L
Journal of Experimental Marine Biology and Ecology 243 (2000) 185–208
www.elsevier.nl / locate / jembe
How costly is molluscan shell erosion? A comparison of two
patellid limpets with contrasting shell structures
*
Elizabeth G. Day , George M. Branch, Cecilia Viljoen
Coastal Ecology Unit, Zoology Department, University of Cape Town, Rondebosch 7701, South Africa Received 24 November 1997; received in revised form 6 July 1999; accepted 27 July 1999
Abstract
The energetic cost of replacing shell loss due to erosion has been overlooked in previous research on molluscan energy budgets, although several studies have shown that shell breakage incurs a significant short-term cost and may reduce growth. We measured the rate of shell erosion for two species of patellid limpets: Patella granatina, which has a shell predominated by calcite, and P. argenvillei, in which aragonite is dominant. Highest rates of erosion of P. granatina shells (36% p.a.) were recorded when the shells were grazed by a co-occurring limpet, P. granularis; in their absence, erosion was 15.2%. P. argenvillei experienced a slightly lower rate of erosion (32.4% p.a.) when its shells were grazed by juvenile conspecifics, whereas ungrazed shells lost 12.5%. P. argenvillei shells are often coated with an encrusting coralline, and infested by a boring lichen, but neither had a significant effect on the rate of erosion.
By modelling the energetic costs of shell erosion, we showed that the shell mass produced by P.
argenvillei each year is substantially greater than that produced by P. granatina, as is the loss of mass due to erosion. However, the energetic costs of shell production, and the losses due erosion, are higher for P. granatina, reflecting its faster growth and the higher energy content of its shells. For P. argenvillei, the cost of erosion increases with age from about 8–20% of the total energy devoted to ‘‘production’’ (somatic growth, shell production and gonadial output). For P. granatina it is 8–12%.
Experimental elimination of erosion failed to yield higher somatic, visceral or gonadial masses. This suggests that shell erosion does not carry a short-term cost equivalent to that experienced when shells are broken. Rather, it seems that compensation for shell erosion is an ongoing process involving a long-term cost. As a more explicit test of this hypothesis, we coated the shells of some individuals to protect them against erosion, left others unprotected, and coated one side of the shells of a third series of animals. Protected shells or sides of shells became thicker than those that were unprotected. Deposition of shell continued whether the shells (or portions of shells) were being eroded or not. Thus, compensation for shell erosion is an process of continual maintenance that is not simply initiated where and when erosion occurs, and will carry long-term costs.
2000 Elsevier Science B.V. All rights reserved.
*Corresponding author.
Keywords: Erosion; Shell structure; Limpet; Energy budget; Patella
1. Introduction
Most early studies on energy budgets for marine invertebrates focused primarily in tissue growth and reproduction (e.g., Grodzinski et al., 1975; Greenwood, 1979) but, more recently, attention has turned to mucus production (Davies et al., 1990; Niu et al., 1998), shell growth (Palmer, 1983) and shell repair in response to breakage (Vermeij et al. 1981; Blundon and Vermeij, 1983; Geller, 1990). In an examination of 15 species of prosobranch gastropods, Palmer (1983) has shown that such shell repair is energetically costly. For both bivalves (Peterson and Quammen, 1982) and gastropods (Geller, 1990) the coast of repair can be great enough to decrease reproductive effort. Intensified shell deposition associated with infestation of a sabellid worm on shells of Haliotis spp. also leads to stunted growth and loss of flesh mass (Culver et al., 1997)
One potentially important aspect of shell production that has been largely overlooked in research on molluscan energy budgets is the cost of replacing shell material lost by erosion. Patellid limpets are known to compensate for erosion by the deposition of inner shell layers (Branch, 1974), but the cost of this is unknown. Bioerosion of corals incurs substantial energetic costs (Sammarco et al., 1987).
If molluscs lose significant amounts of shell due to erosion, then it can no longer be presumed that shell deposition decreases with age and may eventually cease (as Branch, 1974, 1981 and Picken, 1980 assumed when calculating growth rates and life-time energy budgets for limpets). Molluscs may continually lay down shell to counter erosion. Indeed, such compensatory deposition may increase with age, perhaps even contributing to the commonly-observed pattern that incremental shell growth decreases with age. Thinning due to erosion may also make shells fragile, with potentially lethal conse-quences.
The vulnerability of a shell to erosion depends on several factors. Shell microstructure may influence its inherent resistance, although the exact significance of different micro-structural elements in molluscan shells has been much debated (Kennedy et al., 1969; Vermeij, 1978: Carter, 1980; Currey, 1980). External agents influencing erosion include wave action, sand scour and abrasion due to other animals boring into or grazing over molluscan shells (Lazar and Loya, 1991). Internal eroders of mollusc shells include lichens and blue-green algae (Webb and Korrubel, 1992; Espoz et al., 1995), while the external surfaces of limpet shells are often eroded by smaller limpets grazing on them (Eekhout et al., 1992). Limpet grazing is particularly powerful, due to their possession of hard, goethite-capped radular teeth (Branch, 1981), and may therefore significantly increase erosion rates.
In this study, two intertidal South African limpets, Patella granatina and P.
argenvillei, were used to assess rates of shell erosion and whether consequent shell
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 187
namely aragonite and calcite (Carter, 1980), aragonite being the weaker of the two (Currey and Taylor, 1974). In P. argenvillei the shell material is primarily aragonitic and very thick. The shell of P. granatina is thinner and composed mainly of calcite.
In this paper we address two primary question. The first regards the energetic cost of total shell reproduction (growth and maintenance to offset erosion) in relation to the investment of energy in growth and reproduction, and compares the relative costs of producing different types of shell. This leads to a second, and more fundamental, question—whether shell replacement to counter erosion entails short-term costs compar-able to those demonstrated for shell repair following damage by predators. If so, erosion should be acted upon when and where it occurs, and incur an immediate but short-lived energetic cost (which we refer to as ‘‘repair cost’’). Alternatively, erosion may be compensated by continual shell growth, which will carry an ongoing ‘‘maintenance cost’’. In this case, one would hypothesise that manipulation of erosion rates will have little effects on other elements of production. Shell maintenance would thus constitute a ‘‘safety factor’’ (sensu Lowell, 1985), forming a routine part of shell deposition whether or not erosion takes place.
Our experiments tested which of these competing hypotheses is most probable. More specifically, by undertaking manipulative field experiments on P. granatina and P.
argenvillei, we explored the following four issues: (1) Natural rates of erosion, and the
degree to which limpet grazing, lichen infestation and coralline cover affect erosion rates. (2) By protecting shells against erosion, we assessed whether shell maintenance is continual, or a short-term response that is initiated only when and where erosion occurs. (3) By modelling energy investment in gonadial output, shell and somatic growth, we estimated whether erosion has a significant energetic cost. (4) Finally, by experimentally eliminating erosion, we determined if this results in more energy being channelled into other aspects of production (i.e., changes in tissue or gonadial mass). If shell maintenance to combat erosion is a continual process, then even if erosion is eliminated, shell accretion should continue unabated and no extra energy will be available for growth or reproduction. Alternatively, if it is a flexible response initiated only when erosion occurs, then shell repair should stop with cessation of erosion, and any surplus energy may lead to increases in body or gonadial mass.
2. Materials and methods
2.1. Study sites and species
Patella granatina and P. argenvillei occur predominantly on the west coast of South
Africa, where they achieve densities and biomasses amongst the highest recorded anywhere for grazers (Bustamante et al., 1995). P. granatina is a mid-shore limpet that
22
achieves its highest densities (up to 184 m ) in sheltered bays where it can feed on drift kelp. It co-occurs with a smaller grazer, P. granularis, which is also abundant (up to 80
22
m ) and frequently grazes on its shells. Juveniles of P. granatina also graze on shells of adults, but their densities are low in comparison with those of P. granularis.
platforms and feeds by trapping live kelp (Bustamante et al., 1995; Bustamante and
22
Branch, 1996). It occurs in dense aggregations (as many as 198 m ; Bustamante et al., 1995) and its juveniles (defined here as individuals ,41 mm shell-length) almost
exclusively occupy and graze on adult shells (Eekhout et al., 1992). On average, juveniles constitute 24% (612% S.D.) of the population and, in dense assemblages 88%
(66% S.D.) occur on adult shells.
The experimental manipulations were carried out at two different areas in Namaqua-land on the West coast of South Africa. Work on P. granatina was conducted in 1990 at Rooiklippies (29842.819S; 1783.789E), a sheltered, boulder-strewn bay 200 km north of
Groenrivier that supports high densities of both P. granatina and P. granularis (Bosman et al., 1990). The experimental sites for P. argenvillei, set up during 1993, were located at Island Point, 16 km south of Groenrivier (30855.099S; 17837.899E), on a semi-exposed
rocky shore. Final work on both species took place in 1994 at Island Point.
2.2. Measurements of densities of grazers on shells
Densities of P. granularis grazing on P. granatina shells were assessed in May and August 1990. Since P. granularis grazes predominantly at night during low tide (Stephenson, in Thorpe, 1962), observations were done then. On each occasion, 20 quadrats (30360 cm) were randomly placed on the rock surface, and the numbers of
both P. granatina and P. granularis and their positions on rock or shells, were recorded. Because statistical analysis revealed no differences between sampling periods, data were pooled.
The area of limpet shell available for grazing was calculated by assuming shell area to be a cone, and describable by the formula: prl (Edwards and Penney, 1982), where
r5average shell radius and l5limpet height. The residual rock area available was
calculated as the area of the quadrat, less the total basal area occupied by limpet shells
2
(pr ).
Densities of juvenile P. argenvillei grazing on shells of adults were taken from existing data (Bosman et al., 1990).
2.3. Measurements of shell erosion and its effects
A combination of both live limpets and empty shells was used in separate manipulative caging experiments on both P. granatina and P. argenvillei. Empty shells were used to quantify erosion under different conditions, and live animals to test the effect of shell erosion on somatic mass and reproductive output. In all sets of experiments, cages consisted of square fences 4003400 mm, constructed from
alu-minium angle-iron rods, 40 mm high, and attached by rawl plugs and nails. These fences were placed in the zones of maximal limpet abundance and served to isolate the shells or live limpets and to either include or exclude molluscs known to graze on limpet shells.
2.3.1. Patella granatina
E
.G
.
Day
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
243
(2000
)
185
–
208
189
Table 1
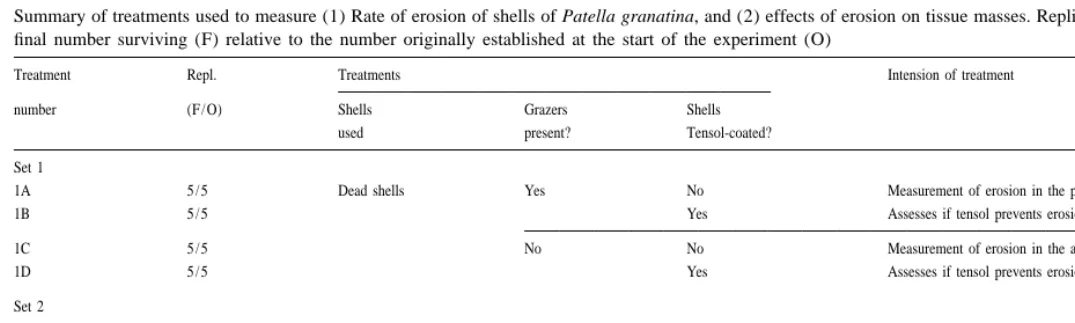
Summary of treatments used to measure (1) Rate of erosion of shells of Patella granatina, and (2) effects of erosion on tissue masses. Replicates (Repl.) indicate the final number surviving (F) relative to the number originally established at the start of the experiment (O)
Treatment Repl. Treatments Intension of treatment
number (F / O) Shells Grazers Shells
used present? Tensol-coated?
Set 1
1A 5 / 5 Dead shells Yes No Measurement of erosion in the presence of grazers
1B 5 / 5 Yes Assesses if tensol prevents erosion in presence of grazers
1C 5 / 5 No No Measurement of erosion in the absence of grazers
1D 5 / 5 Yes Assesses if tensol prevents erosion in the absence of grazers
Set 2
2A 5 / 5 Live limpets Yes No Measures effect of grazer-induced erosion on tissue mass
2B 5 / 5 Yes Control for 2A: Measures tissue mass when erosion is zero
2C 5 / 5 No No Measures effect of erosion on tissue mass in the absence of grazers
different treatments were established, divided between four sets of fences (replicated five times), as summarised in Table 1. The first four sets of treatments (1A–D, see Table 1) tested two things: whether erosion could be prevented by a coating of Tensol-16 cementing glue (1B, 1D), and the rates of erosion that take place on unprotected dead shells in the presence (1A) or absence (1C) of grazers that feed on shells (represented in this case by Patella granularis). Dead shells were used in this component of the work because erosion cannot be measured on live animals which have the capacity to replace eroded material.
]
Empty shells of a limited size range (x 57 mm, range 53–65 mm) were attached to the substratum using epoxy putty, after being dry-weighed, measured and individually labelled internally. The shells of animals in treatments requiring protection from erosion were painted with Tensol-16, a fast-drying adhesive glue which prevented further erosion of areas to which it adhered. Ten shells of P. granatina were used in each enclosure, half protected with tensol, half not. Half the enclosures contained grazers, i.e.,
P. granularis (n510–15 per plot, size range 30–40 mm), and half lacked them.
A second set of treatments (2A–D) were designed in parallel and were identical except that live Patella granatina were used instead of dead shells. These treatments tested whether erosion has an impact on somatic or gonadial mass, by comparing tensol-protected animals (2B, 2D) with unprotected animals (2A, 2C). By including (2A) or excluding (2C) grazers, their impacts could be assessed.
The experiment ran for three months, maintained monthly. Thereafter, both live animals and shells were removed and live animals were preserved in formalin. After fixation, bodies were removed and shell volumes established by measuring the volume of water they could hold. Body flesh was separated into gonad, viscera and muscle, and dry-weighed. These masses were expressed as a mass per unit shell volume to standardise for body size. Sizes of shells and live animals were homogeneous between treatments (Anova, F51.52, df57, p.0.05). Shells were weighed and measured, and
weight differences over time expressed as a percentage of the initial shell mass. Tensol was peeled off the protected shells before weighing.
2.3.2. Patella argenvillei
Nine treatments (each replicated four times) were established for P. argenvillei at Island Point in March 1993, and run for five months. The treatments are summarised in Table 2. Dead shells (Set 1) again served to measure erosion and live animals (Set 2) to determine if erosion has any effect on tissue or gonadial mass. Since access to the P.
argenvillei zone was only possible during very low spring tides, time did not permit us
to glue individual shells to the rock. Instead, empty shells were attached to asbestos plates with araldite resin, and these plates were then attached to the rock in the experimental fences. A minimum of eight shells per plot was used, all 61–75 mm in length.
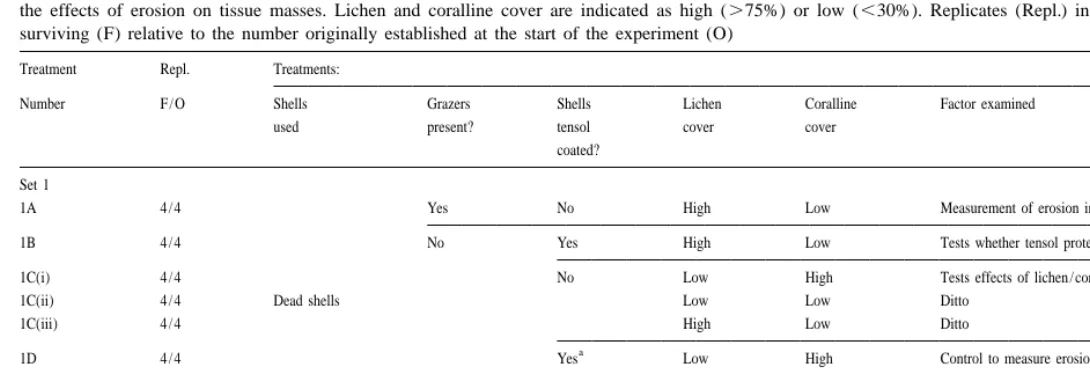
E
Summary of treatments used to measure (1) rates of erosion of shells of Patella argenvillei and the influence of grazers and lichen / coralline cover on erosion, and (2) the effects of erosion on tissue masses. Lichen and coralline cover are indicated as high (.75%) or low (,30%). Replicates (Repl.) indicate the final number
surviving (F) relative to the number originally established at the start of the experiment (O)
Treatment Repl. Treatments:
Number F / O Shells Grazers Shells Lichen Coralline Factor examined
used present? tensol cover cover
coated?
Set 1
1A 4 / 4 Yes No High Low Measurement of erosion in presence of grazers
1B 4 / 4 No Yes High Low Tests whether tensol protection prevents erosion
1C(i) 4 / 4 No Low High Tests effects of lichen / coralline on erosion
1C(ii) 4 / 4 Dead shells Low Low Ditto
1C(iii) 4 / 4 High Low Ditto
a
1D 4 / 4 Yes Low High Control to measure erosion of from coralline
Set 2
2A 4 / 4 Live limpets Yes No High Low Measures effects of grazer-induced erosion on tissue mass
2B 1 / 4 No Yes High Low Control for 2A: measures tissue mass when erosion is zero
2C 2 / 4 No High Low Effects of erosion on tissue mass in absence of grazers
a
effects of erosion on somatic and gonadial mass in the presence (2A) or absence (2B–C) of grazing by juveniles.
The design of these treatments followed the rationale and protocol outlined above for
P. granatina, but an additional factor applied to P. argenvillei: many of the shells were either covered with the encrusting coralline Spongites yendoi (which had the potential to reduce erosion by forming a protective layer), or were infested by the boring lichen
Pyrenocholema sp. (which potentially increases erosion). Three additional treatment
(1Ci–1Ciii) were therefore set up to establish their effects separately: dead shells were used to monitor erosion from shells that had combinations of high (.75%) lichen and
low (,30%) coralline cover, low lichen and high coralline cover, and low cover of both.
No shells had high cover of both. (All other treatments were standardised by using shells with high lichen and low coralline cover.) In measuring erosion from coralline-covered shells, there was an additional complication in that the shells might have lost weight both by shell erosion and by erosion from the coralline. To adjust for the latter, a separate control (1D) was introduced. Using shells with high coralline cover, tensol was applied to protect only those parts of shells that were not covered with coralline. This allowed measurement of coralline erosion alone. The percentage cover of coralline on each shell in all of the treatments was recorded, so coralline erosion could be calculated for each shell and subtracted from overall weight loss in other treatments to determine the actual losses due to shell erosion. As this control was used only as a correction factor, no results are reported for it.
Because of the plethora of factors investigated for P. argenvillei, it was logistically impossible to compare all possible combinations in an orthogonal design. Instead, the treatments were designed to compare the most critical combinations of factors (see Table 2).
After five months had elapsed, animals and shells were removed, dried, and the same variables measured as outlined for P. granatina (above).
2.4. Statistical analyses
Statistical analyses were performed using the SAS statistical package (SAS, 1987). A parametric three-way Anova was run on data for P. argenvillei shell treatments. The entire experimental design for P. granatina treatments, having two orthogonal factors (tensol-covered and uncovered; with and without grazers), and one nested factor, was analysed as a nested factorial design, as suggested by Underwood (1981). Where applicable, Tukey’s studentised range (HSD) a posteriori tests were used, with confidence limits set at 95%. Similar analyses were applied to the dead shells of P.
argenvillei. However, in the case of live P. argenvillei, several replicates were,
unfortunately, destroyed by a storm shortly before being collected. Thus, a non-parametric Kruskal-Wallis one-way analysis by rank was run between individual plots. This meant that replication in this instance was only at the level of individual shells within plots, and not between plots.
2.5. Compensation for erosion versus repair of shell damage
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 193
between March and October 1994 to test whether shell erosion is treated by the animals as a form of ‘‘damage’’ and repaired when it takes place, or whether compensation for erosion is continual.
For both species, three treatments were initiated, each involving 20 free-ranging, randomly-selected individuals, of standard length (40–45 mm P. granatina and 65–75 mm P. argenvillei). The treatments entailed (1) protecting entire shells by coating them with epoxy putty; (2) protecting one side of the shells with putty, while the other side was exposed to erosion; (3) control animals with unprotected shells.
We hypothesised that if the limpets treat shell erosion as a form of shell damage and repair it only when and where it occurs, there should be no differences in the thickness of shells that had been protected from erosion versus those that had not. (Only the shell halves that had been exposed to erosion should have been repaired in response, making them the same thickness as the protected halves.) If, on the hand, shell material is being continually laid down to compensate for erosion, then shells (or portions of shells) that were protected from erosion should become thicker than those that were not.
At the conclusion of the experiment, the body contents were removed from the shells, and shell length and height measured. Shells were then cut in half, and their thickness measured mid-laterally at three corresponding points on each shell half: close to the apex, just above the muscle attachment scar, and 0.5 cm above the base of the shell.
The means of these three measurements were then compared between sides of shells and between treatments. Left and right hand sides were compared within treatments, to check for intrinsic differences between sides of shells, before treatments were compared.
2.6. Energy budgets
The organic contents of shells of P. argenvillei and P. granatina were measured using the Trichloroacetic acid (TCA) method (Goulletquer and Wolowicz, 1989). This method purports to yield a better estimate than the use of HCl to dissolve shell material, or simple combustion of the whole shell (e.g., Griffiths and King, 1979). The peripheral (younger) portions of shells were used, to avoid biases due to erosion (Palmer, 1983). Five replicates were analysed.
The calorific values of foot muscle and female gonads were measured using a CP500 automatic bomb calorimeter, calibrated with benzoic acid. Six replicate samples of dried tissue from both limpet species were individually ground, and samples of approximately 0.25 g were analysed. The energy value used for shell organic material was taken as 17
21
kJ g , based on the mean value found for shell organic material in five Patella species (unpublished data). This value lies very close to that found by Goulletquer and Wolowicz (1989) for other marine molluscs.
2.7. Models of energy allocations
To compute the importance of erosion in the energy budgets of P. granatina and P.
argenvillei, annual energy budgets were calculated for a range of sizes (35–62 mm for
P5Pg1Pr1Ps1Pe
where P5total annual production; Pg5somatic growth; Pr5reproductive output, Ps5
incremental shell growth, and Pe5shell deposition to compensate for erosion.
To simplify the number of variables, calculations were confined to females only, and the rates of erosion used were those found for limpets in grazed shell treatments. To estimate energy expenditure on growth and reproduction, the following regressions were extracted from Bosman et al. (1990), with growth rates being taken from Raubenheimer (1991).
Length (L, mm) vs. dry body mass (M, g):
26 3.2
P. granatina M5(3.5310 )3L (1)
26 3.38
P. argenvillei M5(1.26310 )3L (2)
Length (L, mm) vs. dry shell mass (S, g):
25 3.28
P. granatina S5(2.1310 )3L (3)
25 3.45
P. argenvillei S5(1.38310 )3L (4)
21
Growth, i.e., increase in shell length (G, mm y ):
P. granatina G5(20.3413L )121.4 (5)
P. argenvillei G5(20.2073L )117.07 (6)
Annual gonadial output (dry mass, g):
P. granatina: 60% of body weight, after reproductive age (Branch, 1974) (7)
P. argenvillei: since this species exhibits a clear spawning peak in late May, gonad output was calculated as the difference between May and October gonad mass (immediately prior to, and after completion of, spawning), using the following regressions, where Lm refers to shell length at mid-year.
213 6.619
Wet mass of female gonad (May)59.908310 3Lm (8)
210 4.467
Wet mass of female gonad (October)51.607310 3Lm (9)
Wet mass (WW) of female gonadial output was converted to dry mass (DW) by:
DW50.41 WW (60.02 S.D.)
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 195
this species at our research site (Bosman et al., 1990), but is problematic due to the apparent existence of multiple spawnings there, which probably inflate the estimate. Both values lead to the same overall conclusions, so we preferred the more conservative estimate for Kommetjie.
3. Results
3.1. Incidence of grazing on shells
Shells of P. granatina were frequently grazed by P. granularis. Night-time densities of
P. granularis on P. granatina shells were 0.33 (S.D.60.18) per shell in May, and 0.40
(60.17) in August. A marked preference was exhibited by P. granularis for grazing on
P. granatina rather than on rock. Expressed per square meter of substrate, 185 (681
S.D.) grazed on shells of P. granatina, but only 41 (626 S.D.) on rock (Anova:
F596.19; df574; p,0.001). Data were pooled from the two sampling sessions, as no
statistical differences were found between them (Anova: F51.327; df578; p.0.05).
A large proportion (88%) of juveniles of P. argenvillei juveniles live and graze on shells of adults, and 92% of all adults have at least one juvenile on their shells (Bosman et al., 1990). The frequency of grazing on adult shells is thus likely to be high. Conclusive evidence that the limpets do graze on, and erode, the shells of both P.
granatina and P. argenvillei was provided by the frequent radular scrape-marks on their
shells—often cutting through the outer shell layers.
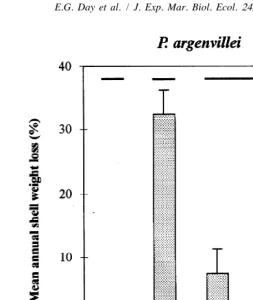
3.2. Rates of shell erosion
Rates of erosion for dead P. granatina shells, extrapolated to an annual total, are shown in Fig. 1. Practically no erosion occurred from tensol-coated shells, irrespective of whether they were grazed or not, showing that this treatment effectively eliminates erosion. Unprotected shells exposed to grazing exhibited the highest rates of erosion (36% per annum), but even ungrazed, unprotected shells eroded 15.2% (Fig. 1). Both protection (i.e., tensol-coating vs. no coating) and the presence or absence of grazers had significant effects on the percentage loss of shell mass, as did their interaction (Two-way ANOVA: Protection, F578.4, df51, p,0.001; Grazers, F59.72, df51, p,0.0025;
Interaction, F512.39, df51, p,0.0007). Tukey’s test indicated significant differences
between all four treatments except the two tensol-treated lots of shells (Fig. 1). For P. argenvillei, highest erosion rates were experienced by shell treatments with grazers (32.4% per annum), while ungrazed shells with comparable lichen and coralline cover lost about 12.5% (Fig. 2). A noticeable effect of excluding grazers from some shell treatments was the rapid growth of macro-algae on these shells. Results of the Anova, testing the effects of different treatments and shell length on weight loss, showed that only the treatments had significant effects ( p,0.05). Thus differences between
Fig. 1. Mean (1S.D.) rates of shell erosion from shells of dead P. granatina that were protected or unprotected by a coating of tensol, in the presence (1) or absence (2) of grazers. Horizontal lines above the
bars indicate those groups of results that were significantly different from one another (Tukey’s test, p,0.05).
differences detected between treatments with contrasting amounts of lichen and coralline cover.
3.3. Effect of erosion on tissue mass
Tissue masses for P. granatina are shown in Table 3. Two-way Anovas revealed no significant differences between any treatments for any tissue type ( p.0.05). Thus,
whether the limpets were tensol-protected or not, or exposed to grazers or not, made no difference to their tissue masses.
Similar results were obtained for P. argenvillei. A Kruskall-Wallis one-way analysis by rank of P. argenvillei tissue masses revealed no significant differences between any treatments, at the 95% confidence interval. The data are illustrated in Table 3, and again show that tissue masses were not influenced by the presence or absence of grazers, or by tensol-protection preventing erosion.
3.4. Energy budgets for shell production
The organic content (AFDW) of shells of P. argenvillei (1.75%60.31 S.D.) was found
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 197
]
Fig. 2. Rates of erosion from shells of dead Patella argenvillei (x1S.D.). Data were pooled between plots
after Anova failed to detect significant difference between them ( p,0.05). Horizontal lines above the bars
indicate the three groups of results that were significantly different from one another (Tukey’s test, p,0.05).
Table 3
Gonadial, visceral, foot and total flesh masses of (a) Patella granatina and (b) P. argenvillei. The data (means6S.D.) are standardised as mass per unit of internal shell volume, and shown for different treatments involving the provision of protection against erosion (protected vs. unprotected) and the presence (1) or absence (2) of grazers
Species Tissue Tissue mass (g / unit volume) for treatments:
Protected, Protected, Unprotected Unprotected
grazers1 grazers2 grazers1 grazers2
P. granatina Gonad 0.00860.007 0.01260.007 0.00760.007 0.01860.036
Viscera 0.02060.003 0.02260.006 0.01960.007 0.03060.048
Foot 0.04860.009 0.04560.016 0.04360.010 0.06460.082
Total 0.07660.012 0.08960.011 0.06960.017 0.11260.018
P. argenvillei Gonad – 0.00360.001 0.00460.001 0.00360.001
Viscera – 0.01760.000 0.01960.000 0.01560.000
Foot – 0.06760.010 0.06960.016 0.06560.012
Table 4
Energy values of tissues of P. argenvillei and P. granatina
Tissue Mean calorific content AFDW: DW conversion
21
(kJ g DW,6Std. dev.) (6Std. dev.)
Foot muscle:
P. argenvillei 19.97 (60.32) 0.957 (60.083)
P. granatina 16.12 (60.81) 0.856 (60.071) Female gonad:
P. argenvillei 24.86 (61.14) 0.948 (60.080)
P. granatina 27.58 (61.22) 0.856 (60.072)
the shell and tissues of the two species are shown in Table 4. These values were used in the energetic models developed below in the discussion.
Using the regressions cited earlier, erosion values, and the energetic values obtained for shell organic components, Fig. 3 compares shell production in P. granatina with that in P. argenvillei, across a range of shell lengths. P. argenvillei lays down considerably more shell mass, and loses more to erosion, than does P. granatina (Fig. 3a). However, energetic expenditure by P. granatina on shell growth is consistently greater than in P.
argenvillei, and the energetic costs of its shell erosion are also higher (Fig. 3b). These
differences between the species reflect, in part, the faster growth rate of P. granatina (Branch, 1974; Eekhout et al., 1992), but also the higher organic content of P. granatina shells.
3.5. Compensation for erosion, versus repair of shell damage
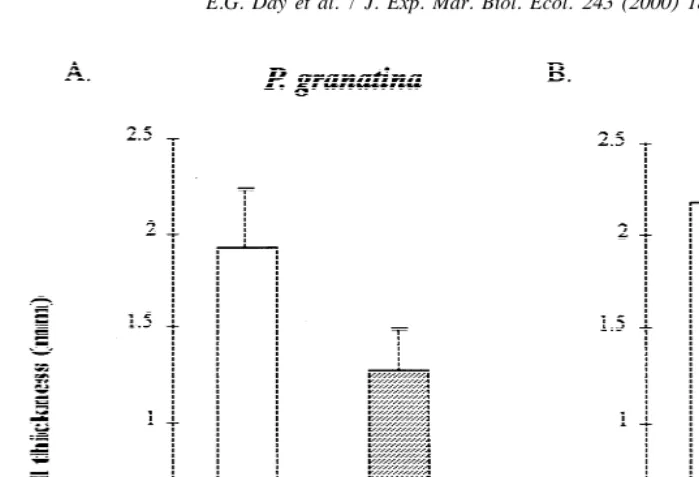
Shells of both P. granatina and P. argenvillei became thicker when protected with putty, relative to unprotected shells (Fig. 4a,b), although the magnitude of the difference was only statistically significant in the case of P. argenvillei (Newman-Keuls a
posteriori test, p,0.05). For both species, there were no significant intrinsic differences
between left and right halves of shells (paired t5tests, t always ,2.262; df510).
calc
Because of this, direct comparisons could be made between the two sides of individual shells that were ‘‘half-puttied’’, i.e., one side was protected with putty while the opposite side was left unprotected (Fig. 5). For both species, puttied sides were significantly thicker than unputtied sides (paired t-test, df518, p,0.001).
4. Discussion
4.1. Rates of erosion and factors effecting erosion
The shells of both P. argenvillei and P. granatina are regularly grazed by smaller limpets; mainly conspecific juveniles on P. argenvillei, and P. granularis in the case of
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 199
Fig. 3. Annual investment by Patella granatina and P. argenvillei in overall shell growth and in compensation for erosional losses, expressed in terms of (a) shell mass, and (b) energy expenditure.
considers the excavating power of patellogastropod limpets (Runham et al., 1969; Branch, 1981; Creese and Underwood, 1982; Steneck and Watling, 1982; Steneck, 1982).
Fig. 4. Mean (1S.D.) shell thickness achieved by (a) Patella granatina (b) and P. argenvillei, in relation to whether their shells were protected by putty or not. Data are shown separately for left and right halves of shells.
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 201
Fig. 5. Comparisons of the mean (1S.D.) thickness achieved by (a) Patella granatina and (b) P. argenvillei when one half of their shells was protected from erosion by putty.
additional protection imbued by coralline cover, or extra erosion associated with the lichens, could have been masked. Differences in erosion related to coralline or lichen cover might have been more obvious in the presence of grazers. In addition, because of the peculiar shell structure of P. argenvillei, with differential orientation of calcium carbonate crystals in different layers (Currey, 1980), the lichen does not usually bore through more than one layer of shell. This may also reduce its impact.
4.2. Energy budgets and costs
The rates of shell erosion in both P. granatina and P. argenvillei (36% and 32% of total shell mass annually), under near-natural conditions where they are grazed, suggests a significant cost, which rises proportionally as the animals age and incremental shell growth declines (Fig. 3). Such potential costs were, however, not evidenced in the experiments on live limpets, which failed to increase their tissue masses when erosion was prevented (Table 3). What then are the actual costs of erosion?
Fig. 6. Models of the proportional investment of energy by (a) Patella granatina and (b) P. argenvillei into shell growth, compensation for shell erosion, somatic tissue growth and reproductive output, expressed in relation to body size. The data were derived from the equations in the Methods and the Results in Table 4.
increasing with age. Thus, erosion is energetically costly but, nonetheless, the implica-tions of this need to be re-interpreted.
If compensation for shell-erosion occurs at the expense of other aspects of production such as somatic growth and reproduction, one would predict that as the erosion of shell increases with age, correspondingly greater proportions of energy will be devoted to shell generation. In fact, the proportion of energy allocated by P. argenvillei to shell production actually decreases from about 22% of overall production at a size of 30 mm to about 12% at 80 mm (Fig. 6), and remains more or less constant in P. granatina. For large limpets, incremental shell growth is negligible. Shell deposition, although subsequently eroded, is however continual. Thus, rather than imposing a cost in terms of energy foregone, erosion merely detracts from incremental growth. The notion of an energetic trade-off between shell growth and other functions (Grahame and Branch, 1985) thus falls away.
The results of these experiments and calculations appear at first sight to contradict the work of Vermeij (1978), Palmer (1981, 1983) and Geller (1990), who measured the costs of shell repair in terms of reduced growth rates. However, we argue below that damage-induced shell-repair a fundamentally different process from that due shell erosion, and that the energetic costs incurred by shell erosion are only measurable as an ongoing response, which is not equivalent to the short-term reaction to shell breakage, which is associated with reduced growth.
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 203
these experiments, it has been concluded that repair is energetically costly. Equating these costs with those of erosion assumes that the two phenomena must be similar. In practice, however, predator-induced damage is usually a sudden, localised effect on a shell, often extensive enough to penetrate right through the shell matrix (Geller, 1990). By contrast, erosion is a continual process acting over the entire area of a shell. Its effects at any one moment are therefore probably slight, and only gain stature as a cumulative effect over time. Only if erosion is extreme enough to result in shell breakage will it be detectable in the same manner as predator-damage, and elicit short-term shell repair. Thus, only in its extremes can shell erosion approximate predatory damage. Observations of the interior of P. argenvillei shells reinforce this idea, in that visible scarring is only obvious beneath shell fractures, while the rest of the shell has an evenly-laid lining, regardless of how eroded it may be in places. This suggests that erosion is offset by a steady deposition of shell material that is spread over the whole shell and is independent of erosion.
Thus, shell lost through erosion, by virtue of its gradual and continual nature, will not stimulate a short-term increase in the rate of shell deposition. Limpets lay down shell continually (Branch, 1974; Cohen and Branch, 1992) and (Littorina littorea) deposit shell at a constant rate (Kemp and Bertness, 1984). Simultaneously, layers from the outside are eroded away.
The results of the experiments in which putty was used to protect shells from erosion add substantial weight to this argument. If the animals were responding differentially to different degrees of erosion of their shells, one would expect both sides of the ‘‘half puttied’’ shells to be of equal thickness, with repair being restricted to the eroded portions only, and the uneroded portions remaining a constant thickness. Such localised repair might carry a short-term cost, diverting energy from other components of production. Instead, as one would predict from the ‘‘continual growth’’ hypothesis, the protected sides of the shells became relatively thicker than the other side, continual shell accretion adding to their thickness even although they were not being eroded away (Fig. 5). Similarly, shells that were completely protected from erosion became thicker than unprotected shells (Fig. 4), indicating that there is no ‘‘saving’’ of energy associated with reduced erosion, and thus no surplus that can be channelled to other functions.
A closer look at the models in Fig. 6 indicates that, far from increasing shell production as erosion increases with size, total allocations to shell production actually decrease or, at best, remain constant, while increasing effort is devoted to reproductive output. Initially, this appears counter-productive. If, in large animals, erosion exceeds shell regeneration, which is implied by the upper limits of Fig. 6, the shell will gradually thin, and likelihood of fracture will increase. At this point, one would expect higher allocations of energy towards shell production to be vital.
into gonad production. Geller (1990) noted that molluscs in general increase reproduc-tive effort as risks of mortality increase, and Branch (1981) records the same response more specifically for two species of Patella.
The theory so far advanced is that the short-term costs associated with erosion are low, because the rates of shell regeneration are fixed. What, then, are these maintenance costs ?
4.3. Maintenance costs
Regardless of the fact that there appear to be no short-term trade-offs between rates of erosion versus energy for reproduction or somatic growth, it remains a fact that the annual rates of erosion experienced by P. granatina and P. argenvillei, 36% and 32% respectively, must represent a cost to the animals, in terms of energy foregone for other functions, although Palmer (1983) suggests this is only significant if the costs of shell generation are high.
Gastropod shells comprise two principal components: an organic matrix, which interlaces a framework of calcium carbonate crystals (Goulletquer and Wolowicz, 1989). Of these, the organic matrix is metabolically the more demanding (Palmer, 1983). Costs
21
of calcium accretion are as low as 1–2 J mg CaCO produced (Palmer, 1992). This is3
21
low when compared to the calorific value of organic matrix (17 J mg ). Thus, in assessing the significance of shell erosion in the two limpet species, it is the organic component of the shells that will be most informative. Differences in proportions of organic matrix (3.58% for P. granatina, as opposed to 1.75% for P. argenvillei) suggest functional differences between the shells of the two species.
When the energy allocations to shell growth are compared between P. argenvillei and
P. granatina (Fig. 3b), they confirm that the higher organic content of P. granatina shells imposes a higher cost of shell production than for P. argenvillei. Fig. 3a, by contrast, compares the mass of shell produced and eroded in each species. Not surprisingly, given the high cost of producing the organic matrix, considerably less shell mass is produced in P. granatina, a trade-off suggested by Currey and Taylor (1974). These differences prompt one to search for a possible functional significance in having more or less organic matrix.
4.4. Possible functional differences between shell types
The difference in organic content between the two species reflects their shell microstructure, with P. granatina containing a higher proportion of the calcitic CaCO3
morph than the aragonitic morph, which has a much lower proportion of organic material (Carter, 1980). P. argenvillei shells are composed primarily of aragonite.
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 205
Patella granularis, the calcite content of the shell rises as temperature drops (Cohen and
Branch, 1992).
The energetically cheaper aragonite has one advantage in that large quantities of shell can be produced at little cost (Currey, 1980). Indeed, the aragonitic shells of P.
argenvillei are substantially heavier, mass per length, than those of P. granatina. For
low-shore dwellers, there are particular benefits associated with a thick, heavy shell. Stanley (1988) suggests that where wave action is likely to be a problem, a heavy shell confers extra stability. This would not be required by the higher-shore P. granatina, which, moreover, inhabits primarily sheltered areas (Bustamante et al., 1995). The generality of this can be assessed by considering South African patellids as a whole. Six out seven high- to mid-shore species have light, calcite-predominated shells, whereas the shells of six out of eight low-shore and subtidal species are heavy and predominantly aragonitic (Ridgway et al., 1998). It is also a general rule that those species with heavy, aragonitic shells have high powers of tenacity but, as an apparent trade-off, are relatively immobile compared to species with light, calcitic shells (Branch and Marsh, 1978).
Another potential advantage of aragonite is the claim that it is more resistant to abrasion than calcite (Taylor and Reid, 1990). Although P. granatina and P. argenvillei are both abraded by small grazers, P. argenvillei loses greater absolute amounts of shell in the process (Fig. 3a). Thus, an energetically cheaper, less erosive shell composition minimises energetic losses.
However, the higher proportion of organic matrix associated with calcite means that it is less likely to crack, forces being diverted into shearing, the energy of which is quickly dissipated across the matrix (Currey and Taylor, 1974). For mid-shore limpets, exposed to potentially high rates of predation from birds (Hockey and Branch, 1984; Branch et al., 1987), an ability to resist cracking caused by sharp blows may be advantageous.
There is, of course, an alternative manner of looking at the functional significance of shell microstructure in relation to erosion. The possession of a particular shell type may have a phylogenetic explanation, rather that being an adaptive response to circum-stances. A recent cladistic analysis of the world’s patellid limpets revealed four clades (Ridgway et al., 1998). Of those in South Africa, all members of the Scutellastra clade (including P. argenvillei) have predominantly aragonitic shells, while those of the
Cymbula clade (including P. granatina) and the Helcion clade have aragonitic shells.
Clearly, these hypotheses present a cause and effect conundrum, which must remain purely theoretical. In any event, however, rates of shell accretion have to match rates of erosion for the animals to survive at all, and all the evidence points to this being an continual process of maintenance.
5. Conclusion
rate of erosion to which a species is likely to be exposed. As such, they are much more akin to the safety factors built into shell strength to cater for extreme events (Lowell, 1985), than to emergency repairs following damage and involving short-term energetic trade-offs between different aspects of production.
Acknowledgements
This research was supported by a research grant from the South African Network for Coastal and Oceanographic Research (SANCOR) and the Foundation for Research Development (FRD) to G. M. Branch, and bursaries to E. Day and C. Viljoen. We also appreciate assistance with field work from S. Eekhout and R. Bustamante, constructive discussions with S. Eekhout and statistical advice from Rob Day. [AU]
References
Blundon, J.A., Vermeij, G.J., 1983. Effect of shell repair on shell strength in the gastropod Littorina irrorata. Marine Biology 76, 41–45.
Bosman, A.L., Eekhout, S., Branch, G.M., 1990. The exploitation potential of limpets on the coast of Namaqualand. Final report to the Chief Director, Marine Development, Sea Fisheries Research Institute, Cape Town.
Branch, G.M., 1974. The ecology of Patella Linnaeus from the Cape Peninsula, South Africa. 3. Growth Rates. Transactions of the Royal Society of South Africa 41, 111–160.
Branch, G.M., 1981. The biology of limpets: physical factors, energy flow and ecological interactions. Oceanography and Marine Biology an Annual Review 19, 235–380.
Branch, G.M., Barkai, A., Hockey, P.A.R., Hutchings, L., 1987. Biological interactions: causes or effects of variability in the Benguela ecosystem? South African Journal of Marine Science 5, 425–445.
Branch, G.M., Marsh, A.C., 1978. Tenacity and shell shape in six Patella species: adaptive features. Journal of Experimental Marine Biology and Ecology 34, 111–130.
Bustamante, R., Branch, G.M., 1996. The dependence of intertidal consumers on kelp-derived organic matter on the west coast of South Africa. Journal of Experimental Marine Biology and Ecology 196, 1–28. Bustamante, R., Branch, G.M., Eekhout, S., 1995. Maintenance of an exceptional intertidal grazer biomass in
South Africa: subsidy by subtidal kelps. Ecology 76, 2314–2329.
Carter, J.G., 1980. Environmental and biological controls of bivalve shell mineralogy and microstructure. In: Rhoads, D.C., Lutz, R.A. (Eds.), Skeletal Growth of Aquatic Organisms, Vol. 1, Plenum, London, pp. 52–96.
Cohen, A.L., Branch, G.M., 1992. Environmentally controlled variation in the structure and mineralogy of
Patella granularis shells from the coast of southern Africa: implications for paleotemperature assessments.
Palaeogeography, Palaeoclimatology and Palaeoecology 91, 49–57.
Creese, R.J., Underwood, A.J., 1982. Analysis of inter- and intra-specific competition amongst intertidal limpets with different methods of feeding. Oecologia (Berl.) 53, 337–346.
Culver, C.S., Kuris, A.M., Beede, B., 1997. Identification and management of exotic sabellid pests in California abalone aquaculture. California Sea Grant College Programme. Publication No. T-041. Currey, J.D., 1980. Mechanical properties of Mollusc shells. In: Symposia of the Society for Experimental
Biology, No. XXXIV. The Mechanical Properties of Biological Materials. Cambridge University Press, Cambridge, pp. 75–97.
E.G. Day et al. / J. Exp. Mar. Biol. Ecol. 243 (2000) 185 –208 207 Davies, M.S., Hawkins, S.J., Jones, H.D., 1990. Mucus production and physiological energetics in Patella
vulgata L. Journal of Molluscan Studies 56, 1–5.
Edwards, C.H. Jr., Penney, D.E., 1982. Calculus and Analytic Geometry. Prentice-Hall Inc., Englewood Cliffs, New Jersey.
Eekhout, S., Raubenheimer, C.M., Branch, G.M., Bosman, A.L., Bergh, M.O., 1992. A holistic approach to the exploitation of intertidal stocks; limpets as a case study. South African Journal of Marine Science 12, 1017–1029.
Espoz, C., Guzmann, G., Castilla, J.C., 1995. The lichen Thelidium litorale on shells of intertidal limpets: a case of lichen-mediated cryptic mimicry. Marine Ecology Progress Series 119, 191–197.
Geller, J.B., 1990. Reproductive responses to shell damage by the gastropod Nucella emarginata (Deshayes). Journal of Experimental Marine Biology and Ecology 136, 77–87.
Goulletquer, P., Wolowicz, M., 1989. The shell of Cardium edule, Cardium glaucum and Ruditapes
philippinarum: organic content, composition and energy value, as determined by different methods. Journal
of the Marine Biological Association, United Kingdom 69, 563–572.
Grahame, J., Branch, G.M., 1985. Reproductive patterns of marine invertebrates. Biological Annual Review 23, 373–398.
Greenwood, P.J., 1979. Growth, respiration and tentative energy budgets for two populations of the sea urchin
Parechinus angulosus (Leske). Estuarine and Coastal Marine Science 10, 347–367.
Griffiths, C.L., King, J.M., 1979. Energy expended on growth and gonad output in the ribbed mussel
Aulacomya ater. Marine Biology 53, 217–222.
Grodzinski, W., Klelowski, R.Z., Duncan, A., 1975. Methods for ecological bioenergetics—International Biological Programme handbook No. 24. Blackwell Scientific Publications, Oxford.
Hockey, P.A.R., Branch, G.M., 1984. Oystercatchers and limpets: impact and implications. A preliminary assessment. Ardea 26, 199–206.
Kemp, P., Bertness, M.D., 1984. Snail shape and growth rates: evidence for plastic shell allometry in Littorina
littorea. Proceedings of the National Academy of Sciences, USA 81, 811–813.
Kennedy, W.J., Taylor, J.D., Hall, A., 1969. Environmental and biological controls on bivalve shell mineralogy. Biological Reviews 44, 499–530.
Lazar, B., Loya, Y., 1991. Bioerosion of coral reefs—a chemical approach. Limnology and Oceanography 36 (2), 377–383.
Lowell, R.B., 1985. Selection for increased safety factors of biological structures as environmental unpredictability increases. Science, New York 228, 1009–1011.
Niu, C.-J., Nakao, S., Goshima, S., 1998. Energetics of the limpet Lottia kogamogai (Gastropoda: Acmaeidae) in an intertidal rocky shore in southern Hokkaido, Japan. Journal of Experimental Marine Biology and Ecology 224, 167–181.
Palmer, A.R., 1981. Do carbonate skeletons limit the rate of body growth? Nature 292, 150–152.
Palmer, A.R., 1983. Relative cost of producing inorganic matrix versus calcification: evidence from marine gastropods. Marine Biology 75, 287–292.
Palmer, A.R., 1992. Calcification in marine molluscs: how costly is it? Proceedings of the National Academy of Sciences, USA 89, 1379–1382.
Peterson, C.H., Quammen, M.L., 1982. Siphon nipping: its importance to small fishes and its impact on growth of the bivalve Protothoca staminea (Conrad). Journal of Experimental Marine Biology and Ecology 63, 249–268.
Picken, G.B., 1980. The distribution, growth and reproduction of the antarctic limpet, Nacella(Patinigera)
concinna (Strebel, 1908). Journal of Experimental Marine Biology and Ecology 42, 71–85.
Raubenheimer, C.M., 1991. Length-based simulation model for assessing the exploitation potential of the limpet, Patella granatina. Unpublished M.Sc. thesis. University of Cape Town, Cape Town, South Africa. Ridgway, S.A., Reid, D.G., Taylor, J.D., Branch, G.M., Hodgson, A.N., 1998. A cladistic phyllogeny of the
family Patellidae (Mollusca: gastropoda). Phil. Trans. Roy. Soc. Lond. B. 353, 1645–1671. Runham, P.R., Thornton, W., Dhaw, D.H., Wayte, R.C., 1969. Z. Zell. Forsch. 99, 608–626.
Sammarco, P.W., Risk, M.J., Rose, C., 1987. Effects of grazing and damselfish territoriality on internal bioerosion of dead corals: indirect effects. Journal of Experimental Marine Biology and Ecology 112, 185–199.
Stanley, S.M., 1988. Adaptive morphology of the shell in bivalves and gastropods. In: Trueman, E.R., Clamie, M.R. (Eds.), The Mollusca, Vol. 11, Chap. 5, Academic Press, Inc, pp. 296–345.
Steneck, R.S., 1982. A limpet-coralline alga association: adaptations and defences between a selective herbivore and its prey. Ecology 63 (2), 507–522.
Steneck, R.S., Watling, L., 1982. Feeding capabilities and limitations of herbivorous molluscs: a functional approach. Mar. Biol. 68, 299–319.
Taylor, J.D., Reid, D.G., 1990. Shell microstructure and mineralogy of the Littorinidae: ecological and evolutionary significance. Hydrobiologia 193, 199–215.
Thorpe, W.H., 1962. Learning and instinct in animals. Methuen, London, pp. 558.
Underwood, A.J., 1981. Techniques of analysis of variance in experimental marine biology and ecology. Oceanography and Marine Biology an Annual Review 19, 513–605.
Vermeij, G.J., 1978. Biogeography and Adaptation: Patterns of Marine Life. Harvard University Press, Cambridge, Massachusetts.
Vermeij, G.J., Schindel, D.E., Zipser, E., 1981. Predation through geological time: evidence from gastropod shell repair. Science, New York 214 (27), 1024–1026.
Watabe, N., 1983. Shell repair. In: Saleuddin, S.M., Wilbur, K.M. (Eds.), The Mollusca, Vol. 4, Ch. 7, Academic Press, Inc, pp. 280–327.