Floral induction in Eucalyptus nitens
M. W. MONCUR1 and O. HASAN2
1 CSIRO, Division of Forestry, P.O. Box 4008, Canberra, ACT 2600, Australia
2 Cooperative Research Center for Temperate Hardwood Forestry and Department of Plant Science,
University of Tasmania, GPO Box 252C, Hobart, Tasmania 7001, Australia
Received January 3, 1994
Summary
Eucalyptus nitens (Deane & Maiden) Maiden takes at least five years to initiate flower buds from seed
and is an infrequent and light flowerer. Because this behavior constitutes a major impediment to breeding programs, we examined the mechanisms controlling floral induction in E. nitens, with the long-term aim of reducing generation time and increasing seed yield.
Application of paclobutrazol reduced the concentration of endogenous gibberellic acid (GA) in apical tissue and enhanced the reproductive activity of grafted trees maintained outside over winter in Canberra, Australia. Grafts maintained in a warm greenhouse over winter did not produce flower buds, despite the paclobutrazol-induced reduction in GA concentration of the apical tissue. Exposing untreated grafts, which had been maintained over winter in a warm greenhouse, to low temperature the following spring reduced growth but did not induce flower bud production. Addition of GA3 to paclobutrazol-treated grafts
reduced the effect of paclobutrazol on reproductive activity.
Keywords: flowering, generation time, gibberellins, low temperature, paclobutrazol, seed yield.
Introduction
Tree breeding and seed orchard production depend on early and consistent flowering, neither of which is characteristic of forest trees in general. Species of the Eucalyptus genus are important for forestry both in Australia and overseas, and several genetic improvement programs are in progress. However, flower induction in Eucalyptus is not well understood. Eucalyptus species produce flower buds in the leaf axils of new growth in spring following a cold winter (Tibbits 1989), suggesting that changing day length or exposure to low temperature may be involved in the flower induction process. A number of Australian native species have a low temperature requirement for initiation of flowering (King et al. 1992). Eucalyptus lansdowneana F. Muell. & J. Brown initiates flower buds when plants are transferred from a warm to a cold regime for 4--6 weeks and then back to a warm regime, regardless of day length (Moncur 1992), whereas Bolotin (1975) reported that Eucalyptus occidentalis Endl. seedlings, less than one year old, flowered precociously when grown in a long day regime of 16 h or longer. Floral initiation in Eucalyptus nitens (Deane & Maiden) Maiden occurs in spring following a cold winter (Moncur, unpublished data), but the mechanisms controlling flowering have yet to be determined.
Hetherington et al. 1991, Griffin et al. 1993, Moncur et al. 1994). Moncur et al. (1994) reported that a single application of paclobutrazol resulted in reduced endo-genous GA concentrations in E. nitens plants six to eight weeks before flower buds were first visible.
We have examined the effects of low temperatures and paclobutrazol on flower bud production by E. nitens grafts. We used a single clone to avoid provenance variation, and grafts were used because E. nitens, in common with many Eucalyptus species, has a well defined juvenile period (Jacobs 1955). Although there have been no reports of reversion to juvenility following grafting in eucalypts, this possibility was tested by the use of different aged grafts.
We investigated the association between the concentration of active GA in apical tissues at the time of flower bud initiation and the number of flower buds observed 12 weeks later. The hypothesis that paclobutrazol enhances flowering by reducing GA concentrations was tested by treating paclobutrazol-treated material with a commercially available GA with very similar structure and activity to endogenous GA1.
Materials and methods
In October 1989 and 1990, scions from mature E. nitens trees of a single clone were grafted onto E. nitens seedling root stock originating from the Associated Pulp and Paper Mills (now North Forest Products) breeding program at Ridgley, Tasmania. Eighty grafts were potted in 460 or 380 mm containers filled with a 3/1/1/1 (v/v) mix of composted tan bark, perlite, vermiculite and rice hulls, and grown outdoors in Canberra, Australia. The grafts were fertilized with Osmocote, irrigated and sprayed to control insects as required. Grafts were repotted at the end of the first year to avoid root constriction.
Effects of temperature and paclobutrazol on 6- and 18-month-old E. nitens grafts
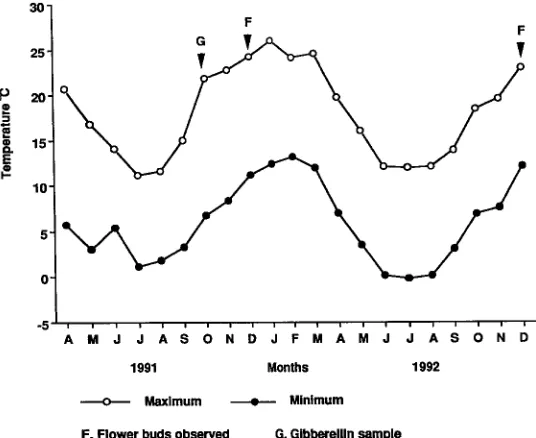
In April 1991, paclobutrazol was applied at the rate of 0.02 g of active ingredient per mm of stem circumference. The required dose was added to 400 ml of water for 18-month-old grafts or 200 ml of water for 6-month-old grafts and applied as a root drench. Five grafts of each age were then placed in a naturally lit greenhouse with a day (0800--1600 h)/night temperature of 25/17 °C. The remaining grafts were grown outside (see Figure 2). A further five grafts of each age were transferred to the greenhouse in June 1991 and September 1991. Grafts were held in these conditions until early December 1991, when flower bud production is normally completed, and then all grafts were moved outdoors until December 1992.
Extracts were purified by C18 Sep-Pak chromatography, anion-exchange
chromatog-raphy and high performance liquid chromatogchromatog-raphy (Hasan et al. 1994). Both GA1
and GA20 were subsequently quantified by gas chromatography--mass spectrometry
in the selected ion monitoring mode (Hasan et al. 1994).
Relationship between endogenous GA concentrations and flower bud production in espaliered grafts of E. nitens
In April 1989, one-year-old grafts maintained in the Canberra espalier orchard were treated with one of three concentrations of paclobutrazol by either trunk injection or soil drench (Moncur et al. 1994). During late September 1992, samples of apical shoot tissue, including meristematic apices and developing leaves, were harvested from the four-year-old grafted trees and quantified for GA1 concentrations according
to the methods described by Hasan et al. (1994). Flower buds were recorded in mid-December 1992.
Interaction between GA3 and paclobutrazol on growth and flower bud development on E. nitens grafts
Grafts were established in October 1991 (Figure 1). Each graft was potted in a 300 mm container filled with a 1/1/1/3 (v/v) mixture of rice hulls, perlite, vermiculite and composted tan bark. In April 1992, 90 grafts were placed in a factorial combina-tion of paclobutrazol with two concentracombina-tions of GA3. Paclobutrazol was applied at
20 mg of active ingredient per mm of stem circumference in 200 ml of water as a root drench. An aqueous stock solution of 0.3 mM GA3 was applied as a soil drench on
June 5 and July 2, 1992 at two rates (High-GA3 = 50 ml, Low-GA3 = 10 ml). Grafts
were grown outdoors until December 1992 when flower bud production was com-plete.
Effects of continuous and cyclic temperatures on floral induction in E. nitens
Scions were grafted onto seedling root stock and grown at Ridgley, Tasmania. Twenty nine-month-old grafts were transferred to Canberra and placed in the CSIRO Phytotron. Grafts were potted in 250 mm containers filled with a 1/1 (v/v) mix of perlite and vermiculite. Each pot was watered with Hoagland’s nutrient solution twice a week, increasing to three times a week after Week 10, and tap water as required. Pots were placed in a greenhouse with a day (0830-1630 h)/night tempera-ture of 21/16 °C. The photoperiod was extended to 16 h by use of incandescent lighting from 0430--0830 and 1630--2030 h.
After two weeks, 10 grafts were placed in a warm diurnal regime (24/19 °C) in an open greenhouse, and another 10 grafts were subjected to a cyclic temperature treatment consisting of one week in a naturally lit cabinet at 15/10 °C, followed by six weeks at 7/4 °C, one week at 15/10 °C, four weeks at 24/19 °C and finally, one week at 15/10 °C. This treatment was repeated until Week 25 when grafts were too large for the cabinet and were transferred to a larger cabinet with a temperature regime of 10/5 °C. Both treatments received a photoperiod extended to 16 h with incandescent lighting.
Treatments were terminated after 35 weeks and the grafts were then transplanted to 300 mm pots and grown outside (Figure 2). Five grafts remained outdoors at Ridgley for comparison with the experimental grafts.
Results
Paclobutrazol suppressed height growth and leaf production (Table 1), but flowering occurred in the paclobutrazol-treated grafts that were exposed to low temperatures over winter (Figure 2, Table 2). By December 1992 (Year 2), all grafts had experi-enced two winters outdoors and those treated with paclobutrazol produced flower buds (Table 2). Two 18-month-old and two 6-month-old paclobutrazol-treated grafts that initiated flower buds in Year 1 (Table 2) were grown in a greenhouse (25/18 °C) during the second winter. In contrast to the grafts maintained outdoors over the second winter, the grafts in the warm greenhouse failed to produce flower buds the following season.
At the time of sampling the espalier orchard in late September, the concentration of GA1 in the apical meristematic tissue was related to the number of flower buds
produced in December (Figure 3). Gibberellin concentrations greater than approxi-mately 0.6 ng gDW−1 in the apical meristematic tissue were associated with a marked
reduction in subsequent flower bud production. However, lower GA1 concentrations
responses. Treatment with paclobutrazol can reduce GA concentrations (Table 3) without a flowering response (Table 2).
Application of GA3 by soil drench significantly increased stem diameter (P <
0.001), indicating that applied GA3 was assimilated by the grafts. The stem
thicken-ing response has been noted as the most dramatic effect of GA3 application on
vegetative tissues of apples (Mauk et al. 1990), which may explain its detection in the absence of any significant effect on height increment (P = 0.863) (Table 4b). Flower bud numbers were reduced by up to 40% following application of GA3. There
were more grafts with less than 25 flower buds in the paclobutrazol + Low-GA3 and
paclobutrazol + High-GA3 treatments than in the paclobutrazol treatment (data not
shown), indicating a reduction in reproductive activity following GA3 application.
Growth of untreated 6- and 18-month-old grafts was continuous in the greenhouse, but growth was suppressed when grafts were grown outside at low temperatures (Table 1). The height and number of leaves of grafted trees increased with increasing exposure to warm greenhouse conditions (Table 1). No flower buds were produced by untreated grafts in Year 1; however, in Year 2, one untreated graft produced flower buds (Table 2).
Height increment and leaf production of grafts grown at 24/19 °C in the Phytotron increased over time. When grafts were transferred to low temperatures (cyclic treatment), growth virtually ceased, but growth resumed when the grafts were returned to 24/19 °C. When grafts were returned to 24/19 °C, leaves were produced
at a faster rate than for grafts grown continuously at 24/19 °C. Leaf production in both treatments declined with age. All new leaves were vegetatively adult, as indicated by their alternate and petiolate appearance. No flower buds were observed under either regime. By late November, following transfer of grafts outdoors, flower buds were observed in 20% of the surviving grafts with no significant difference between initial treatments. Grafts at Ridgley produced adult leaves, but no flower buds.
There was no indication of reversion to juvenility, as demonstrated by the produc-tion of juvenile foliage, following treatment of grafted material with paclobutrazol. The effect of paclobutrazol on flowering was similar for both the 6- and 18-month-old grafts, indicating that juvenility was not a confounding factor.
Discussion
Under natural conditions, E. nitens produces flower buds at about age five years, and
Table 1. Effects of paclobutrazol and duration of cold treatment on height increment and number of leaves of 6- and 18-month-old grafts of E. nitens grown at Canberra.
TreatmentPaclobutrazolControl
HeightNo. of leavesHeightNo. of leaves
Oct1Dec2OctDecOctDecOctDec
18-Month-old grafts
April3202206 7.811.232839224.632.0 June204213 6.810.628536421.828.6 September17718110.612.221027115.423.8 Outside201214 8.415.619423810.217.8
6-Month-old grafts
April 90 98 8.213.419830525.833.0 June 97101 7.2 9.419726924.633.0 September 94 98 8.0 8.412018115.626.6 Outside 92 98 8.2 9.011217614.223.0
ANOVAHeightNo. of leaves
production increases with age. Although the mechanisms for induction are not known, trees growing in natural environments will experience a series of cold periods, changing day lengths and, in many cases, variations in water status. The effect of these stimuli, either singly or in combination, may be cumulative, resulting in a flowering threshold being surpassed after several potentially inductive events.
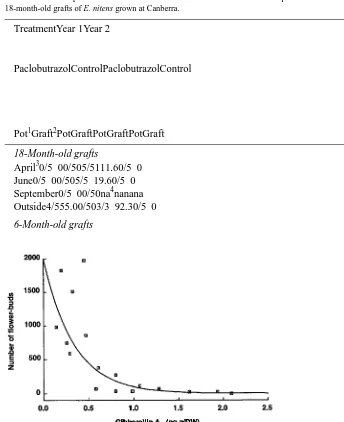
Table 2. Effects of paclobutrazol and duration of cold treatment on flower bud production in 6- and 18-month-old grafts of E. nitens grown at Canberra.
TreatmentYear 1Year 2
PaclobutrazolControlPaclobutrazolControl
Pot1Graft2PotGraftPotGraftPotGraft
18-Month-old grafts
April30/5 00/505/5111.60/5 0 June0/5 00/505/5 19.60/5 0 September0/5 00/50na4nanana Outside4/555.00/503/3 92.30/5 0
6-Month-old grafts
Figure 3. Relationship between flower bud production and GA1 in an E. nitens espaliered orchard.
Concentration of GA1 was detected in apical tissue by the method of Hasan et al. (1994). Regression: y =
Exposure to low temperatures in the absence of paclobutrazol treatment did not produce a floral response in Year 1, although a few grafts produced a small number of flower buds after a second winter outside. The apparent inability of changing day length to induce flowering in combination with paclobutrazol, but in the absence of maximum cold treatment, suggests that photoperiod is not a strong stimulus of flowering in E. nitens. This contrasts with the results of Bolotin (1975) for E.
occi-dentalis and, may be indicative of a species-specific response within the genus.
All grafts that flowered had low GA concentrations. The action of paclobutrazol in stimulating reproductive activity by reducing net biosynthesis of endogenous GA was also partially substantiated by the finding that reproductive activity in paclobu-trazol-treated grafts partially reverted to the untreated state following treatment with GA3. Enhanced growth in GA3-treated grafts may have redirected assimilates away
from reproductive centers, resulting in a reduction in reproductive activity. The application of GA generally inhibits flowering in woody angiosperms. Most notably, GA3 and GA4/7 inhibit or suppress flowering in a wide variety of fruit trees
(refer-ences cited by Pharis and King 1985).
Our data indicate that the concentration of active GA is not the sole factor determining the induction of floral primordia in E. nitens. Paclobutrazol-treated material exposed to different low temperature treatments had similar GA concentra-tions, although only material receiving maximum cold exposure produced flower buds. This response may resemble the vernalization response in annual plants, though its function in a woody perennial species is not clear. A period of cold could result in destruction of a flowering inhibitor, a change in inter-organ competition, or both. Metzger (1985) suggests that vernalization activates one or more steps in GA biosynthesis such that a specific GA accumulates above a threshold concentration.
The promotion of flowering by paclobutrazol constitutes a practical method of promoting flowering for breeding and seed production in several eucalypt species. This finding is particularly important in E. nitens, because domestication of this species has been limited by shortage of seed. Detailed characterization of the environmental stimuli of flowering could further enhance the promotive effects of paclobutrazol application, as well as indicating ideal sites to conduct and establish seed orchards.
Table 3. Effects of paclobutrazol and duration of cold treatment on endogenous GA concentrations (pg gFW−1) in apical tissue of 18-month-old grafts of E. nitens growing at Canberra.
TreatmentGA1GA20
ControlPaclobutrazolControlPaclobutrazol
Acknowledgments
We are grateful to Associated Pulp and Paper Mills, Ridgley, Tasmania for supplying the clones and financial assistance to conduct the Phytotron work. We thank CSIRO, Division of Plant Industry staff at the Phytotron for maintaining the trees utilized in the second experiment and J. Turner for technical assistance with the remaining experiments. R.P. Pharis gave valuable advice with GA3 applications.
R. King, P. Kriedemann and D. Sheriff made many valuable comments on an earlier draft.
Table 4. Effects of applied GA31 on (a) flower bud production and GA concentrations, and (b) height,
leaves, diameter and volume2 in grafts of E. nitens.
(a)
TreatmentNumber of potsMean flowerEndogenous GA concentration with flower budsbuds per pot(pg gFW−1)
GA1GA20
Control 10.701.710.58
Paclobutrazol (P)1033.300.440.14 P + Low-GA31218.700.550.21
P + High-GA3 917.800.520.18
Control + Low-GA3 00.001.570.56
Control + High-GA3 10.301.760.93
ANOVA P< 0.001 GA0.293 P × GA0.342
s.e.d P4.65 GA5.70 P × GA8.06
(b)
TreatmentIncrement increase (April--December)
HeightLeavesDiameterStem volume (mm)(mm)(cm3)
Control56713.806.2355.09
Paclobutrazol (P)1494.673.3915.36 P + Low-GA31454.873.6116.41
P + High-GA31516.534.3621.07
Control + Low-GA357514.875.7138.94
References
Bolotin, M. 1975. Photoperiodic induction of precocious flowering in a woody species, Eucalyptus
occidentalis Endl. Bot. Gaz. 136:358--365.
Griffin, A.R., P. Whiteman, T. Rudge, I.P. Burgess and M.W. Moncur. 1993. Enhancement of flower initiation in two species of Eucalyptus with paclobutrazol. Can. J. For. Res. 23:640--647.
Hasan, O., B.G. Ridoutt, N.W. Davies and J.B. Reid. 1994. Identification and quantification of endo-genous gibberellins in apical buds and the cambial region of Eucalyptus. Physiol. Plant. In press. Hetherington, S. and K.M. Jones. 1990. The effect of paclobutrazol on the growth of Eucalyptus globulus
seedlings. Can. J. For. Res. 20:1811--1813.
Hetherington, S., K.M. Jones and T.B. Koen. 1991. Stimulation of bud production in Eucalyptus globulus by paclobutrazol applications. In Intensive Forestry: The Role of Eucalypts. Ed. A.P.G. Schonau. IUFRO, Durban, pp 104--109.
Jacobs, M.R. 1955. Growth habits of the eucalypts. Commonwealth Government Printer, Canberra, 262 p.
Jones, K.M., T.B. Koen, M.J. Oakford and S.B. Longley. 1989. Using Ethephon and Daminozide to regulate growth and initiate flower buds on bearing ‘‘Red Delicious’’ trees. Acta Hortic. 240:185--188. King, R.W., I.A. Dawson and S.S. Speer. 1992. Control of growth and flowering in two Western
Australian species of Pimelea. Aust. J. Bot. 40:377--388.
Mauk, C.S., C.R. Unrath, S.M. Blankenship and L.J. Lehman. 1990. Influence of method of application of paclobutrazol on soil residues and growth retardation in a ‘‘Starkrimson-Delicious’’ apple orchard. Plant Growth Regul. 9:27--35.
Metzgar, J.D. 1985. The role of gibberellins in the environmental control of stem growth in Thiaspi
arvense L. Plant Physiol. 78:8--13.
Moncur, M.W. 1992. Effect of low temperature on floral induction of Eucalyptus lansdowneana F. Muell. & J. Brown subsp. lansdowneana. Aust. J. Bot. 40:157--167.
Moncur, M.W., G.F. Rasmussen and O. Hasan. 1994. Effect of paclobutrazol on flower bud production in Eucalyptus nitens (Deane & Maiden) Maiden espalier seed orchards. Can. J. For. Res. 24:46--49. Pharis, R.O. and R.W. King. 1985. Gibberellins and reproductive development in seed plants. Annu. Rev.
Plant Physiol. 36:517--568.