in CSB in the presence of 100 mM DTE (Abe and Davies, 1991), and viewed under a Leica microscope under excitation at 515 nm and emission at 570 – 590 nm. (A) Stereogram (anaglyph) to be viewed with red and green glasses. (B) Conventional B/W image.
In summary, we anticipate that more of these plant-specific cytoskeleton components will be dis-covered using plant-specific methods, rather then relying on techniques and probes derived from animal systems and we recommend that funding agencies support such novel approaches to deci-phering plants and not just rely on ‘me-too’ ap-proaches using animal-derived protocols. It is our hope that the proteomics of the plant cytoskeleton will soon become as well understood as is that of the chloroplast [72], so that a concerted effort can be launched into plant cytomics.
Acknowledgements
This work was supported in part by a grant from NASA (547 – 574) and from the North Caro-lina Agriculture Research Service (project No. 06446) to ED, by a Sasagawa Scientific Research Grant from the Japanese Science Society to KS, and a grant from the Japanese Ministry of Educa-tion, Science, Culture and Sports to SA for the purchase of the 300 kV IP-enhanced analytical electron microscope.
References
[1] D.E. Ingber, The riddle of morphogenesis: a question of solution chemistry or molecular cell engineering?, Cell 75
(1993) 1249 – 1252.
[2] P.S. Agutter, Nucleocytoplasmic mRNA transport: a plea for methodological dualism, Trends Cell Biol. 4 (1994) 278 – 279.
[3] P.A. Srere, H.R. Knull, Location — location — loca-tion, Trends Biochem. Sci. 23 (1998) 319 – 320.
[4] C. Masters, On the role of the cytoskeleton in metabolic compartmentation, in: J.E. Hesketh, I.M. Pryme (Eds.), The Cytoskeleton, Role in Cell Physiology, vol. 2, JAI Press, 1996, pp. 1 – 30.
[5] S. Penman, Rethinking cell structure, Proc. Natl. Acad. Sci. USA 92 (1995) 5251 – 5257.
[6] P.W. Hochachka, The metabolic implications of intra-cellular circulation, Proc. Natl. Acad. Sci. USA 96 (1999) 12233 – 12239.
[7] E. Straus, Detecting enzyme activity in live animals, Science 287 (2000) 1575.
[8] E. Davies, Intercellular and intracellular signals in plants and their transduction via the membrane-cytoskeleton interface, Semin. Cell Biol. 4 (1993) 139 – 147.
[9] E. Fuchs, Y. Yang, Crossroads on cytoskeleton high-ways, Cell 98 (1998) 547 – 550.
[10] S.R. Hameroff, Coherence in the cytoskeleton: implica-tions for biological information processing, in: H. Frohlich (Ed.), Biological Coherence and Response to External Stimuli, Springer, Berlin, 1987, pp. 242 – 265. [11] P.A. Janmey, The cytoskeleton and cell signaling:
com-ponent localization and mechanical coupling, Physiol. Rev. 78 (1998) 763 – 781.
[12] M.W. Klymkowsky, Weaving a tangled web: the inter-connected cytoskeleton, Nat. Cell Biol. 1 (1999) E121 – E123.
[14] D. Volkmann, F. Baluska, Actin cytoskeleton in plants: from transport networks to signaling networks, Microsc. Res. Tech. 47 (1999) 135 – 154.
[15] A. Abbot, A post-genomic challenge: learning to read patterns of protein synthesis, Nature 402 (2000) 715 – 720.
[16] Z. Tan, W.F. Boss, Association of phosphatidylinositol kinase, phosphatidylinositol monophosphate kinase, and diacylglycerol kinase with the cytoskeleton and F-actin fractions of carrot (Daucus carota L.) cells grown in suspension culture, Plant Physiol. 100 (1992) 2116 – 2120.
[17] J.H. Butler, S. Hu, S.R. Brady, M.W. Dixon, G.K. Muday, In vitro and in vivo evidence for actin associa-tion of the naphthylphthalamic acid-binding protein from zucchini hypocotyls, Plant J. 13 (1998) 291 – 301.
[18] H. Winter, J.L. Huber, S.C. Huber, Identification of sucrose synthase as an actin-binding protein, FEBS Lett. 430 (1998) 205 – 208.
[19] S. Abe, E. Davies, Methods for isolation and analysis of the cytoskeleton, in: Methods in Cell Biology, vol. 50, Academic Press, New York, 1995, pp. 223 – 236.
[20] E. Davies, B.D. Fillingham, S. Abe, The plant cytoskele-ton, in: J.E. Hesketh, I.M. Pryme (Eds.), The Cytoskele-ton, Role in Specialized Tissues, vol. 3, JAI Press, 1996, pp. 405 – 449.
[21] P.J. Hussey, J.A. Traas, K. Gull, C.W. Lloyd, Isolation of cytoskeletons from synchronized plant cells: the inter-phase microtubule array utilizes multiple tubulin iso-types, J. Cell Sci. 88 (1987) 225 – 230.
[22] P. Xu, C.W. Lloyd, C.J. Staiger, B.K. Drobak, Associa-tion of phosphatidylinositol 4-kinase with the plant cy-toskeleton, Plant Cell 4 (1992) 941 – 951.
[23] J. Chan, C.G. Jensen, L.C.W. Jensen, M. Bush, C.W. Lloyd, The 65-kDa carrot microtubule-associated protein forms regularly arranged filamentous cross-bridges between microtubules, Proc. Natl. Acad. Sci. USA 96 (1999) 14931 – 14936.
[24] J.M. Andersland, D.C. Dixon, R.-W. Seagull, B.A. Triplett, Isolation and characterization of cytoskeletons from cotton fiber cytoplasts, In Vitro Cell. Dev. Biol. — Plant 34 (1998) 173 – 180.
[25] B. Stankovic, A. Clore, B. Larkins, S. Abe, E. Davies, Actin in protein synthesis and protein body formation, in: C.J. Staiger, F. Baluska, D. Volkmann, P. Barlow (Eds.), Actin: a Dynamic Framework for Multiple Plant Cell Functions, Kluwer Academic Publishers, Dor-drecht, 2000, pp. 129 – 143.
[26] S. Abe, E. Davies, Isolation of F-actin from pea stems: evidence from fluorescence microscopy, Protoplasma 163 (1991) 51 – 61.
[27] E. Davies, B.D. Fillingham, Y. Ito, S. Abe, Evidence for the existence of cytoskeleton-bound polysomes in plants, Cell Biol. Int. 15 (1991) 973 – 981.
[28] W. You, S. Abe, E. Davies, Co-sedimentation of pea root polysomes with the cytoskeleton, Cell Biol. Int. Rep. 16 (1992) 663 – 673.
[29] S. Abe, W. You, E. Davies, Protein bodies in corn
endosperm are enclosed by and enmeshed in F-actin, Protoplasma 165 (1991) 139 – 149.
[30] E. Davies, E.C. Comer, J.M. Lionberger, B. Stankovic, S. Abe, Cytoskeleton-bound polysomes in plants: polysome-cytoskeleton-membrane interactions in corn endosperm, Cell Biol. Int. 17 (1992) 331 – 339.
[31] B. Stankovic, S. Abe, E. Davies, Co-localization of polysomes, cytoskeleton and membranes with protein bodies from corn endosperm: evidence from fluorescence microscopy, Protoplasma 177 (1993) 66 – 72.
[32] B. Stankovic, S. Abe, K. Azama, K. Shibata, Y. Ito, S. Weidner, E. Davies, Associations of maize protein bod-ies with cytoskeleton, membranes, and ribosomes in the endosperm of wild type and opaque-2 mutant, Acta Physiol. Plantarum 21 (1999) 383 – 389.
[33] A.M. Clore, J.M. Dannenhoffer, B.A. Larkins, EF-la is associated with a cytoskeletal network surrounding protein bodies in maize endosperm cells, Plant Cell 8 (1996) 2003 – 2014.
[34] E. Davies, S. Abe, B.A. Larkins, A.M. Clore, R.S. Quatrano, S. Weidner, The role of the cytoskeleton in plant protein synthesis, in: J. Bailey-Serres, D.R. Gallie (Eds.), A Look Beyond Transcription: Mechanisms De-termining mRNA Stability and Translation in Plants, ASPP, 1998, pp. 115 – 124.
[35] T.W. Okita, X. Li, M.W. Roberts, Targeting of mRNAs to domains of the endoplasmic reticulum, Trends Cell Biol. 4 (1994) 91 – 96.
[36] D.G. Muench, Y. Wu, S.J. Coughlan, T.W. Okita, Evidence for a cytoskeleton-associated binding site in-volved in prolamine mRNA localization to the protein bodies in rice endosperm tissue, Plant Physiol. 116 (1998) 559 – 569.
[37] O. Numata, T. Takemasa, I. Takai, M. Hironi, J. Chiba, Y. Watanabe, Tetrahymena 14-nm filament-forming protein has citrate synthase activity, Biochem. Biophys. Res. Commun. 174 (1991) 1028 – 1034.
[38] Y. Kurasawa, O. Numata, M. Katoh, H. Hirano, J. Chiba, Y. Watanabe, Identification of Tetrahymena 14-rim filament-associated protein as elongation factor 1 alpha, Exp. Cell Res. 203 (1992) 251 – 258.
[39] F. Cao, N. Yanagihara, J.M. Burke, Progressive associa-tion of a soluble glycolytic enzyme with the detergent-in-soluble cytoskeleton during in vitro morphogenesis of MCCK epithelial cells, Cell Motil. Cytoskeleton 44 (1999) 133 – 142.
[40] E.V. Koonin, P. Bork, C. Sander, A novel RNA-binding motif in omnipotent suppressors of translation termina-tion, ribosomal proteins and a ribosome-modifying en-zyme?, Nucleic Acids Res. 22 (1994) 2166 – 2167. [41] K. Shibata, Y. Morita, S. Abe, B. Stankovic, E. Davies,
Apyrase from pea stems: isolation, purification, charac-terization and identification of a NTPase from the cy-toskeleton fraction of pea stem tissue, Plant Physiol. Biochem. 37 (1999) 881 – 888.
E.Da6ies et al./Plant Science160 (2001) 185 – 196
196
[43] R.A. Ivey, B.D. Bruce, In vivo and in vitro interaction of DnaK and a chloroplast transit peptide, Cell Stress Chaperones 1 (2000) 62 – 71.
[44] E.A. Shestakova, L.P. Motuz, L.P. Gavrilova, Co-local-ization of components of the protein-synthesizing ma-chinery with the cytoskeleton in GO-arrested cells, Cell Biol. Int. 17 (1993) 417 – 424.
[45] J.C. Sedbrook, R. Chen, P.H. Masson, ARG1 (altered response to gravity) encodes a DnaJ-like protein that potentially interacts with the cytoskeleton, Proc. Natl. Acad. Sci. USA 96 (1999) 1140 – 1145.
[46] X. Li, Y. Wu, D.-Z. Zhang, J.W. Gillikin, R. Boston, V.R. Franceschi, T.W. Okita, Rice prolamine protein body synthesis is a BiP-mediated process, Science 262 (1993) 1054 – 1056.
[47] Y. Wu, D.G. Muench, Y.-T. Kim, Y.-S. Hwang, T.W. Okita, Identification of polypeptides associated with an enriched cytoskeleton-protein body fraction from devel-oping rice endosperm, Plant Cell Physiol. 39 (1998) 1251 – 1257.
[48] N.R. Smalheiser, Proteins in unexpected locations, Mol. Biol. Cell 7 (1996) 1003 – 1011.
[49] K.C. Chou, D.W. Elrod, Protein sub-cellular location predictions, Protein Eng. 12 (1999) 107 – 118.
[50] D. Kraemer, G. Blobel, mRNA binding protein mrnp 41 localizes to both nucleus and cytoplasm, Proc. Natl. Acad. Sci. USA 94 (1997) 9119 – 9124.
[51] J. Saas, K. Ziegelbauer, A. von Haeseler, B. Fast, M. Boshart, A developmentally-regulated aconitase related to iron-regulatory protein-1 is localized in the cytoplasm and in the mitochondrion of Trypanosoma brucei, J. Biol. Chem. 275 (1999) 2745 – 2755.
[52] C. Thomas, Y. Sun, K. Naus, A. Lloyd, S. Roux, Apyrase functions in plant phosphate nutrition and mo-bilizes phosphate from extracellular ATP, Plant Physiol. 119 (1999) 543 – 551.
[53] C. Thomas, A. Rajagopal, B. Windsor, R. Dudler, A. Lloyd, S.J. Roux, A role for ectoapyrase in xenobiotic resistance, Plant Cell 12 (2000) 519 – 533.
[54] H.M.S. Smith, N.V. Raikhel, Nuclear localization signal receptor importin-a associates with the cytoskeleton, Plant Cell 10 (1998) 1791 – 1799.
[55] C.-C. Chu, T. Henneberry, R. Reynoso, Effect of cotton defoliants on leaf abscission, immature bolls, and lint yields in a short-season production system, J. Prod. Agric. 5 (1992) 268 – 272.
[56] S. Berger, R.L. Shoeman, P. Traub, Detection of dense intra- and perinuclear 10 nm filament systems by whole mount and embedment-free electron microscopy in sev-eral species of the green algal order Dasycladales, Proto-plasma 190 (1996) 204 – 220.
[57] C.R. Lending, A novel tubular array associated with protein bodies in the rough endoplasmic reticulum of opaque-2 maize, Protoplasma 195 (1996) 68 – 77.
[58] N.C.A. De Ruijter, A.M.C. Emons, Actin-binding proteins in plant cells, Plant Biol. 1 (1999) 26 – 35. [59] J.-Y. Lee, B.-C. Yoo, W.J. Lucas, Parallels between
nuclear-pore and plasmodesmatal trafficking of informa-tion molecules, Planta 210 (2000) 177 – 187.
[60] M. Heinlein, B.L. Epel, H.S. Padgett, R.-N. Beachy, Interaction of tobamovirus movement proteins with the plant cytoskeleton, Science 270 (1995) 1983 – 1985. [61] A.A. Sanderfoot, S.G. Lazarowitz, Getting it together in
plant virus movement: cooperative interactions between bipartite geminivirus movement proteins, Trends Cell Biol. 6 (1996) 353 – 358.
[62] B. Xoconostle-Cazares, Y. Xiang, R. Ruiz-Medrano, H.-L. Wang, J. Monzer, B.-C. Yoo, K.C. McFarland, V.R. Francheschi, W.J. Lucas, Plant paralog to viral movement protein that potentiates transport of mRNA into the phloem, Science 283 (1999) 94 – 98.
[63] B.G. Pickard, R.N. Beachy, Intercellular connections are developmentally controlled to help move molecules through plants, Cell 98 (1999) 5 – 8.
[64] G.A. Thompson, A. Schultz, Macromolecular trafficking in the phloem, Trends Plant Sci. 4 (1999) 354 – 360. [65] M.K. Kandasamy, R.B. Meagher, Actin-organelle
inter-action: association with chloroplast in Arabidopsis leaf mesophyll cells, Cell Motil. Cytoskeleton 44 (1999) 110 – 118.
[66] A. Genre, P. Bonfante, Cytoskeleton-related proteins in tobacco mycorrhizal cells: gamma-tubulin and clathrin localisation, Eur. J. Histochem. 43 (1999) 105 – 111. [67] C. Reuzeau, K.W. Doolittle, J.G. McNally, B.G.
Pick-ard, Covisualization in living onion cells of putative integrin, putative spectrin, actin, putative intermediate filaments, and other proteins at the cell membranes and in an endomembrane sheath, Protoplasma 199 (1997) 173 – 197.
[68] C. Reuzeau, J.G. McNally, B.G. Pickard, The en-domembrane sheath: a key structure for understanding the plant cell?, Protoplasma 200 (1997) 1 – 9.
[69] P. Thuleau, J.L. Schroeder, R. Ranjeva, Recent ad-vances in the regulation of plant calcium channels: evi-dence for regulation by G-proteins, the cytoskeleton and second messengers, Curr. Opin. Plant Biol. 1 (1998) 424 – 427.
[70] I. Sabolic, T. Katsura, J.-M. Verbavatz, D. Brown, The AQP2 water channel: effect of vasopressin treatment, microtubule disruption, and distribution in neonatal rats, J. Membr. Biol. 143 (1995) 165 – 175.
[71] I.M. Perera, I. Heilmann, W.F. Boss, Transient and sustained increases in inositol 1,4,5-trisphosphate pre-cede the differential growth response in maize pulvini, Proc. Natl. Acad. Sci. 96 (1999) 5838 – 5843.
[72] J.-B. Peltier, G. Friso, D.E. Kalume, P. Roepstorff, F. Nilsson, I. Adamska, K.J. van Wyk, Proteomics of the chloroplast: systematic identification and targeting anal-ysis of luminal and peripheral thylakoid proteins, Plant Cell 12 (2000) 319 – 341.