AMELINDA IRENA
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2010
ABSTRAK
AMELINDA IRENA. Isolasi dan Optimasi Protease Bakteri Termofilik dari
Sumber Air Panas Tangkuban Perahu Bandung. Dibimbing oleh I MADE
ARTIKA dan ERNY YUNIARTI.
Enzim protease merupakan enzim yang banyak digunakan dalam bidang
industri pangan. Bakteri termofilik dapat menghasilkan enzim protease yang dapat
bertahan pada suhu tinggi. Penelitian ini bertujuan mengisolasi bakteri termofilik
dan optimasi pH dan suhu protease pada isolat yang memiliki indeks proteolitik
terbesar.
Isolasi dilakukan terhadap 33 isolat bakteri termofilik dari tiga sumber air
panas. Isolasi dilakukan dengan menggunakan metode cawan tuang. Aktivitas
protease diukur dengan metode Bergmeyer, dan karakterisasi enzim protease
dilakukan pada pH 6-10 dan suhu 50
oC-90
oC. Hasil penelitian menunjukkan
bahwa dua isolat dari sumber air panas Sipaholon, sebelas isolat dari sumber air
panas Tangkuban Perahu, dan dua isolat dari sumber air panas Ciseeng dapat
membentuk daerah bening di sekitar koloninya. Isolat T8 yang berasal dari
Tangkuban Perahu menunjukkan indeks proteolitik paling tinggi, yaitu sebesar
33.5 dan digunakan untuk uji selanjutnya. Aktivitas proteolitik tertinggi bakteri
T8 diperoleh pada jam ke-21 sebesar 3.8375 x 10
-3U/mL. Aktivitas protease
meningkat pada fraksi amonium sulfat 60%, yaitu 7.7257 x 10
-3U/mL. Hasil
analisis statistik menunjukkan bahwa pH dan suhu berpengaruh terhadap aktivitas
protease. Aktivitas optimum diperoleh pada pH 8 bufer Clark & Lubs dan suhu
60
oC.
Thermophilic Bacteria from Tangkuban Perahu Hot Spring Bandung. Supervised
by I MADE ARTIKA and ERNY YUNIARTI.
Protease is an enzyme widely used in food industry. Thermophilic bacteria
produce protease that can withstand high temperature. The research was aimed to
isolate thermophilic bacteria and characterize protease of isolates with highest
proteolytic index.
Isolation was carried out on 33 isolates from three hot springs. Isolation was
done by using pour plate method. Protease activity was measured by Bergmeyer,
and protease characterization was conducted at pH 6-10 and temperature of 50
oC-90
oC. The results showed that two isolates from Sipaholon hot spring, eleven
isolates from Tangkuban Perahu hot spring, and two isolates from Ciseeng hot
spring established clear areas around their colonies. T8 isolate derived from
Tangkuban Perahu hot spring showed the highest proteolytic index of 33.5 and
was used for further testing. The highest proteolytic activity of T8 isolate was
obtained at 21
thhour amounted to 3.8375 x 10
-3U/mL. Protease activity increased
in ammonium sulphate 60% fraction, which was 7.7257 x 10
-3U/mL. Statistical
analysis showed that pH and temperature affected protease activity. The optimum
protease activity was obtained at pH 8 Clark & Lubs buffer and temperature of
60
oC.
ISOLASI DAN OPTIMASI PROTEASE BAKTERI
TERMOFILIK DARI SUMBER AIR PANAS
TANGKUBAN PERAHU BANDUNG
AMELINDA IRENA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2010
NIM
:
G84062714
Disetujui
Komisi Pembimbing
Dr. Ir. I Made Artika, M.App.Sc
Erny Yuniarti, S.Si, M.Si
Ketua
Anggota
Diketahui
Dr. Ir. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan
karunia-Nya sehingga penulis dapat menyelesaikan penelitian dan penulisan karya
ilmiah ini. Karya ilmiah yang berjudul Isolasi dan Optimasi Protease Bakteri
Termofilik dari Sumber Air Panas Tangkuban Perahu Bandung ditulis
berdasarkan hasil penelitian di Laboratorium Biologi Tanah, Balai Penelitian
Tanah selama bulan Maret sampai dengan Agustus 2010.
Penulis mengucapkan terima kasih kepada Dr. Ir. I Made Artika, M.App.Sc
selaku pembimbing utama dan Erny Yuniarti, S.Si, M.Si selaku pembimbing
kedua yang telah memberikan arahan, bimbingan, dan dorongan semangat selama
menyusun karya ilmiah ini. Ucapan terima kasih juga penulis sampaikan kepada
kedua orang tua penulis yang tidak pernah berhenti memberikan doa restu dan
kasih sayangnya, Persica Puspa Chandrakirana dan Ferry Maulana yang telah
memberikan semangat dan dukungan, serta semua staf di Laboratorium Biologi
Tanah, Balai Penelitian Tanah atas semua bantuan dan kerja samanya selama
penelitian ini berlangsung.
Penulis menyadari masih terdapat banyak kekurangan dalam penulisan karya
ilmiah ini. Oleh karena itu, penulis mengharapkan kritik dan saran dari berbagai
pihak. Semoga karya ilmiah ini dapat memberikan manfaat bagi penulis sendiri
maupun semua pihak yang membutuhkannya demi perkembangan ilmu
pengetahuan dan teknologi.
Bogor, November 2010
Didik Purwanto dan Endang Setyorini. Penulis merupakan anak pertama dari dua
bersaudara.
Tahun 2006 penulis lulus dari SMA Negeri 1 Bogor dan pada tahun yang
sama penulis lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB
(USMI) pada Program Studi Biokimia, Fakultas Matematika dan Ilmu
Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum
mata kuliah Struktur Fungsi Subseluler tahun ajaran 2010/2011 dan mata kuliah
Pengantar Penelitian Biokimia tahun ajaran 2010/2011. Penulis juga aktif di
beberapa kepanitiaan kampus dan mengikuti berbagai seminar. Penulis melakukan
praktik lapangan di Balai Penelitian Tanah Bogor selama periode bulan Juni
sampai dengan bulan Agustus 2009 dan menulis laporan ilmiah berjudul Analisis
Kualitatif Kemampuan Melarutkan Fosfat dari Mikrob Pelarut Fosfat.
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
TINJAUAN PUSTAKA
Bakteri Termofilik ... 1
Protease ... 2
Isolasi Bakteri ... 3
Kurva Pertumbuhan Mikrob ... 4
BAHAN DAN METODE
Alat dan Bahan ... 5
Metode Penelitian ... 5
HASIL DAN PEMBAHASAN
Hasil Isolasi Koloni Tunggal Isolat Bakteri Termofilik ... 7
Hasil Uji Aktivitas Protease secara Kualitatif ... 7
Hasil Uji Aktivitas Protease secara Kuantitatif ... 8
Aktivitas Protease Bakteri Termofilik ... 10
Optimasi Protease ... 11
SIMPULAN DAN SARAN
Simpulan ... 13
Saran ... 13
DAFTAR PUSTAKA ... 13
DAFTAR GAMBAR
Halaman
1 Bakteri termofilik Bacillus stearothermophilus ... 2
2 Pola pertumbuhan mikrob ... 5
3 Koloni tunggal bakteri ... 7
4 Hasil uji kualitatif isolat T8 ... 8
5 Grafik indeks proteolitik isolat bakteri ... 8
6 Kurva pertumbuhan bakteri T8 dan produksi protease ... 9
7 Optimasi pH terhadap aktivitas protease ... 13
8 Optimasi suhu terhadap aktivitas protease ... 13
DAFTAR LAMPIRAN
Halaman
1 Bagan alir penelitian ... 17
2 Alur kerja isolasi bakteri termofilik ... 18
3 Pembuatan media ... 19
4 Pembuatan Bufer borat (pH 8), bufer fosfat (pH 6-8) dan bufer Clark
& Lubs (pH 8-10) ... 20
5 Hasil uji kualitatif isolat Ciseeng, Tangkuban Perahu, dan Sipoholon ... 21
6 Alur kerja analisis aktivitas protease ... 22
7 Metode analisis aktivitas protease ... 23
8 Uji kuantitatif aktivitas protease pada ekstrak kasar protease ... 26
9 Uji kuantitatif aktivitas protease dengan pengendapan amonium sulfat ... 27
10 Aktivitas protease pada suhu 50
oC dan berbagai pH ... 28
11 Aktivitas protease pada pH 8 dan berbagai kisaran suhu ... 29
12 Hasil uji statistik menggunakan program SAS untuk pengaruh suhu
terhadap aktivitas protease ... 30
13 Hasil uji statistik menggunakan program SAS untuk pengaruh suhu
terhadap aktivitas protease ... 32
PENDAHULUAN
Indonesia merupakan salah satu negara yang memiliki ekosistem yang beragam. Ekosistem tersebut terdiri atas 42 ekosistem daratan alami dan lima ekosistem lautan. Keragaman ekosistem tersebut membuat Indonesia sangat potensial sebagai tempat untuk mengeksplorasi berbagai jenis organisme, salah satunya adalah mikroorganisme di daerah vulkanik. Kondisi lingkungan daerah vulkanik yang beraneka ragam menyebabkan terdapat beragam mikroorganisme yang dapat tumbuh pada kondisi ekstrim, seperti suhu, pH, dan konsentrasi garam yang tinggi (Madigan & Mars 1997).
Adanya aktivitas vulkanik menyebabkan terbentuknya sumber air panas. Pada sumber air panas, terdapat keragaman mikrob yang sangat menarik untuk dikaji. Mikrob tersebut merupakan mikrob termofilik yang dapat bertahan hidup pada suhu tinggi di sumber air panas. Mikrob termofilik mempunyai kemampuan untuk menggunakan nutrisi yang luas, yaitu dapat hidup secara autotrof maupun heterotrof; aerob dan anaerob; serta fototropik dan kemotropik (Brock 1986).
Keanekaragaman bakteri termofilik memberikan gambaran potensi yang dapat dimanfaatkan untuk berbagai tujuan. Pada saat ini bakteri termofilik dipelajari dan diteliti secara intensif karena alasan pengembangan penelitian dasar dan aplikasi bioteknologi. Bakteri termofilik berpotensi sebagai sumber-sumber enzim khas yang dapat digunakan pada proses pengolahan limbah maupun pelapukan mineral (Brock 1986). Enzim-enzim tersebut mampu bertahan dan aktif pada temperatur yang tinggi. Sifat seperti ini sangat dibutuhkan oleh industri-industri berbasis enzim. Penggunaan enzim yang mampu bertahan pada suhu tinggi dalam bidang bioteknologi dapat menurunkan biaya operasi dan meningkatkan kecepatan reaksi (Aguilar et
al. 1998).
Salah satu enzim yang dapat dihasilkan oleh bakteri termofilik adalah protease. Protease adalah enzim yang berperan dalam reaksi biokatalis yang menyebabkan pemecahan protein. Protease banyak digunakan di bidang industri pangan, seperti keju, bir, roti, industri deterjen, dan industri kulit (Suhartono 1992). Menurut Rao et al. (1998), protease dimanfaatkan untuk proses pengolahan dalam industri pembuatan roti, industri pengolahan kedelai, industri deterjen, dan industri kulit. Protease dapat
dimanfaatkan pada bidang farmasi, tekstil, dan kertas. Protease juga dapat diaplikasikan dalam bidang pertanian. Salah satunya sebagai bioaktivator dalam proses pengomposan. Selama proses pengomposan, bahan kompos mengalami perombakan oleh mikroorganisme, yaitu bakteri atau fungi yang tahan suhu tinggi. Penggunaan protease terutama protease yang diperoleh dari mikrob memiliki beberapa kelebihan, yaitu dapat diproduksi dalam jumlah yang besar, produktifitasnya mudah ditingkatkan, mutu enzim yang dihasilkan seragam, dan harganya murah (Stanbury & Whitaker 1984).
Isolasi dan optimasi mengenai protease yang dihasilkan oleh bakteri termofilik masih terbatas di Indonesia. Padahal Indonesia memiliki banyak sumber air panas. Jika Indonesia yang memiliki banyak sumber air panas dapat mengembangkan penelitian ini, maka banyak keuntungan yang akan diperoleh. Keuntungan tersebut antara lain protease yang diperoleh dari bakteri termofilik dapat digunakan pada suhu tinggi, sehingga sangat menguntungkan dalam bidang industri; protease dapat dihasilkan dalam jumlah yang besar; dan harga protease yang dihasilkan murah.
Penelitian ini bertujuan mengisolasi bakteri termofilik dari sumber air panas Tangkuban Perahu, Ciseeng, dan Sipoholon yang mampu menghasilkan protease termostabil. Selain itu, penelitian juga bertujuan menentukan pH dan suhu optimum protease termostabil yang memiliki indeks proteolitik terbesar. Hipotesis dari penelitian ini adalah isolat bakteri termofilik yang didapatkan dari sumber air panas Tangkuban Perahu, Ciseeng, dan Sipoholon mampu menghasilkan protease termostabil, dan diperoleh kondisi pH dan suhu yang optimum untuk aktivitas protease termostabil. Protease akan memiliki aktivitas tertinggi pada pH dan suhu yang optimum. Isolat bakteri termofilik penghasil protease yang diperoleh pada penelitian ini diharapkan memiliki potensi yang dapat diaplikasikan pada proses industri yang melibatkan protease pada suhu tinggi. Selain itu, protease yang diperoleh diharapkan memiliki aktivitas yang tinggi pada pH dan suhu optimum.
TINJAUAN PUSTAKA
Bakteri Termofilik
Salah satu jenis mikroorganisme yang banyak dieksplorasi adalah mikroorganisme termofilik yang hidup di daerah sekitar
gunung berapi dan sumber air panas. Mikroorganisme yang dapat hidup pada suhu termofilik hanyalah prokariot dari kelompok Arkaea dan Bakteri (Madigan & Marrs 1997). Suhu merupakan salah satu faktor lingkungan yang mempengaruhi pertumbuhan dan kemampuan bertahan hidup mikroorganisme. Mikroorganisme memiliki suhu minimum dan maksimum yang menjadi batas pertumbuhannya, serta suhu optimum yang menunjukkan pertumbuhan tercepat. Suhu optimum tersebut biasanya mendekati suhu maksimum (Brock 1986). Berdasarkan suhu optimum pertumbuhannya, secara umum mikroorganisme dibedakan menjadi psikofilik (-3-20ºC), mesofilik (20-45ºC), termofilik (45-65ºC), ekstrim termofilik (65-85ºC), dan hipertermofilik (85-110ºC) (Edwards 1990).
Bakteri termofilik merupakan kelompok bakteri yang dapat beradaptasi dengan kondisi lingkungan yang bersuhu tinggi. Bakteri termofilik dapat tumbuh pada suhu yang relatif tinggi dengan suhu minimum 25°C, suhu optimum 45-55°C, dan suhu maksimum 55-65°C. Bakteri termofilik dapat beradaptasi dengan pH ekstrim (<2.0, >10.0). Habitat alami bakteri termofilik tersebar luas di seluruh permukaan bumi. Salah satu lingkungan alaminya terbentuk akibat aktivitas vulkanik atau perpindahan kerak bumi pada saat gempa tektonik. Fenomena geologi tersebut menghasilkan kawah air panas.
Bakteri termofilik pertama kali diisolasi pada tahun 1879 oleh Miquel, yang mampu
berkembang biak pada suhu 72oC. Bakteri
termofilik juga berhasil diisolasi dari kawah air panas dan sedimen lautan geotermal (Edwards 1990). Spesies termofilik umumnya banyak ditemukan pada lingkungan yang ekstrim, misalnya daerah vulkanik dan sumber air panas. Bakteri ini umumnya berasal dari genus Beggiatoa,
Thermochromatium, Acidithiobacillus, Thermithiobacillus, Thermomonas, Methylococcus, Pyrococcus dan Alterococcus
(Labeda 1990).
Menurut Brock (1986), terdapat tiga faktor yang menyebabkan bakteri termofilik mampu bertahan hidup dan berkembang biak pada suhu tinggi, yaitu kandungan enzim dan protein yang lebih stabil dan tahan terhadap panas, molekul pensintesis protein yang stabil terhadap panas, dan membran lipid sel termofil mengandung banyak asam lemak jenuh yang membentuk ikatan hidrofobik yang sangat kuat. Bakteri termofilik, contohnya Thermus aquaticus memiliki
membran termostabil yang akan memproduksi lemak yang memiliki titik cair yang lebih tinggi ketika temperatur lingkungan naik. Bakteri termofilik mampu mensintesis molekul stabil, seperti enzim yang mampu mengkatalisis reaksi-reaksi biokimia pada suhu tinggi dan lebih stabil dibandingkan enzim yang dihasilkan bakteri mesofilik.
Keanekaragaman bakteri termofilik memberikan gambaran potensi yang dapat dimanfaatkan untuk berbagai tujuan. Pada saat ini bakteri termofilik dipelajari dan diteliti secara intensif karena alasan pengembangan penelitian dasar dan aplikasi bioteknologi. Bakteri termofilik berpotensi sebagai sumber-sumber enzim khas yang dapat digunakan pada proses pengolahan limbah maupun pelapukan mineral (Brock 1986). Enzim-enzim tersebut mampu bertahan dan aktif pada temperatur yang tinggi. Sifat seperti ini sangat dibutuhkan oleh industri-industri berbasis enzim. Penggunaan enzim yang mampu bertahan pada suhu tinggi dalam bidang bioteknologi dapat menurunkan biaya operasi dan meningkatkan kecepatan reaksi (Aguilar et
al. 1998).
Gambar 1 Bakteri termofilik Bacillus stearothermophilus.
Protease
Enzim merupakan unit fungsional dari metabolisme sel. Enzim bekerja dengan urut-urutan yang teratur. Enzim mengkatalisis ratusan reaksi bertahap yang menguraikan molekul nutrien. Enzim juga mengkatalisis reaksi penyimpan energi kimiawi dan membuat makromolekul sel dari prekursor sederhana (Lehninger 1982). Organisme hidup yang berperan sebagai sumber enzim meliputi tanaman, hewan, dan mikrob. Enzim yang banyak dimanfaatkan oleh manusia adalah enzim yang berasal dari mikrob. Hal tersebut dikarenakan enzim yang berasal dari mikrob memiliki produktivitas tinggi, pertumbuhannya mudah, sifat enzim yang dapat diubah ke arah yang menguntungkan, dan berkembangnya teknik fermentasi,
mutasi, dan rekayasa genetik. Salah satu enzim yang dihasilkan oleh mikrob adalah protease. Protease merupakan salah satu enzim yang resisten terhadap panas. Berbagai jenis bakteri seperti Bacillus, Pseudomonas,
Clostridium, Proteus, dan Seratia merupakan
penghasil enzim protease yang cukup potensial (Suhartono 1989). Protease juga dihasilkan oleh sejenis kapang, seperti
Aspergillus oryzae, Penicillium, Aspergillus niger, dan Mucor miehei (Ramadzanti 2006).
Protease adalah enzim yang berperan dalam reaksi biokatalis yang menyebabkan pemecahan protein. Protease menghidrolisis protein menjadi polipeptida dan asam amino. Menurut Komisi Tatanama Internasional
Union of Biochemist and Molecular Biologist, protease termasuk ke dalam
kelompok hidrolase pemecah protein (kelompok ke III subkelompok IV). Kerja enzim ini membutuhkan air, sehingga dimasukkan dalam kelas hidrolase (Ward 1983). Protease sering disebut enzim proteolitik karena dapat merusak protein (Janzen et al. 1982). Protease terdiri atas proteinase dan peptidase. Proteinase mengkatalisis reaksi hidrolisis molekul protein menjadi fragmen besar polipeptida. Peptidase menghidrolisis fragmen besar polipeptida menjadi asam amino (Winarno 1985).
Protease dapat dihasilkan secara intraseluler dan ekstraseluler oleh hewan, tanaman, dan mikrob yang berperan penting dalam metabolisme dan regulasi sel. Protease ekstraseluler diperlukan makhluk hidup untuk menghidrolisis nutrisi protein menjadi peptida kecil dan asam amino, sehingga dapat diserap dan dimanfaatkan oleh sel tubuhnya. Protease intraseluler bertanggung jawab terhadap degradasi proteolitik secara cepat dan irreversible bagi protein sel yang fungsinya tidak diperlukan lagi atau protein abnormal yang tidak bermanfaat bahkan mengganggu metabolisme sel (Suhartono 1992).
Secara garis besar protease dibagi menjadi dua kelompok, yaitu endopeptidase dan eksopeptidase. Endopeptidase akan memotong ikatan peptida yang berada pada bagian tengah rantai polipeptida, sedangkan eksopeptidase memotong di ujung rantai polipeptida baik berupa ujung karboksil maupun ujung asam amino. Penggolongan protease lainnya adalah berdasarkan data deret asam amino enzim yang mengarah kepada hubungan evolusi dan struktur enzim. Klasifikasi ini sangat penting, mengingat kemiripan struktur enzim di dalam keluarga
yang sama, biasanya mencerminkan kemiripan dalam hal mekanisme katalitik. Menurut Rao et al. (1998), berdasarkan pH kerjanya protease dapat dikelompokkan menjadi tiga bagian, yaitu protease asam, netral, dan alkali. Kelompok protease asam terdiri atas protease aspartat dan beberapa protease sistein yang memiliki pH optimum antara dua sampai enam. Protease netral aktif pada kisaran pH netral. Protease alkali ditemukan aktif pada pH antara delapan dan tiga belas.
Ward (1983) mengklasifikasikan protease berdasarkan sifat-sifat kimia sisi aktif enzim, yaitu protease serin (memiliki asam amino serin pada sisi aktifnya), protease sulfhidril (memiliki gugus sulfhidril pada sisi aktifnya), protease metal (memiliki ion logam pada sisi aktifnya), dan protease asam (memiliki dua gugus karboksil pada sisi aktifnya). Menurut Sadikin (2002), berdasarkan mekanisme katalitiknya protease dapat dibagi menjadi empat, yaitu protease serin, protease sistein, protease aspartat, dan protease logam.
Protease banyak digunakan di bidang industri pangan, seperti keju, bir, roti, industri deterjen, dan industri kulit (Suhartono 1992). Menurut Rao et al. (1998), protease dimanfaatkan untuk proses pengolahan dalam industri pembuatan roti, industri deterjen, dan industri kulit. Protease juga dapat dimanfaatkan pada bidang farmasi, industri tekstil, dan kertas. Penggunaan protease terutama protease yang diperoleh dari mikrob memiliki beberapa kelebihan, yaitu dapat diproduksi dalam jumlah yang besar, produktifitasnya mudah ditingkatkan, mutu enzim yang dihasilkan seragam, dan harganya murah (Stanbury & Whitaker 1984).
Isolasi Bakteri
Proses isolasi bakteri termofilik pada dasarnya tidak berbeda jauh dengan isolasi bakteri mesofilik, tetapi isolasi bakteri termofilik memerlukan suhu tinggi. Salah satu kendala yang dihadapi ketika mengisolasi bakteri, terutama bakteri termofilik adalah kesulitan untuk mengisolasi bakteri yang memiliki kecepatan pertumbuhan yang rendah. Pada umumnya bakteri yang berhasil diisolasi adalah bakteri yang memiliki kecepatan pertumbuhan yang tinggi, sehingga menjadi dominan di lingkungannya. Populasi bakteri termofilik mungkin sangat sedikit di habitat lain, sehingga media untuk proses isolasi harus diperkaya (Sanfitri 2007).
Hal-hal yang harus diperhatikan sebelum mengisolasi bakteri adalah pemilihan substrat pada media yang digunakan. Media yang digunakan untuk isolasi dapat berupa media diferensial, media selektif, dan media penyubur. Hal lain yang harus diperhatikan adalah tempat bakteri ditemukan. Sebagai contoh, untuk mengisolasi bakteri yang dapat mendegradasi senyawa hidrokarbon polisiklik aromatik akan lebih mudah untuk mendapatkannya pada daerah yang terkontaminasi oleh senyawa hidrokarbon tersebut yang kemudian diperkaya oleh senyawa lain sebagai sumber karbon atau sumber energi (Sanfitri 2007). Faktor-faktor lain yang harus diperhatikan dalam mengisolasi bakteri adalah keasaman dan suhu lingkungan.
Proses isolasi bakteri termofilik sama dengan teknik isolasi pada umumnya. Beberapa tahapan yang dapat dilakukan untuk mengisolasi dan seleksi bakteri termofilik adalah pemilihan substrat yang sesuai dengan bakteri yang akan diisolasi. Sebagian sampel yang diperoleh dapat ditumbuhkan langsung dalam medium agar (dicawankan) atau disuspensikan dan diencerkan dalam media cair sebelum dicawankan. Tahapan selanjutnya adalah penyuburan dengan cara menambahkan nutrisi tertentu agar meningkatkan populasi bakteri yang ingin diisolasi. Setelah tahap penyuburan, dilakukan pemurnian dan isolasi bakteri yang diinginkan. Bakteri yang telah diisolasi, dapat diseleksi. Seleksi bertujuan menguji kemampuan isolat menghasilkan produk yang diharapkan. Seleksi isolat biasanya menggunakan media spesifik dengan substrat tertentu (Labeda 1990).
Kurva Pertumbuhan Mikrob
Pertumbuhan adalah penambahan secara teratur semua komponen sel suatu jasad. Pertumbuhan juga dapat diartikan sebagai penambahan jumlah sel, misalnya pertumbuhan yang terjadi pada kultur suatu mikrob. Pada organisme bersel tunggal (uniseluluer) pertumbuhan merupakan penambahan jumlah individu. Pembelahan sel pada organisme bersel banyak (multiseluler) tidak menghasilkan penambahan jumlah individunya, tetapi terbentuknya jaringan atau peningkatan ukuran suatu jasad (Sokatch 1973).
Bakteri yang dimasukkan ke dalam medium yang sesuai akan tumbuh memperbanyak diri. Penambahan jumlah sel bakteri dilakukan melalui penambahan sel secara biner. Pembelahan biner adalah
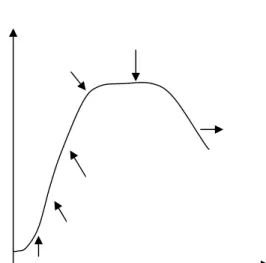
pembelahan langsung tanpa adanya tahapan mitosis. Proses pembelahan biner diawali dengan proses replikasi DNA, kemudian diikuti dengan pembagian sitoplasma. Proses pembelahan biner berlangsung cepat. Setiap 20 menit, satu sel bakteri Escherichia coli akan membelah menjadi dua sel yang sama. Secara matematika, pertumbuhan tersebut termasuk ke dalam pertumbuhan eksponensial. Jika pada waktu-waktu tertentu jumlah bakteri dihitung dan dibuat grafik hubungan antara jumlah bakteri dan waktu, maka akan diperoleh suatu grafik atau kurva pertumbuhan (Gambar 2). Pola pertumbuhan pada Gambar 2 menunjukkan pertumbuhan mikrob yang terdiri atas beberapa fase, yaitu fase permulaan (adaptasi), fase pertumbuhan awal, fase pertumbuhan logaritma (eksponensial), fase pertumbuhan akhir, fase stasioner maksimum, fase kematian dipercepat, dan fase kematian logaritma (Sokatch 1973).
Fase pertumbuhan dimulai pada fase permulaan (adaptasi). Pada fase permulaan, bakteri baru menyesuaikan diri dengan lingkungannya yang baru, sehingga bakteri belum membelah diri. Fase permulaan dipengaruhi oleh medium dan jumlah inokulum. Bakteri mulai membelah diri pada fase pertumbuhan yang dipercepat. Fase permulaan hingga fase pertumbuhan dipercepat disebut fase lag. Pada fase lag, sel akan mengalami penambahan ukuran, peningkatan metabolisme, peningkatan struktur DNA, dan peningkatan protein. Kecepatan sel membelah diri paling cepat terjadi pada fase pertumbuhan logaritma (eksponensial). Selama fase logaritma, metabolisme sel paling aktif, sintesis bahan sel sangat cepat dengan jumlah konstan hingga nutrien habis. Fase log sangat dipengaruhi oleh komposisi media pertumbuhan. Setelah fase logaritma, pertumbuhan bakteri mulai terhambat, kecepatan pembelahan sel berkurang, dan jumlah sel yang mati mulai bertambah. Pada fase stasioner maksimum, jumlah sel yang mati akan sebanding dengan jumlah sel yang hidup, sehingga jumlah sel konstan. Kecepatan kematian sel akan terus meningkat pada fase kematian yang dipercepat. Populasi bakteri akan mengalami fase kematian karena nutrien sudah tidak tersedia dan terjadi penimbunan hasil metabolisme yang bersifat toksik. Kecepatan kematian sel mencapai maksimal sampai pada fase kematian logaritma, sehingga jumlah sel hidup menurun dengan cepat seperti deret ukur. Penurunan jumlah sel hidup tidak mencapai
nol, karena dalam jumlah minimum tertentu sel bakteri akan tetap bertahan sangat lama dalam medium (Sokatch 1973).
Gambar 2 Pola pertumbuhan mikrob.
BAHAN DAN METODE
Alat dan Bahan
Alat-alat yang digunakan adalah termos panas, mikropipet, tip, tabung Eppendorf, autoklaf, laminar air flow cabinet, cawan Petri, waterbath shaker, oven, sentrifus, jarum ose, spektrofotometer, tabung reaksi, vortex, termometer, pH meter digital, inkubator, neraca analitik, magnetic stirrer, pipet Mohr, lemari pendingin, plastik tahan panas, kapas, karet, tisu, aluminium foil, dan palstik wrap. Pada penelitian ini juga digunakan berbagai macam peralatan gelas, seperti labu Erlenmeyer dan gelas ukur.
Bahan-bahan yang digunakan adalah sampel air panas yang diambil dari sumber air panas Tangkuban Perahu, Ciseeng, dan Sipoholon. Bahan lain yang digunakan adalah alkohol 70%, akuades, NaCl, bakto pepton, glukosa, bakto agar, susu skim cair, ekstrak ragi, amonium sulfat, spirtus, bufer borat 0.1 M, kasein 20 mg/mL, HCl 0.05 M,
standar tirosin 5 mM, TCA 0.1 M, CaCl2 0.2
mM, Na2CO3 0.4 M, pereaksi Folin
Ciocalteau, bufer fosfat (pH 6.0-8.0), dan bufer Clark & Lubs (pH 8.0-10.0).
Metode Penelitian
Pelaksanaan penelitian meliputi pengambilan sampel, isolasi bakteri termofilik, pembuatan media susu skim agar, seleksi bakteri proteolitik termofilik, pembuatan stok kultur proteolitik termofilik, pembuatan media produksi, pembuatan kurva pertumbuhan dan kurva produksi protease, Isolasi protease, pengendapan ekstrak kasar
dengan amonium sulfat, penentuan aktivitas protease, optimasi aktivitas protease, dan analisis statistika.
Pengambilan Sampel
Sampel yang digunakan berupa air panas yang berasal dari sumber air panas di Ciseeng, Tangkuban Perahu, dan Sipoholon. Sampel air panas dibawa ke laboratorium dalam termos panas.
Isolasi Bakteri Termofilik (Cappucino & Sherman 1983)
Sampel diencerkan dengan garam fisiologis steril (0.85%). Sebanyak 5 mL sampel dimasukkan ke dalam labu Erlenmeyer yang berisi 45 mL garam fisiologis steril, kemudian dibuat deret
pengenceran 10-2, 10-3, 10-4, 10-5, dan 10-6.
Sebanyak 1 mL dari tiap pengenceran dicawankan pada media PGA (Pepton Glukosa Agar) (duplo) menggunakan metode cawan tuang. Media yang telah diinokulasi, diinkubasi pada suhu 50°C selama 2 hari. Koloni tunggal ditumbuhkan pada media agar miring pepton glukosa dan diinkubasi pada suhu 50°C selama 2 hari. Setelah inkubasi, media agar miring pepton glukosa disimpan pada suhu ruang.
Pembuatan Media Susu Skim Agar
Sebanyak 6 gram bakto agar dimasukkan ke dalam Erlenmeyer yang berisi 200 mL akuades, kemudian disterilisasi 15 menit pada suhu 121°C. Setelah itu 100 mL susu skim yang telah steril dicampurkan ke dalam media agar dalam keadaan hangat (50°C) lalu dihomogenkan. Segera setelah itu media dituang ke dalam cawan Petri steril.
Seleksi Bakteri Proteolitik Termofilik (Oktafianti 2005)
Isolat bakteri hasil isolasi diseleksi kemampuan aktivitas proteolitiknya secara kualitatif pada media susu skim agar. Isolat bakteri termofilik dari media agar miring ditotolkan menggunakan lup inokulasi dengan ujung runcing di pusat media agar cawan Petri. Media diinkubasi pada suhu 50°C selama 2 hari. Adanya aktivitas protease secara kualitatif ditunjukkan dengan terbentuknya daerah bening di sekitar koloni. Aktivitas proteolitik semikuantitatif adalah rasio diameter daerah bening dan diameter koloni. Pengukuran diameter koloni dan diameter daerah bening dilakukan menggunakan kertas milimeter blok. Koloni bakteri yang mempunyai nilai rasio tinggi merupakan isolat bakteri yang memiliki Fase adaptasi
Fase pertumbuhan awal Fase logaritmik Fase stasioner Fase pertumbuhan akhir Fase kematian
5
aktivitas proteolitik dan mempunyai peluang untuk dapat dikembangkan atau dimanfaatkan lebih lanjut.
Pembuatan Stok Kultur Proteolitik Termofilik
Media yang digunakan adalah PGB (Peptone Glucose Broth). Media PGB dibuat sebanyak 100 mL dalam labu Erlenmenyer 250 mL dengan komposisi 5 g/L glukosa dan 10 g/L pepton. Sebanyak satu ose isolat dari biakan kerja pada agar miring pepton glukosa diinokulasikan ke dalam media PGB, kemudian biakan diinkubasi bergoyang dalam pemanas air dengan kecepatan 120
rpm pada suhu 50oC selama 20 jam.
Pembuatan Media Produksi
Media produksi yang digunakan adalah PGY (Peptone Glucose Yeast Extract). Media produksi dibuat sebanyak 300 mL dalam Labu Erlenmeyer 500 mL dengan komposisi 5 g/L glukosa, 10 g/L pepton, dan 10 g/L ekstrak ragi, kemudian disterilisasi menggunakan autoklaf selama 15 menit pada
suhu 121oC. Setelah itu media produksi
ditambahkan 1% (v/v) susu skim cair yang telah steril.
Pembuatan Kurva Pertumbuhan dan Kurva Produksi Protease
Sebanyak 1% (v/v) inokulum segar dari media PGB diinokulasikan ke dalam media PGY, kemudian diinkubasi bergoyang
dengan kecepatan 120 rpm dan suhu 50oC.
Densitas sel diukur setiap 2 jam selama 30 jam pada panjang gelombang 600 nm hingga diperoleh kurva pertumbuhan. Kurva produksi diperoleh dengan cara pengukuran aktivitas proteolitik secara kuantitatif pada supernatan sampel yang diambil setiap tiga jam. Berdasarkan percobaan tersebut, diperoleh korelasi antara kurva pertumbuhan dan kurva produksi yang menunjukkan waktu optimum untuk panen protease.
Isolasi Protease (Sutandi 2003)
Isolasi enzim dilakukan dengan menggunakan sentrifus pada kecepatan 10000 rpm selama 5 menit. Isolasi protease dilakukan pada waktu optimum aktivitas protease. Filtrat yang diperoleh merupakan ekstrak kasar protease.
Pengendapan Ekstrak Kasar Protease dengan Amonium Sulfat (Susanti 2003)
Ekstrak kasar protease diendapkan dengan amonium sulfat konsentrasi kejenuhan 20%, 40%, 60%, dan 80%.
Penambahan amonium sulfat dilakukan sedikit demi sedikit sambil diaduk pada suhu
5oC hingga larutan jenuh. Ekstrak kasar yang
telah diendapkan, kemudian disentrifugasi pada kecepatan 6000 rpm selama 30 menit. Endapan yang diperoleh dilarutkan dengan bufer fosfat 50 mM pH 8 dan disimpan di dalam lemari pendingin.
Penentuan Aktivitas Protease (Bergmeyer 1983)
Sebanyak tiga buah tabung reaksi bersih diberi label blanko, sampel, dan standar, kemudian sebanyak 1 mL bufer borat (0.1 M), 1 mL kasein (20 mg/mL), dan 0.2 mL HCl (0.05 M) dimasukkan masing-masing ke dalam tiga tabung reaksi yang berbeda. Tabung blanko, sampel, dan standar, masing-masing ditambahkan 0.2 mL akuades, 0.2 mL ekstrak kasar enzim dalam
CaCl2 (0.2 mM), dan 0.2 mL standar tirosin
(5 mM), kemudian ketiga tabung diinkubasi
selama 10 menit pada suhu 50oC. Setelah itu
ke dalam masing-masing tabung ditambahkan
2 mL TCA (0.1 M), dan 0.2 mL CaCl2
(0.2 mM) ke dalam tabung sampel dan
0.2 mL ekstrak kasar enzim dalam CaCl2
(0.2 mM) ke dalam tabung blanko dan standar, lalu ketiga tabung reaksi tersebut
diinkubasi kembali pada suhu 50oC selama
10 menit. Kemudian masing-masing larutan disentrifus selama 10 menit dengan kecepatan 4000 rpm. Sebanyak 1.5 mL filtrat dari masing-masing larutan dipindahkan ke dalam tiga tabung reaksi baru yang berbeda, lalu ke dalam masing-masing tabung ditambahkan
5 mL Na2CO3 (0.4 M) dan 1 mL pereaksi
Folin. Setelah itu masing-masing larutan tersebut diukur menggunakan spektrofotometer pada panjang gelombang 578 nm.
Unit aktivitas enzim dihitung berdasarkan persamaan:
UA = x P x x x [standar]
Keterangan :
UA : Unit aktivitas enzim Asp : Nilai absorbansi sampel Abl : Nilai absorbansi blanko Ast : Nilai absorbansi standar
P : Faktor pengenceran
T : Waktu inkubasi
Optimasi Aktivitas Protease (Vratyastoma 2006)
Optimasi pH dan suhu dilakukan untuk memperoleh aktivitas protease tertinggi. Pengukuran pH optimum dilakukan dengan
cara mengukur aktivitas protease pada kisaran pH 6, pH 7, pH 8, pH 9, dan pH 10 menggunakan metode Bergmeyer (1983). Larutan bufer yang digunakan adalah bufer fosfat (pH 6-7), bufer borat (pH 8), dan bufer Clark & Lubs (pH 8-10). Penentuan suhu optimum dilakukan dengan cara mengukur
aktivitas protease pada kisaran suhu 50-90oC
dengan selang suhu 10oC pada pH optimum.
Analisis Data Statistik
Rancangan penelitian yang digunakan dalam penelitian adalah Rancangan Acak Lengkap (RAL), dengan model liner aditif yang digunakan adalah model tetap. Model tetap merupakan suatu model dengan perlakuan-perlakuan yang digunakan berasal dari populasi-populasi yang terbatas dan pemilihan perlakuan ditentukan oleh peneliti (Matjik & Sumertajaya 2000). Uji lanjut yang digunakan adalah uji pembanding berganda Duncan.
Model linier yang digunakan pada rancangan acak lengkap adalah sebagai berikut.
Yij = µ + τi + εij Keterangan :
Yij = aktivitas protease pada perlakuan ke-i dan ulangan ke-j
µ = rataan umum
τi = pengaruh perlakuan ke-i
εij = pengaruh acak pada perlakuan ke-i dan ulangan ke-j
i = pH 6, pH 7, pH 8, pH 9, pH 10 j = 1, 2, 3
Hipotesis :
H0 : tidak ada pengaruh pH terhadap aktivitas protease.
H1 : paling sedikit ada satu pengaruh pH terhadap aktivitas protease.
Yij = µ + τi + εij Keterangan :
Yij = aktivitas protease pada perlakuan ke-i dan ulangan ke-j
µ = rataan umum
τi = pengaruh perlakuan ke-i
εij = pengaruh acak pada perlakuan ke-i dan ulangan ke-j
i = suhu 50oC, 60oC, 70oC, 80oC, dan 90oC
j = 1, 2, 3 Hipotesis :
H0 : tidak ada pengaruh suhu terhadap aktivitas protease.
H1 : paling sedikit ada satu pengaruh suhu terhadap aktivitas protease.
HASIL DAN PEMBAHASAN
Hasil Isolasi Koloni Tunggal Isolat Bakteri Termofilik
Bakteri termofilik yang digunakan pada penelitian ini merupakan bakteri yang diisolasi dari sumber air panas Ciseeng, Tangkuban Perahu, dan Sipoholon. Pada ketiga sumber air panas tersebut diduga terdapat bakteri termofilik yang memiliki aktivitas proteolitik. Proses isolasi bakteri termofilik pada dasarnya tidak berbeda jauh dengan isolasi bakteri mesofilik, tetapi isolasi bakteri termofilik memerlukan suhu tinggi. Suhu inkubasi yang digunakan pada tahap
isolasi adalah 50oC. Tahapan isolasi
bertujuan memperoleh bakteri dalam koloni tunggal dari campuran populasi bakteri di sumber air panas Tangkuban Perahu, Ciseeng, dan Sipoholon.
Isolasi koloni tunggal bakteri dilakukan dengan media PGA (Pepton Glukosa Agar). Isolasi dilakukan dalam media padat dengan teknik cawan tuang. Prinsip dari teknik isolasi cawan tuang adalah mengencerkan organisme, sehingga individu spesies dapat dipisahkan satu dengan lainnya. Gambar 3 menunjukkan koloni tunggal yang terbentuk pada tahap isolasi. Berdasarkan hasil isolasi yang dilakukan pada media PGA, diperoleh sepuluh isolat dari air panas Sipoholon, 15 isolat dari sumber air panas Tangkuban Perahu, dan delapan isolat dari sumber air panas Ciseeng.
Gambar 3 Koloni tunggal bakteri.
Hasil Uji Aktivitas Protease secara Kualitatif
Isolat-isolat bakteri yang diperoleh pada tahap isolasi, diuji secara kualitatif pada media seleksi. Uji kualitatif bertujuan mengetahui bakteri yang memiliki aktivitas proteolitik. Media seleksi yang digunakan dalam penelitian ini adalah susu skim. Menurut Widhyastuti & Naiola (2002), media yang digunakan untuk seleksi bakteri proteolitik mengandung susu skim dan agar. Susu merupakan media yang sesuai untuk pertumbuhan mikrob karena kaya akan
koloni tunggal
n y a p s t d d p ( k d k k m b d ( p a p b m m L b d b a i p ( u u p m d p s s d d m k p T d m s m a d p l d k p nutrien. Susu yang tertinggal atau seluruhny pada lemak da sedangkan se terkandung pa dengan yang dipisahkan. Su protein, 1% (Hartoto 2003) Pengukura kualitatif dila daerah bening kualitatif m kemampuan merombak pro besarnya daer dengan be (Widhyastuti protein susu amino menyeb putih menjadi bakteri yang media seleksi membentuk da Luasnya daera bakteri tidak m dihasilkan oleh bening yang akan bertamba inkubasi. Oleh pada bakteri (Sutandi 2003) uji kualitatif untuk meng proteolitik sua menunjukkan di sekitar ko proteolitik. Berdasark secara kualita sumber air pa dari sumber air dua isolat dari mampu memb koloninya. Ga proteolitik isol T8 yang ber dengan indek merupakan iso selanjutnya. menjadi dasa aktivitas enzi dilakukan uji proteasenya licheniformis daerah bening kemampuan ya protease (Sian skim adalah b l setelah krim ya. Perbedaan an vitamin-vita emua kompo ada susu skim
ada pada su usu skim meng
lemak, dan ). an aktivitas p akukan deng g. Aktivitas h merupakan g isolat bakt otein dengan rah bening d sarnya dia & Naiola 2 oleh protease babkan peruba i tidak berwa diuji secara memiliki prot aerah bening d ah bening di mewakili jumla h suatu bakter dihasilkan ole ah dengan bert h karena itu kan
perlu diuji s ). Menurut Sal merupakan u etahui perbe atu isolat bak daerah bening loni yang m kan uji aktiv atif, diperoleh anas Sipoholon r panas Tangku sumber air pan bentuk daerah b ambar 5 menu lat yang diuji rasal dari Tan ks proteolitik olat yang digu Uji kualitati ar yang baik im protease, i lanjutan ter (Ward 19 hanya mamp yang kecil, na ang besar dala n 1992). Oleh
bagian dari su diambil sebagi susu skim han amin larut lem nen gizi ya m tidak berbe usu yang belu gandung 37.40 49.20% lakto proteolitik seca gan pengamat hidrolisis seca gambaran d teri proteoli membandingk i sekitar kolo ameter kolo 002). Degrad e menjadi asa ahan warna d arna. Jika iso kualitatif pa tease, maka ak di sekitar kolo sekeliling kolo ah protease ya ri, karena daer eh suatu bakt tambahnya wak ndungan protea ecara kuantita lem et al. (200 uji pendahulu edaan aktivi kteri. Gambar g yang terbent emiliki aktivi vitas proteoli dua isolat d n, sebelas iso uban Perahu, d nas Ciseeng ya bening di seki unjukkan inde kualitatif. Iso ngkuban Pera k sebesar 33 unakan untuk f tidak sela untuk melih sehingga pe rhadap aktivi 983). Bacil pu menghasilk amun mempuny am menghasilk
karena itu sela usu ian nya ak, ang eda um 0% osa ara tan ara dari itik kan oni oni dasi am dari olat ada kan oni. oni ang rah teri ktu ase atif 09), uan itas r 4 tuk itas itik dari olat dan ang itar eks olat ahu 3.5 uji alu hat erlu itas llus kan yai kan ain uji peng kuan perb kual diseb alam di l tidak terda beni deng Bebe peny hidro aktiv kece med yang G Gam sang sepe kom 0 5 10 15 20 25 30 35 40 Inde k s P rot eo lit ik aktivitas pr gukuran akt ntitatif perlu bedaan nilai itatif dan babkan oleh mi pertumbuhan aboratorium, k optimum (Sta Menurut Wa apat korelasi ng di sekitar gan kemampu erapa faktor yebab tidak te olisis secara vitas enzim epatan pertum dium padat atau g diberikan pad Gambar 4 Hasi mbar 5 Grafik bakteri. Hasil Uji A secar Pertumbuhan gat dipengaruh erti fisika-kim mposisi media 3.75 13.25 11.36 15 17 0 5 0 5 0 5 0 5 0 T1 T2 T3 T4 T rotease secara tivitas prote u dilakukan aktivitas prot kuantitatif perbedaan k n bakteri deng sehingga aktiv anbury & Whit ard (1983) yang baik an koloni pada uan organism yang didu rkorelasinya n kualitatif d secara kuanti mbuhan setiap
u cair dan juml da kedua mediu il uji kualitatif indeks proteo Aktivitas Prot ra Kuantitatif suatu mik hi oleh faktor mia (suhu da a tumbuh (S 7.5 3.56 33.5 3.75 3.2 3.12 T6 T7 T8 T9 T11 T14 Kode Isolat a kualitatif, ase secara n. Adanya tease secara antara lain kondisi suhu gan perlakuan vitas bakteri taker 1984). tidak selalu ntara daerah media padat me tersebut. uga menjadi nilai aktivitas dengan nilai itatif adalah isolat pada lah inokulum um. isolat T8. olitik isolat tease f kroorganisme lingkungan, an pH) dan Stanbury & 2 5 6.75 4 2 5.2 T15 C5 C3 S3 S7
Whitaker 1984). Media produksi untuk menghasilkan enzim harus memenuhi kebutuhan dasar untuk menghasilkan sel serta produk. Unsur utama yang paling dibutuhkan adalah nitrogen dan karbon. Nitrogen diperlukan untuk pertumbuhan sel, sedangkan karbon digunakan untuk meningkatkan energi biosintesis. Media produksi yang digunakan pada penelitian ini adalah PGY (Peptone
Glucose Yeast Extract) yang ditambahkan 1%
(v/v) susu skim cair. Media PGY terdiri atas pepton, glukosa, dan ekstrak ragi. Menurut Purbowo (1988) aktivitas protease yang dihasilkan pada media sintetik lebih besar dibandingkan aktivitas protease yang dihasilkan pada media nonsintetik. Sebagai
starter digunakan biakan bakteri T8 yang
sudah ditumbuhkan dalam medium PGB (Peptone Glucose Broth). Sebanyak 1% inokulum dari stok kultur proteolitik termofilik dimasukkan ke dalam media produksi. Konsentrasi inokulum yang diinokulasikan berpengaruh terhadap produksi protease. Produksi protease maksimal pada konsentrasi inokulum 2%. Penambahan inokulum di atas konsentrasi 2%, akan menurunkan produksi protease (Sharmin et al. 2005).
Menurut Fardiaz (1988) dalam memproduksi enzim dalam suatu bioproses memerlukan beberapa faktor, antara lain jenis mikrob, kurva pertumbuhan, dan kondisi optimum untuk meningkatkan aktivitas enzim. Oleh karena itu perlu dilakukan pengamatan pertumbuhan bakteri T8. Kemampuan bakteri memperbanyak sel dalam medium ditunjukkan oleh kekeruhan yang terbentuk pada medium. Kekeruhan terjadi karena sel bakteri tumbuh, memperbanyak diri, dan mensekresikan enzim ke medium (Sutandi 2003).
Pembuatan kurva pertumbuhan dilakukan untuk menentukan waktu inkubasi pertumbuhan optimum bakteri T8. Sintesis protease akan mengalami penurunan setelah waktu optimum tercapai. Hal tersebut antara lain disebabkan substrat enzim telah berkurang, selain itu tingginya kandungan asam amino dalam media dapat berperan sebagai represor sintesis enzim atau terjadinya penguraian oleh enzim itu sendiri karena tidak adanya lagi protein yang dapat digunakan sebagai substrat (Stanburry & Whitaker 1984).
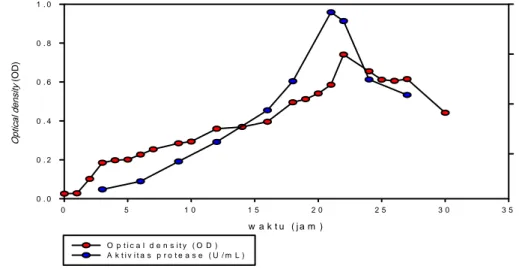
Pengamatan pola pertumbuhan bakteri proteolitik dilakukan berdasarkan metode turbidimetrik. Bakteri yang tumbuh akan menghasilkan pertambahan jumlah sel, sehingga dapat diukur berdasarkan kepekatan sel dalam media. Pengamatan pola pertumbuhan dilakukan selama 1 x 30 jam. Pengukuran angka rapat optis dilakukan setiap dua jam sekali pada panjang gelombang 600 nm. Waktu pengukuran setiap dua jam dipilih sesuai dengan pertumbuhan bakteri yang bertambah jumlah selnya setiap dua jam. Berdasarkan kurva pertumbuhan Gambar 6, bakteri proteolitik isolat T8 memiliki pola pertumbuhan yang terdiri atas fase adaptasi, pertumbuhan, stasioner, dan kematian.
Fase adaptasi terjadi pada jam ke-0 hingga jam ke-1. Selama waktu tersebut bakteri baru menyesuaikan diri dengan lingkungan yang baru, sehingga sel belum membelah. Fase eksponensial terjadi pada jam ke-1 hingga jam ke-22. Pada fase eksponensial sel-sel bakteri sangat aktif membelah dan metabolisme sel berlangsung cepat.
Gambar 6 Kurva pertumbuhan bakteri T8 dan produksi protease.
9
w a k t u ( j a m ) 0 5 1 0 1 5 2 0 2 5 3 0 3 5 Opt ical den si ty (O D ) 0 . 0 0 . 2 0 . 4 0 . 6 0 . 8 1 . 0 Ak ti v it a s p rotease (U/ m L) 0 . 0 0 1 0 . 0 0 2 0 . 0 0 3 0 . 0 0 4 O p t i c a l d e n s i t y ( O D ) A k t i v i t a s p r o t e a s e ( U / m L )Pertumbuhan bakteri mulai melambat ketika memasuki fase stasioner, yaitu mulai pada jam ke-22. Pada fase stasioner, nutrisi di dalam media mulai berkurang, sehingga kematian sel meningkat. Jumlah sel yang mati semakin meningkat sampai terjadi suatu keadaan dimana jumlah sel yang hidup sama dengan jumlah sel yang mati. Fase kematian terjadi pada jam ke-30. Fase kematian terjadi ketika jumlah substrat menurun di bawah konsentrasi yang dibutuhkan untuk menjaga ketahanan sel, sehingga sel lisis dan mati. Pada bakteri T8, pertumbuhan optimal sel berada pada fase eksponensial.
Pada bakteri sintesis enzim ekstraseluler dalam jumlah terbesar secara normal terjadi pada saat sebelum sporulasi, yaitu pada akhir fase eksponensial dan awal fase stasioner. Keadaan tersebut diduga disebabkan karena pada masa transisi fase eksponensial juga diikuti dengan penurunan sumber karbon dalam medium (Suhartono 1992). Puncak produksi protease alkali termostabil umumnya terjadi pada akhir fase eksponensial sampai akhir fase statis (Kubo
et al. 1988). Menurut Vortuba et al. (1987),
pada genus Bacillus, sintesis protease netral dan alkali biasanya terjadi pada akhir fase eksponensial.
Fase kematian tidak dapat diamati sepenuhnya dengan metode turbidimetrik. Metode turbidimetrik tidak dapat membedakan antara sel yang hidup dan sel yang mati karena metode ini mengukur pertumbuhan berdasarkan kepekatan media. Penurunan absorbansi sel bakteri setelah fase stasioner dianggap sebagai permulaan fase kematian.
Berdasarkan hasil penentuan kurva produksi protease pada Gambar 6, dapat disimpulkan bahwa aktivitas protease berkorelasi positif terhadap pertumbuhan bakteri T8. Semakin besar jumlah sel, maka semakin besar juga aktivitas protease yang dihasilkan. Namun pada waktu inkubasi jam ke-22, aktivitas protease mengalami penurunan. Hasil penelitian menunjukkan bahwa pada jam ke-21, bakteri T8 menghasilkan aktivitas protease yang paling
tinggi, yaitu sebesar 3.8375 x 10-3 U/mL.
Waktu inkubasi dengan aktivitas ptotease paling tinggi tersebut digunakan sebagai waktu inkubasi optimum untuk produksi protease. Tahap akhir dari proses produksi enzim protease dari bakteri T8 adalah isolasi dengan menggunakan sentrifus pada kecepatan 10000 rpm selama 5 menit. Supernatan yang diperoleh merupakan ekstrak kasar protease.
Ekstrak kasar yang diperoleh dimurnikan dengan mengendapkan protein mengunakan garam amonium sulfat dengan tingkat kejenuhan berbeda untuk memisahkan protein enzim dari protein lainnya, sehingga diperoleh fraksi-fraksi protein (Bugg 2004). Pengendapan menggunakan garam didasarkan pada kelarutan protein yang berinteraksi polar dengan molekul air, interaksi ionik protein dengan garam, dan daya tolak menolak protein yang bermuatan sama. Ion garam yang ditambahkan mempengaruhi kelarutan protein. Ketika konsentrasinya rendah, ion-ion ini akan mengelilingi molekul protein dan mencegah bersatunya molekul-molekul protein, sehingga protein larut (salting in). Penambahan garam dengan konsentrasi tinggi akan menyebabkan kelarutan protein menurun (salting out). Molekul air yang berikatan dengan ion-ion garam semakin banyak yang akhirnya menyebabkan penarikan selubung air yang mengelilingi permukaan protein, sehingga menyebabkan protein saling berinteraksi, beragregasi, dan kemudian mengendap. Amonium sulfat merupakan garam yang paling sering digunakan untuk mengendapkan protein karena memiliki daya larut tinggi di dalam air dan relatif tidak mahal.
Amonium sulfat ditambahkan sedikit demi sedikit ke dalam ekstrak kasar pada
suhu 5oC. Aktivitas protease tertinggi
diperoleh pada hasil pengendapan amonium
sulfat 60%, yaitu sebesar 7.7257 x 10-3
U/mL. Aktivitas protease hasil pengendapan dengan amonium sulfat lebih tinggi dibandingkan aktivitas ekstrak kasar protease,
yaitu sebesar 3.8375 x 10-3 U/mL. Menurut
Joshi (2010), aktivitas protease dapat ditingkatkan dengan pengendapan garam amonium sulfat.
Aktivitas Protease Bakteri Termofilik
Aktivitas protease diukur dengan metode Bergmeyer (1983). Prinsip pengukuran aktivitas protease dengan metode Bergmeyer adalah hidrolisis substrat oleh protease menjadi asam amino dan peptida. Substrat yang digunakan adalah kasein (Suhartono 1992). Kasein merupakan protein susu yang terdiri atas fosfoprotein yang berikatan dengan kalsium membentuk garam kalsium yang disebut kalsium kalseinat. Protease yang disekresikan oleh bakteri T8 akan menghidrolisis kasein untuk menghasilkan asam amino. Laju pembentukan asam amino dan peptida sebanding dengan aktivitas katalitik protease. Intensitas warna biru yang
terbentuk akan sebanding dengan konsentrasi kasein yang terhidrolisis. Semakin banyak kasein yang terhidrolisis, maka semakin pekat warna biru yang terbentuk. Tirosin digunakan sebagai standar dalam pengukuran karena tirosin merupakan salah satu asam amino yang paling banyak terkandung dalam kasein dan berperan dalam menentukan timbulnya warna biru saat uji aktivitas dilakukan.
Asam amino yang dihasilkan dari hidrolisis kasein oleh protease, dipisahkan dari protein yang belum terhidrolisis menggunakan asam trikloroasetat (TCA). Asam amino dan peptida akan dilarutkan dengan TCA, sedangkan protein yang memiliki bobot molekul yang besar akan mengendap. TCA juga berfungsi menginaktifkan protease dan menghentikan waktu inkubasi protease. Tahap pemisahan asam amino dan peptida yang terbentuk selama inkubasi dengan protein yang mengendap atau dengan substrat yang belum terhidrolisis dibantu oleh sentrifugasi pada kecepatan 4000 rpm selama 10 menit. Supernatan yang terbentuk melalui tahap pemisahan tersebut merupakan asam amino hasil hidrolisis kasein oleh protease.
Supernatan ditambahkan Na2CO3 agar
pHnya menjadi sekitar 11.5. pH tersebut merupakan pH optimum untuk intensitas dan stabilitas warna biru. Warna yang terbentuk disebabkan oleh pereaksi Folin Ciocalteau. Larutan dibaca pada panjang gelombang 578 nm. Besarnya serapan berbanding lurus dengan konsentrasi asam amino yang terbentuk. Besarnya serapan juga berbanding lurus dengan konsentrasi protein yang terhidrolisis. Aktivitas satu unit enzim dinyatakan sebagai jumlah milimol tirosin yang terbentuk dalam satu menit per mL enzim pada kondisi percobaan (U/mL atau mmol/menit.mL).
Optimasi Protease
Optimasi yang dilakukan pada penelitian ini meliputi pengaruh pH dan suhu terhadap aktivitas protease. Optimasi protease bertujuan menentukan pH dan suhu optimum aktivitas protease. Pada pH dan suhu optimum, protease akan menunjukkan aktivitas yang paling tinggi. Penentuan pH optimum bakteri T8 dilakukan pada pH 6-10. Bufer yang digunakan adalah bufer borat pH 8, bufer Clark & Lubs pH 8-10, dan bufer fosfat pH 6-8. Pengaruh pH pada aktivitas protease ditunjukkan pada Gambar 7. Aktivitas protease paling tinggi terjadi pada
bufer Clark & Lubs pH 8 sebesar
8.6840 x 10-3 U/mL. Aktivitas protease pada
bufer Clark & Lubs pH 8 lebih tinggi dibandingkan pada bufer borat pH 8 dan bufer fosfat pH 8. Pada bufer borat, aktivitas
protease sebesar 7.7257 x 10-3 U/mL dan
pada bufer fosfat pH 8 aktivitas protease
sebesar 6.2638 x 10-3 U/mL.
Perbedaan aktivitas protease pada ketiga bufer disebabkan oleh perbedaan pengaruh senyawa yang terdapat dalam bufer tersebut (Vratyastoma 2006). Senyawa yang dikandung oleh bufer Clark & Lubs meliputi senyawa borat dan garam potasium klorida, sedangkan bufer borat dan bufer fosfat tidak mengandung garam potasium klorida. Senyawa klorida dapat mempertahankan stabilitas protease hingga minggu kedelapan. Senyawa fosfat hanya mampu mempertahankan stabilitas protease hingga minggu keempat (Sutandi 2003).
Protease yang dihasilkan mikroorganisme memiliki aktivitas optimum pada kisaran pH 8-12 (Ghorbel et al. 2003; Singh et al. 2001; Mehrotra et al. 1999).
Bacillus amovivorus menghasilkan protease
yang optimum pada pH 8-8.5 (Sharmin et al. 2005). Menurut Purbowo (1988) B. subtilis menghasilkan protease yang bersifat alkali dan dapat stabil pada kisaran pH 8-9.
Pengaruh pH terhadap aktivitas protease dianalisis secara statistik menggunakan program SAS, kemudian dilanjutkan dengan uji Duncan. Pada hasil analisis statistik, pH berpengaruh terhadap aktivitas protease. Nilai R-square yang diperoleh sebesar 0.957978 menunjukkan bahwa 95.79% keragaman respon protease dapat dijelaskan oleh perlakuan dalam model (pH), sisanya sebesar 4.21% dijelaskan oleh faktor lain di luar model. Pada uji Duncan, nilai yang terbesar ditunjukkan oleh bufer Clark & Lubs pH 8. Hal tersebut menjelaskan bahwa pH yang paling baik untuk aktivitas protease adalah pH 8 yang diperoleh dari bufer Clark & Lubs.
Bufer borat pH 8, bufer Clark & Lubs pH 8, dan bufer Clark & Lubs pH 9 memiliki taraf yang berbeda nyata karena ketiga pH tersebut tidak memiliki Duncan Grouping. Hal tersebut dapat diartikan bahwa penggunaan bufer borat pH 8, bufer Clark & Lubs pH 8, dan bufer Clark & Lubs pH 9 memberikan pengaruh terhadap aktivitas protease. Hasil uji Duncan juga menjelaskan bahwa bufer fosfat pH 7, bufer fosfat pH 8, dan bufer Clark & Lubs pH 10 tidak berbeda nyata karena memiliki Duncan Grouping yang sama. Penggunaan bufer fosfat pH 6 dan bufer fosfat pH 7 juga tidak berbeda nyata karena antara bufer Fosfat pH 6 dan
bufer fosfat pH 7 berada dalam Duncan
Grouping yang sama.
pH lingkungan berpengaruh terhadap kecepatan aktivitas enzim dalam mengkatalisis suatu reaksi. Hal ini disebabkan konsentrasi ion hidrogen mempengaruhi struktur tiga dimensi enzim dan aktivitasnya. Setiap enzim memiliki pH optimum dimana pada pH tersebut struktur tiga dimensinya paling kondusif untuk mengikat substrat. Jika konsentrasi ion hidrogen berubah dari konsentrasi optimal, maka aktivitas enzim secara progresif hilang hingga pada akhirnya enzim menjadi tidak fungsional (Lehninger 1982). Menurut Palmer (1981), aktivitas enzim yang menurun karena perubahan pH disebabkan oleh berubahnya keadaan ion substrat dan enzim. Perubahan tersebut dapat terjadi pada residu asam amino yang berfungsi untuk mempertahankan struktur tersier dan kuartener enzim aktif. Perubahan struktur tersier dapat mengakibatkan sisi hidrofobik yang awalnya tersimpan pada bagian dalam molekul enzim menjadi terbuka, sehingga kelarutan enzim berkurang. Berkurangnya kelarutan enzim dapat menurunkan aktivitas katalitik enzim secara perlahan. Aktivitas enzim dapat ditingkatkan dengan memulihkan reaksi enzimatis pada pH optimumnya.
Aktivitas enzim ditentukan oleh suhu pada saat mengkatalisis suatu reaksi. Pengaruh suhu terhadap aktivitas enzim
dilakukan pada suhu 50oC-90oC. Aktivitas
enzim akan mengalami peningkatan dengan meningkatnya suhu. Pengaruh suhu terhadap aktivitas proteolitik ditunjukkan pada Gambar 8. Aktivitas protease meningkat seiring dengan peningkatan suhu inkubasi, namun setelah suhu optimum aktivitasnya menurun. Suhu optimum protease dari bakteri
T8 adalah 60oC. Pada suhu 60oC, aktivitas
enzim protease sebesar 8.9235 x 10-3 U/mL.
Aktivitas protease menurun setelah suhu
60oC. Hal tersebut disebabkan karena
terjadinya proses denaturasi yang dapat mempengaruhi sifat katalitik pada sisi aktif enzim, sehingga kecepatan reaksinya berkurang. Pada kondisi tersebut substrat juga dapat mengalami perubahan konformasi, sehingga gugus reaktifnya tidak dapat lagi atau mengalami hambatan untuk berikatan dengan sisi aktif enzim.
Penurunan aktivitas protease dapat dipengaruhi oleh beberapa faktor diantaranya pH dan suhu (Naiola & Widhyastuti 2007). Pada suatu reaksi enzimatik, setelah suhu optimum tercapai laju reaksi akan turun.
Penurunan aktivitas protease terjadi karena perubahan struktur enzim yang akan menyebabkan penurunan laju katalitik. Akibat perubahan struktur enzim, sisi aktif enzim mengalami perubahan bentuk sehingga tidak dapat digunakan secara baik dalam mengikat substrat (Sofro 1990). Peningkatan suhu yang melebihi suhu optimum suatu enzim, akan menyebabkan lemahnya ikatan di dalam enzim secara struktural.
Pengaruh suhu terhadap aktivitas protease dianalisis secara statistik menggunakan program SAS, kemudian dilanjutkan dengan uji Duncan. Pada hasil analisis statistik, suhu berpengaruh terhadap aktivitas protease. Hasil yang didapatkan menunjukkan bahwa adanya pengaruh suhu terhadap aktivitas protease. Nilai R-square yang diperoleh sebesar 0.998806 menunjukkan bahwa 99.88% keragaman respon protease itu dapat dijelaskan oleh perlakuan di dalam model (suhu), sisanya sebesar 0.12% dijelaskan oleh faktor lain di luar model. Pada uji Duncan, nilai yang
terbesar ditunjukkan oleh suhu 60oC. Pada
perlakuan suhu, suhu yang paling baik untuk
aktivitas protease adalah suhu 60oC. Hasil uji
Duncan menjelaskan bahwa antara suhu 50oC
dan suhu 60oC tidak berbeda nyata karena
memiliki Duncan Grouping yang sama. Hal
serupa juga ditunjukkan antara suhu 50oC dan
suhu 70oC. Suhu 80oC dan suhu 90oC
memiliki taraf yang berbeda nyata karena kedua suhu tersebut tidak memiliki Duncan
Grouping.
Menurut Murray et al. (2003), kenaikan suhu akan meningkatkan kecepatan reaksi yang dikatalisis enzim, namun peningkatan kecepatan reaksi hanya pada kisaran suhu tertentu. Kecepatan reaksi akan meningkat sejalan dengan kenaikan suhu dan peningkatan kecepatan reaksi tersebut disebabkan oleh peningkatan energi kinetik molekul-molekul yang bereaksi. Peningkatan energi lebih jauh akan memutuskan ikatan hidrogen dan hidrofobik lemah yang mempertahankan struktur sekunder-tersier dari enzim disertai dengan menurunnya aktivitas katalitik enzim tersebut. Pada suhu maksimum, enzim akan terdenaturasi karena struktur protein terbuka dan gugus nonpolar yang berada di dalam molekul menjadi terbuka lebar, kelarutan protein di dalam air yang polar menjadi turun, sehingga ektivitas enzim juga akan turun (Lehninger 1982). Suhu tinggi juga dapat mempengaruhi konformasi substrat, sehingga mengalami hambatan dalam memasuki sisi aktif enzim (Suhartono 1992).
Gambar 7 Optimasi pH terhadap aktivitas protease.
Gambar 8 Optimasi suhu terhadap aktivitas protease.
SIMPULAN DAN SARAN
Simpulan
Dua isolat dari sumber air panas Sipoholon, sebelas isolat dari sumber air panas Tangkuban Perahu, dan dua isolat dari sumber air panas Ciseeng dapat membentuk daerah bening di sekitar koloninya. Isolat T8 yang berasal dari Tangkuban Perahu dengan indeks proteolitik sebesar 33.5 digunakan untuk uji selanjutnya. Aktivitas tertinggi bakteri T8 ditunjukkan pada sampel yang
diambil pada jam ke-21 sebesar
3.8375 x 10-3 U/mL. Aktivitas protease
meningkat dengan pengendapan amonium
sulfat 60%, yaitu 7.7257 x 10-3 U/mL. Hasil
analisis secara statistik dengan program SAS yang dilanjutkan dengan uji Duncan menunjukkan bahwa pH dan suhu memberikan pengaruh terhadap aktivitas protease. Aktivitas protease tertinggi pada pH
8 bufer Clark & Lubs dan suhu 60oC.
Saran
Penelitian lebih lanjut perlu dilakukan untuk mengetahui pengaruh inhibitor terhadap aktivitas protease, uji ketahanan enzim pada suhu tinggi, dan uji konsentrasi protein untuk mengetahui aktivitas spesifik protease. Selain itu, perlu dilakukan identifikasi bakteri terhadap bakteri T8.
DAFTAR PUSTAKA
Aguilar A, Ingemansson T, Magnien E. 1998. Extremophile microorganisms as cell factories. Sci Technol Aliment 2:367-373.
Bergmeyer HU. 1983. Methods of Enzymatic
Analysis. Weinstein: Verlag Chemie.
Brock TD. 1986. An overview of the thermophiles. dalam Brock TD.
Thermophiles: General Molecular and Applied Microbiology. New York: J
Wiley.
Bugg TDH. 2004. Introduction to Enzyme
and Coenzyme Chemistry. Oxford:
Blackwell.
Cappucino JG, Sherman N. 1983.
Microbiology. Massachusetts:
Addison-Wesley.
Edwards C. 1990. Microbiology of Extreme
Environtments. New York:
McGraw-Hill.
Fardiaz S. 1988. Fisiologi Fermentasi. Bogor: PAU-IPB.
Ghorbel B, Sellami KA, Nasri M. 2003. Stability studies of protease from
Bacillus cereus BG1. Enzyme Microb Technol 32:513-518.
Hartoto M. 2003. Pembuatan yoghurt sinbiotik dengan menggunakan kultur campuran Streptococcus thermophilus,
Bfidobacterium, dan Lactobacillus casei galur Shirota. [skripsi]. Bogor:
Fakultas Teknologi Pangan dan Gizi, Institut Pertanian Bogor.
Janzen JJ, Bishop JR, Bodiene AB. 1982. Relationship of protease activity rushelf life of skim and whole milk. J
Dairy Sci 65:2237-2240.
Joshi BH. Purification and characterization of a novel protease from Bacillus
Firmus. 2010. J Appl Sci Re
6(8):1068-1076.
13
0 0.001 0.002 0.003 0.004 0.005 0.006 0.007 0.008 0.009 0.01 6 7 8 9 10 Akti v ita s Pr o te a se (U /m L ) pH Bufer fosfat Bufer Clark & Lubs Bufer borat 0 0.001 0.002 0.003 0.004 0.005 0.006 0.007 0.008 0.009 0.01 50 60 70 80 90 Ak tivi tas P rote ase ( U /m L) Suhu (oC)Kubo M, Murayama K, Seto K, Imanaka T. 1988. Higly thermostable neutral protease from Bacillus
stearothermophilus. J Ferment
Technol 66(1):13-17.
Labeda DP. 1990. Environtmental
Biotechnology: Isolation of Biotechnological organisms from nature. New York: McGraw-Hill.
Lehninger AL. 1982. Dasar-dasar Biokimia. Thenawidjaja M, penerjemah; Jakarta: Erlangga. Terjemahan dari: Principles
of Biochemistry.
Madigan MT, Marrs BL. 1997. Extremophiles. Sci Am 82-87.
Matjik AA, Sumertajaya M. 2002.
Perancangan Percobaan dengan Aplikasi SAS dan Minitab. Bogor:
Jurusan Statistika FMIPA IPB. Mehrotra S, Pandey PK, Gaur R, Damwal
NS. 1999. The production of alkaline protease by a Bacillus species isolate.
Microb Technol 67:201-203.
Murray RK, Granner DK, Mayes PA, Rodwel VW. 2003. Biokimia Harper. Hartono A, penerjemah; Jakarta: EGC. Terjemahan dari: Harper’s
Biochemistry.
Naiola E, Widhyastuti N. 2007. Semi purifikasi dan karakterisasi enzim protease Bacillus sp. Hayati 13:51-56. Oktafianti F. 2005. Aktivitas protease
Bacillus natto dari empat jenis natto komersial [skripsi]. Bogor: Fakultas Teknologi Pertanian, Institut Pertanian Bogor.
Palmer T. 1981. Understanding Enzymes. England: Ellis Horwood.
Purbowo Y. 1988. Protease dari Bacillus
Subtilis dan penerapannya sebagai
pengempuk daging [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Ramadzanti A. 2006. Aktivitas protease dan kandungan asam laktat pada yoghurt yang dimodifikasi Bifidobacterium
bifidum [skripsi]. Bogor: Fakultas
Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. 1998. Molecular and Biotechnological aspects of microbial
proteases. Microbiol Mol Biol
62(3):597-635.
Sadikin M. 2002. Biokimia Enzim. Jakarta: Widya Medika.
Salem SR, Shabeb SA, Amara AA. 2009. Optimization of thermophilic protease production in Bacillus mixed cultures under mesophilic conditions. J Agric
Sci 5(3):375-383.
Sanfitri EH. 2007. Amplifikasi gen 16S-rRNA bakteri termofilik dari sumber air panas, Gunung Pancar Bogor [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Sharmin S, Hossain T, Anwar MN. 2005. Isolation and characterization of a protease producing bacteria Bacillus
amovivorus and optimization of some
factors of culture conditions for protease production. J Biol Sci 5(3):358-362.
Sian LW. 1992. Mempelajari aktivitas protease Bacillus licheniformis galur Gibson NCTC 10341 pada fermentasi terkontrol menggunakan limbah cair tahu [skripsi]. Bogor: Fakultas Teknologi Pertanian, Institut Pertanian Bogor.
Singh J, Batra N, Sobti RC. 2001. Serine alkaline protease from a newly isolated Bacillus sp. Process Biochem 36:781-785.
Sofro ASM. 1990. Biokimia. Yogyakarta: UGM Pr.
Sokatch JR. 1973. Bacterial Physiology and
Metabolism. New York: Academic Pr.
Stanburry PF, Whitaker A. 1984. Principle of
Fermentation Technology. New York:
Pergamon Pr.
Suhartono MT. 1989. Enzim dan
Bioteknologi. Bogor: IPB Pr.
Suhartono MT. 1992. Protease. Bogor: IPB Pr.
Susanti E. 2003. Isolasi dan karakterisasi protease dari Bacillus
subtilis 1012M15. Biodiversitas
4(1):12-17.
Sutandi C. 2003. Analisis potensi enzim protease lokal [skripsi]. Bogor:
Fakultas Teknologi Pertanian,
Vortuba J, Pazlarova J, Dvaraova M, Vanatalu K, Vachava L. 1987. External factors involved in regulation of an extracellular proteinase synthesis
in Bacillus megaterium. Appl
Microbiol Biotechnol 26:373-377.
Vratyastoma AK. 2006. Optimasi produksi dan karakterisasi enzim protease dari
Bacillus natto [skripsi]. Bogor:
Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Ward OP. 1983. Properties of microbial
proteinase. dalam Forgaty W
Microbial Enzyme and Biotechnology.
London: Applied Sci.
Widhyastuti N, Naiola E. 2002. Isolasi, seleksi, dan optimasi produksi protease dari beberapa isolat bakteri.
Berita Biologi 6:467-473.
Winarno FG. 1985. Enzim Pangan. Jakarta: Gramedia.