Typically, however, the parameres retain only a narrow basal connection with the aedeagus (F), and they may be so far displaced laterally (G, H) that they appear to be independent appendages of the ninth segment. Else (1934) claimed to have traced the phallic crudiments from the appendage buds of the tenth abdominal segment in the embryo of Melano.

NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 7 and two pairs of appendages cannot be developed on one segment
It would therefore seem very probable that the primitive male insects had a pair of pinnae on the genitalia, and that the oviducts of the female opened separately, as they still do in the Ephemeroptera. Admittedly, it is difficult to imagine that the large and complex genital organs of the higher insects could have been developed from a few simple pinnae.
10 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 have no evidence as to what may have been the primitive position of
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS II
PROTURA, COLLEMBOLA, DIPLURA
12 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 They are an early offshoot from the hexapod hue, which proved to be
THYSANURA
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 1
14 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 entia (Vd) open into the base of the distal section (Gpr), the latter
EPHEMEROPTERA
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 1 5
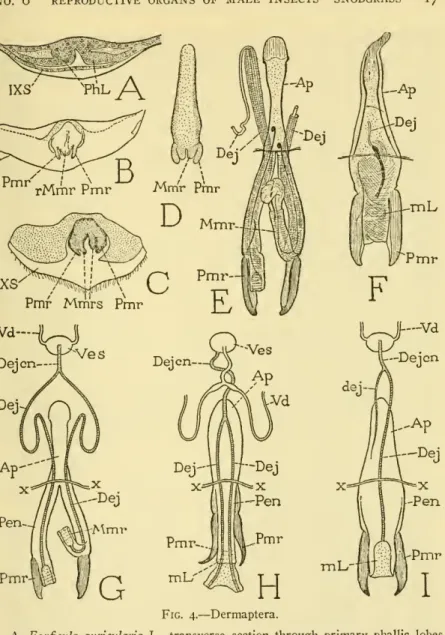
DERMAPTERA
The mesal lobe (Mmr) is a hollow continuation of the stalk that gives access to the enclosed canal. The long ejaculatory ducts of the peniles (G, Dej) unite in a short common duct (Dejcn), which connects a vesicle (Ves) that receives the two vasa deferentia (Vd). Hemimerus talpoides has a single genital organ (Fig. 4H) with two parameres, but the mesomeres are united in a common median lobe {mL), comparable to the aedeagus of the higher insects, except that it gives exit to the two primary ducts.
The two ducts unite in a short duct that joins the vesicle of the earlier formed vasa deferentia. In the development of the higher insects, on the other hand, the phallic lobes are never penetrated by tubes; the common ejaculatory duct is formed as an ectodermal ingrowth between the bases of the globe bone, and, when the lath unit forms anaeedeagus, the duct discharges through the aedeagal lumen.
ISOPTERA, EMBIOPTERA, ZORAPTERA, PLECOPTERA, PSOCOPTERA
6 THE REPRODUCTIVE ORGANS OF MALE INSECTS— SNODGRASS I9lobes themselves, and that these lobes themselves represent a pair of primitive lobes, and that these lobes represent a pair of primitive feathers like those of the Ephemeroptera. On the other hand, the development of the lateral branches of the parameres of the phallic rudiments seems to be related to the penile rudiments of Dermapter with the primary phalliclobe of higher insects. The internal parameres are probably offshoots of the external parameres, but as, to the writer's knowledge, the development of the organs has not been studied, we cannot be certain of their homology.
However, Badonnel (1934) notes that since the parameres and aedeagus are branches of the same trunk, they may be considered lobes of a common origin. A full account of the internal and external genital organs of male Psocoptera, mating of the sexes and insemination of the female with a spermatophore is given by Klier (1956).
ORTHOPTERA
The sternum of the abdomen, and each rudiment divides into an adorsal and a ven-abdominal sterna, and each rudiment divides into an adorsal and a ven-.
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS— SNODGRASS 21
22 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 1 35 writers, it is probable that in all the orthopteroid insects the external
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 23
HEMIPTERA (RHYNCHOTA)
24 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 back to the Thysanura to see that the phallic rudiments have no con-
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 25
26 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 27
28 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 mere and a paramere (B) ; later the mesomeres unite to form the
COLEOPTERA
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 29
30 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL, I35
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 3I
MEGALOPTERA
The two segments of the holometabolous paramers were called hasimeres and distimeres by Crampton (1942), but linguistically telomere should be preferred to the hybrid 'distimere'. However, the close association of the prehensiles with the aedeagus in some adult Megaloptera leaves little doubt that the three parts are derived from common rudiments, as in other insects. Ventral from the base of the parameres, two narrow sclerotic ridges converge in the inferior wall of the aedeagus (C) and follow the edges of a medial groove of the latter to the distal end.
The phallotreme is a transverse opening at the apex of the aedeagus, and opens from a large, probably eternal, endophallicsac. The genitalia of Agulla adnixa (fig. 9 D) is similar to that of the species just described, but the parameres and the aedeagus are broadly continuous at their bases.
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 33
34 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 In Corydalus cornutus the parameres are highly developed, but
The parameres are articulated on the posterior margin of the ninth abdominal segment (Fig. loA, E) and are muscularized from the sternum (C, E). Because of the apparent relation of the parameres to the ninthabdominal segment, viewed from the side, the parameres are commonly regarded as gonopods of this segment, and. As the larva begins to change into the pupa, the genital pouch flattens, bringing the parameres and aedum to the outer surface of the body.
However, the base of the aedeagus becomes surrounded by a circular ingrowth of the body wall, which is the aedeagal pocket of the adult. This account by Zander clearly shows that the trichopteros claspers are parameres derived with the aedeagus from a pair of primary phallic lobes, and that their lateral position in the adult is secondary.
36 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35
LEPIDOPTERA
The genitalia of Lepidoptera have many features in common with those of Trichoptera, particularly the presence of large lateral parameres ("harpes valvae") articulated on the ninth abdominal segment and the partial envelopment of the aedeagus in a pouch of the chamber wall genital. In addition, the bases of the parameres may join in an interparamere bridge behind the edge of the ninth sternum. In some lepidoptera, as in Carpocapsa (fig. iiA), although the parameres (Pnir) are laterally attached to the ninth abdominal sternum, their bases converge medially behind the sternum (B) and are articulated on a small median plate ( BP) on the floor of the genital chamber.
From this plate an arm (full) extends upwards and forks around the mouth of the aedeagal pouch.
38 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35
VUIT
Otherwise, the plate supporting the aedeagal sac is quite comparable to the basal plate of the aedeagus in Homoptera in which the parameres are articulated on it (fig. 6B). The flexor muscles of the lepidopteran parameters are shown by Forbes (1939) on the inner face of the supporting plate of the aedeagus; in Carpocapsa these muscles (fig. 11 B) arise in the branches of. The arm apparently serves as a support for the movement of the aedeagus and is here called the aedeagal fulcrum.
II D), the bases of the parameres are united in an interparameral bridge {pmB), located close to the margin of the ninth sternum, as in the trichopteron Neuronia (Fig. 10 B). The rudiments of the flaps (flaps), he says, are formed later than the odeagal lobes towards the end of the larval period as thickenings of the lateral walls of the genital chamber.
40 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 veloped from a single pair of phallic rudiments as in other insects
MECOPTERA
Most researchers of mecopteran genitalia, including Ferris and Rees (1939), Grell (1942) and Tjeder (1956), considered the panorpid claspers to be the gonopods of the ninth abdominal segment, consisting of "gonocoxytes" and "gonostyli". In this they agree with the terminology previously used by the writer (1935), which is here discarded as untenable. Apparently no studies have been done on the development of mecopteran genitalia, but when we turn to PanorpatoMeropea, we find that the condition strongly suggests that the claspers are phallic parameres. In merope tuber the claspers are long, slender, bisegmented arms (Fig. 12D); their bases converge above the ninth sternum and are united above and below the aedeagus (E, Aed), which is mostly invaginated between them.
I35united, and Michener (1944) says that the prehensiles of Apterobittacus, Bit-united, and Michener (1944) says that the prehensiles of Apterobittacus, Bittacus, and Panorpa are fused both above and below the base of the aedeagus and surround a sexual organ. foramen", but he seems to confirm no. The aedeagus of the Merope tuber (Fig. 12F) is an oval, capsule-like structure with a pair of long, dorsal basal arms and pointed lateral lobes.

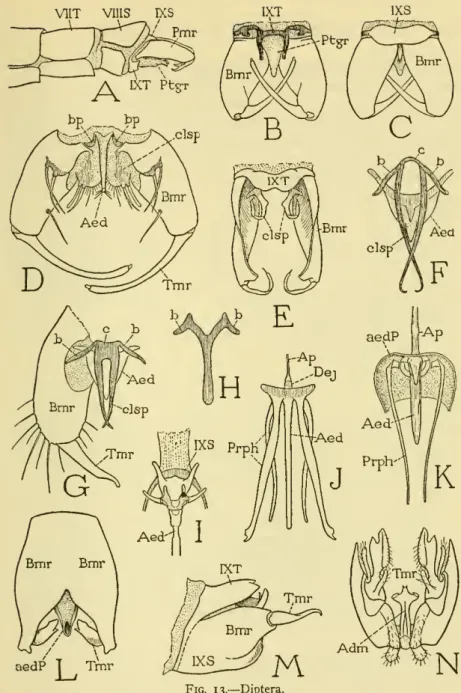
DIPTERA
According to Christophers (1922), inversion in Culex fatigans is complete 24 to 48 hours after the emergence of the adult. Their irregular tergum develops into a pair of processes below the bases of the claspers (B); the small sternum (C) slightly overlaps the upper surfaces of the claspers. The anatomical unity of the genital parts in the mosquito, together with their development from a single pair of rudiments, leaves no doubt that the true parameres are the large external ramps.
Christophers (1922) showed that the entire genital complex of Anopheles is developed from a single pair of primary phallic lobes. 14A, PhL) are formed behind the ninth abdominal sternum as the larva enters its last instar. The lobes are initially buried in epidermal pockets, but in the bud they invert and become relatively large (B).
46 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL, I35 dropodites," each of which is finally divided into a large basal segment
6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 47 These accounts of the development of the phallic organs in two.
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 47 These accounts of the development of the phallic organs in two
HYMENOPTERA
According to Zander (1900), the ring forms relatively late in development from the epidermis at the base of the primary phallus. However, it becomes an essential part of the adult organ, as the external phallic muscles from the ninth abdominal segment are inserted into it. In some cleistogasters, the distal part of the paramere may be flexible or even folded at the basal part (N), but only in a few such cases is the telomere equipped with muscles.
The free part of the aedeagus projects from between the bases of the parameres (Fig. 15A, G, L, Aed) and assumes various shapes. These aedeagal bars are called "parameres" by Beck (1933), who states that they "are the gonapophyses of the ninth sternite." Other writers more commonly.
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 1 35
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 5
The volsellae musculature seems to show that in insects special muscles can be developed where there is a need for them. The development of the hymenopterousphallus has been described by Seurat (1899) in the braconid Dorcytes; by Michaelis (1900) in Apis; by Zander (1900) in Vespa, Bomhus and Apis; by Boulange (1924) in Sirex; and by Snodgrass (1941) in Vespiila and Apis. Below the disk in a pocket in the integument in the enclosed stage are two small primary phallic lobes (C, PJiL).
The much larger phallus of the adult (F, iPhl) will then develop from the pupal organ (pPhl) and finally assume the mature structure (G). The entire genital complex of the adult thus originates from a single pair of minutophallic rudiments.
SUMMARY
In insects other than Ephemeroptera, a secondary median ectodermal ejaculatory duct grows inward between the bases of the phallic lobes. The generally accepted idea that the phallic lobes represent a pair of appendages of the ninth or tenth abdominal segment must be accepted. If the parameres represent the coxae of the lost legs, they must theoretically be regenerated from the penae they once bore.
Given the known facts about the ontogenetic development of the phallic organs, it takes a strong imagination to visualize their development from a pair of legs. Other authors describe the phallic lobes as making their first appearance on the nymph or larva after the abdominal limb buds of the embryo have disappeared.
56 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35
NO. 6 REPRODUCTIVE ORGANS OF MALE INSECTS — SNODGRASS 57
58 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I35 Lindsay, Eder
1953- On the post-embryonic development of the male genitalia (external and internal) of Tribolitim castancum Herbst (Coleoptera, Tenebrionidae).
60 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL, I35 TjEDER, B