BAB I PENDAHULUAN
A. Latar Belakang Masalah
Kanker payudara merupakan tumor ganas yang dimulai dari sekelompok
sel-sel kanker yang dapat tumbuh menyerang jaringan sekitarnya atau menyebar
(metastasis) ke jaringan lain pada tubuh sebagai akibat dari ketidaknormalan
proses apoptosis. Apoptosis atau kematian sel yang terprogram adalah mekanisme
pengendalian terhadap eliminasi sel-sel yang mati pada organisme multiseluler.
Sel dikontrol dan diregulasi selama proses apoptosis, sel yang mati kemudian
difagosit oleh makrofag. Apoptosis berfungsi secara fisiologis antara lain untuk
terminasi sel, mempertahankan homeostasis, perkembangan embrional, interaksi
limfosit, dan involusi hormonal pada usia dewasa (Lumongga, 2008).
Sel-sel yang mati pada proses apoptosis memberikan sinyal dan
diperantarai oleh beberapa gen yang mengkode protein untuk enzim pencernaan
yang disebut caspase. Peristiwa apoptosis terjadi setelah adanya gangguan fungsi
barier membran mitokondria dengan pelepasan sitokrom c melalui jalur caspase-3
pada sel normal. Aktivasi caspase-3 dipengaruhi oleh protein keluarga Bcl-2.
Protein ini terdiri dari pro-apoptosis seperti Bax dan Bak dan anti-apoptosis
seperti Bcl-2 dan Bcl-XL. Overekspresi Bcl-2 anti-apoptosis mencegah aktivasi
caspase-3 pada tahap pelepasan sitokrom c, sehingga apoptosis tidak terjadi. Hal
inilah yang terjadi pada sel kanker payudara MCF-7 (Mooney et al., 2002).
Beberapa senyawa yang terbukti berhasil menghambat aktivitas Bcl-2,
diantaranya pirimidinilpiperazin yang mengikat prosurvival protein Bcl-XL
(Shallal, 2011 cit Green dan Evan, 2002), fenilpirazol dengan afinitas tinggi
mengikat Bcl-2 dan Bcl-XL (Porter et al., 2008 cit Zhang et al., 2007), kendomisin
(Janssen, 2008), dan navitoklaks (Wilson et al., 2010). Kurkumin (bis-α,β -unsaturated β-diketone) dikenal sebagai senyawa yang memiliki aktivitas antiinflamasi dan antikanker (Sharma et al., 2005). Aktivitas biologi kurkumin
pada model praklinis antara lain sitotoksik dan menginduksi apoptosis (Kuo et al.,
1996), menghambat karsinogenesis (Perkins et al., 2002), mempengaruhi fase I
(Firozi et al., 1996) dan II metabolisme enzim karsinogen (Singh et al., 1998),
antioksidan (Chan et al., 1998), serta mempengaruhi angiogenesis (Sharma,
2001). Kurkumin menginduksi apoptosis ketika siklus sel berada pada fase G2
atau M di dalam sel MCF-7 (Sharma et al., 2005). Senyawa lain yang memiliki
aktivitas antikanker adalah diketopiperazin (Aoki et al., 2010 cit Martin dan
Carvalho, 2007) yang merupakan analog dari kurkumin. Diketopiperazin memiliki
beberapa turunan, diantaranya adalah turunan diketopiperazin C-alkil. Aktivitas
inhibisi sel kanker senyawa ini dikarenakan interaksi cincinnya (Deveau et al.,
2008).
Penelitian ini menarik dilakukan dalam rangka penemuan obat baru
berbasis struktur molekul obat, melihat potensi yang dimiliki turunan
diketopiperazin C-alkil diharapkan dapat digunakan untuk menghambat
overekspresi Bcl-2 pada sel MCF-7 sehingga apoptosis dapat terjadi.
B. Perumusan Masalah
Berdasarkan latar belakang masalah yang telah diuraikan, maka
perumusan masalah dari penelitian ini adalah:
1. Mengkaji kedekatan interaksi geometri 3D turunan diketopiperazin C-alkil
dengan senyawa penuntun dan senyawa yang telah diketahui potensinya
sebagai inhibitor Bcl-2?
2. Apakah turunan diketopiperazin C-alkil mempunyai aktivitas sebagai inhibitor
Bcl-2 secara perhitungan docking molekular?
C. Tujuan Penelitian
Berdasarkan uraian perumusan masalah di atas, maka tujuan dari
penelitian ini adalah:
1. Mengetahui kedekatan bentuk geometri 3D turunan diketopiperazin C-alkil
dengan senyawa penuntun dan senyawa yang telah diketahui potensinya
sebagai inhibitor Bcl-2.
2. Memperoleh senyawa baru turunan diketopiperazin (DKP) yang berpotensi
D. Tinjauan Pustaka
1. Docking Molekular
Docking molekular adalah metode komputasi untuk memprediksi
geometri kompleks protein-ligan (Pyrkov et al., 2008). Metode docking dapat
dilakukan dengan 2 cara. Pertama, pendekatan menggunakan fungsi penilaian
(binding activity) untuk peringkat konformasi protein-ligan. Sistem ini
kemudian dimodifikasi oleh algoritma pencarian, dan penilaian yang sama
diterapkan lagi untuk merangking struktur baru. Kedua, penggunaan fungsi
tingkatan penilaian yaitu fungsi reduksi untuk mengarahkan pencarian dan
fungsi menentukan peringkat struktur yang dihasilkan (Kaapro dan Janne,
2002).
Docking molekular merupakan teknik penggabungan dua molekul
bersama-sama yang memiliki keunggulan, diantaranya bisa untuk mencari
model ikatan antara protein dan ligan, dapat memprediksi bentuk kompleks
intermolekuler struktur dua atau lebih molekul, kemungkinan menghasilkan
jumlah konformasi struktur yang banyak. Oleh karenanya, docking molekular
dapat mereduksi biaya pengembangan, meningkatkan efisiensi energi dan
daya guna lingkungan, serta menaikkan produktivitas dan keuntungan
(Pranowo & Abdul, 2011). Docking molekular dapat dilakukan menggunakan
perangkat lunak PyRx Autodock-Vina.
2. MCF-7 (Michigan Cancer Foundation-7)
Sel MCF-7 merupakan sel kanker payudara yang banyak digunakan
untuk penelitian. Sel ini memiliki karakteristik antara lain resisten agen
kemoterapi (Menchetner et al., 1998 & Aouali et al., 2003), mengekspresikan
reseptor estrogen (ER +), overekspresi Bcl-2 (Butt et al., 2000 & Amundson
et al., 2000) dan tidak mengekspresikan caspase-3 (Onuki et al., 2003 &
Prunet et al., 2005). Sel MCF-7 tergolong cell line adherent (Anonim, 2008)
yang mengekspresikan reseptor estrogen alfa (ER-α), dan resisten terhadap doksorubisin (Zampieri et al., 2002). Karakteristik lain dari MCF-7 adalah
mirip dengan Hep G2, dimana sel kanker jenis ini mengekspresikan wildtype
3. Bcl-2 (B-cell lymphoma-2)
Protein Bcl-2 merupakan protein yang memainkan peranan penting
pada regulasi apoptosis, atau kematian sel terprogram (Petros et al., 2010).
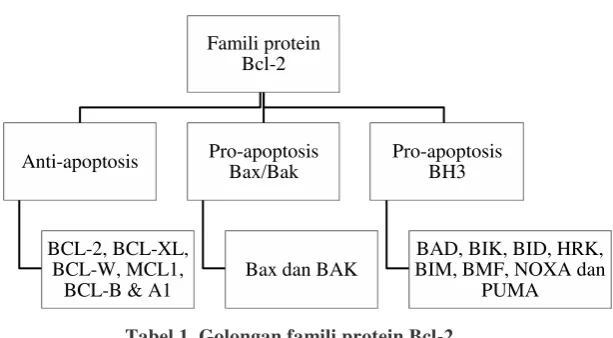
a. Golongan Bcl-2
Keluarga protein Bcl-2 dibagi menjadi tiga kelompok,
anti-apoptosis, pro-apoptosis anggota keluarga BAX/BAK dan pro-apoptosis
protein BH3 (Youle dan Stasser, 2008).
Tabel 1. Golongan famili protein Bcl-2
b. Mekanisme aksi Bcl-2
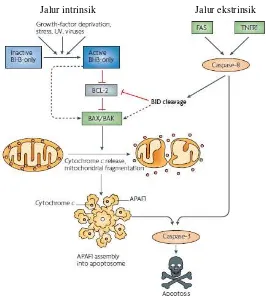
Proses apoptosis suatu sel terjadi melaui dua jalur, yaitu jalur
intrinsik (mitokondrial) dan jalur ekstrinsik (penghantaran sinyal secara
langsung melalui adapter). Dalam kondisi normal, anggota famili Bcl-2
pro-apoptosis dan anti-apoptosis beraktivitas secara seimbang. Protein
Bcl-2 anti-apoptosis berperan dalam proses apoptosis melalui jalur
intrinsik, sewaktu sel mengalami stress, protein ini berkurang jumlahnya
dalam mitokondria dan digantikan oleh protein pro-apoptosis seperti Bak,
Bax dan Bim. Kemudian, permeabilitas mitokondria meningkat, protein
cytochrom-c mengaktifkan cascade caspase. Cytochrom-c berikatan
dengan protein Apaf-1 (apoptosis activating factor-1) dan mengaktivasi
caspase-3, terjadilah apoptosis (Gambar 1). Famili protein
Bcl-2
Anti-apoptosis
BCL-2, BCL-XL, BCL-W, MCL1,
BCL-B & A1
Pro-apoptosis Bax/Bak
Bax dan BAK
Pro-apoptosis BH3
BAD, BIK, BID, HRK, BIM, BMF, NOXA dan
Gambar 1. Proses apoptosis sel melalui jalur intrinsik dan ekstrinsik (Youle dan Stasser, 2008)
Protein Bcl-2 merupakan antiapoptotik yang kuat dan ekspresi
protein Bcl-2 yang berlebihan meningkatkan ketahanan hidup sel (Marleen
et al., 2008). Pada sel kanker, mutasi dari gen Bcl-2 dapat menyebabkan
peningkatan ekspresi yang dapat menekan fungsi normal dari protein
proapoptosis. Jika terjadi pada protein tersebut, maka dapat menyebabkan
penurunan regulasi, sehingga sel kehilangan kemampuan untuk regulasi
apoptosis yang dapat memicu terjadinya kanker (Lumongga, 2008).
Apabila overekspresi Bcl-2 dihambat, maka proses apoptosis dapat
terjadi. Faktor transkipsi dari Bcl-2 adalah NFkB dari downstream
P13K/Akt. Terhambatnya ekspresi Bcl-2 ini selanjutnya akan menginduksi
pelepasan sitokrom c oleh mitokondria kemudian menginduksi jalur
caspase (Fitria et al., 2011).
Overekspresi Bcl-2 dapat dihindari dengan inhibitor Bcl-2 seperti
Bcl-xs yang diekspresikan dalam sel-sel MCF-7 dengan meningkatkan
chemosensitivity untuk VP-16 atau taksol (Sumantran et al., 1995). Jalur ekstrinsik
Penelitian terdahulu menunjukkan penemuan senyawa dengan molekul
kecil yang menghalangi fungsi dan dapat berikatan dengan hydrophobic
pocket Bcl-2 (Zheng et al., 2007).
4. Diketopiperazin
Diketopiperazin (2,5-piperazindion) merupakan model paling sederhana
untuk studi geometri cis-peptida dan untuk memeriksa pengaruh rantai
samping pada cincin konformasi. Diketopiperazin sejauh ini diselidiki
memiliki kandungan asam amino L- dan D- dan residu N-teralkilasi seperti
sarcosin, prolin, atau N-metil asam amino (Zeng et al., 2005). Aktivitas
diketopiperazin dikenal sebagai antitumor dan antiviral (Aoki et al., 2010 cit
Martin dan Carvalho, 2007), sitotoksik (Graz, 2004), antimikroba (Houston et
al., 2004) dan mempengaruhi saraf (Aoki et al., 2010 cit Prakash et al., 2002).
XR9051(N-(4-(2-(6,7-Dimetoksi-1,2,3,4,-tetrahidro-2-isoquinoli)etil)-fenil)-3-((3Z,6Z)-6-benziliden-1-metil-2,5-dioxo-3-piperaziniliden)
metil-benzamid), merupakan turunan diketopiperazin C-alkil yang memiliki
kemampuan melawan resistensi terhadap obat sitotoksik contohnya
doksorubisin dan etoposid, dengan jalan menghambat pengikatan
[3H]vinblastin dengan P-glikoprotein dan menghambat photoaffinity labeling
P-gp. Terhadap uji yang telah dilakukan, XR9051 lebih poten dibanding
siklosporin A dan verapamil (Dale et al., 1998). Stereokimia ring-junction
DKP menghambat pertumbuhan sel kanker dengan sintesis jalan homodimer
carboline dan dievaluasi pertumbuhan penghambatan terhadap sel kanker
PC-3 (prostat) dan NCI-H520 (paru-paru) untuk mengukur sitotoksisitas (Amy et
al., 2008). Turunan DKP C-alkil yang lain, diantaranya siklo-(4-S-hidroksi-R
-prolin-Risoleusin) (Ovenden et al., 2011),
3,6-Bis(indol-3-ilmetilen)piperazin-2,5-dion (Andreani et al., 2008) dan
1-Asetil-3-(3-tieniliden)piperazin-2,5-dion (Asiri, 2000). Turunan Diketopiperazin yang digunakan pada penelitian
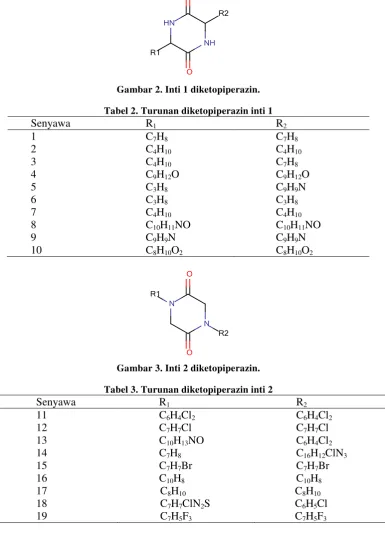
Gambar 2. Inti 1 diketopiperazin. Tabel 2. Turunan diketopiperazin inti 1
Senyawa R1 R2
1 2 3 4 5 6 7 8 9 10
C7H8
C4H10
C4H10
C9H12O
C3H8
C3H8
C4H10
C10H11NO
C9H9N
C8H10O2
C7H8
C4H10
C7H8
C9H12O
C9H9N
C3H8
C4H10
C10H11NO
C9H9N
C8H10O2
Gambar 3. Inti 2 diketopiperazin. Tabel 3. Turunan diketopiperazin inti 2
Senyawa R1 R2
11 12 13 14 15 16 17 18 19
C6H4Cl2
C7H7Cl
C10H13NO
C7H8
C7H7Br
C10H8
C8H10
C7H7ClN2S
C7H5F3
C6H4Cl2
C7H7Cl
C6H4Cl2
C16H12ClN3
C7H7Br
C10H8
C8H10
C6H5Cl
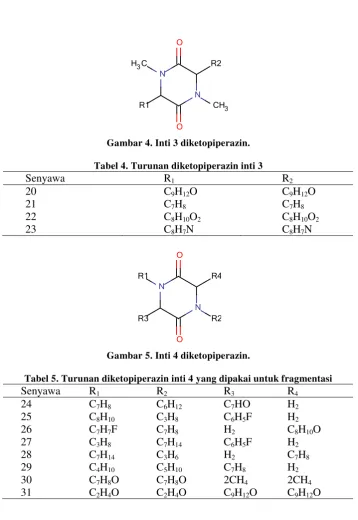
Gambar 4. Inti 3 diketopiperazin.
Tabel 4. Turunan diketopiperazin inti 3
Senyawa R1 R2
20 21 22 23
C9H12O
C7H8
C8H10O2
C8H7N
C9H12O
C7H8
C8H10O2
C8H7N
Gambar 5. Inti 4 diketopiperazin.
Tabel 5. Turunan diketopiperazin inti 4 yang dipakai untuk fragmentasi
Senyawa R1 R2 R3 R4
24 25 26 27 28 29 30 31
C7H8
C8H10
C7H7F
C3H8
C7H14
C4H10
C7H8O
C2H4O
C6H12
C3H8
C7H8
C7H14
C3H6
C5H10
C7H8O
C2H4O
C7HO
C6H5F
H2
C6H5F
H2
C7H8
2CH4
C9H12O
H2
H2
C8H10O
H2
C7H8
H2
2CH4
C9H12O
E. Keterangan empiris
Penelitian ini diharapkan dapat memperoleh turunan diketopiperazin yang
berpotensi memiliki interaksi dengan Bcl-2 pada sel MCF-7 secara docking