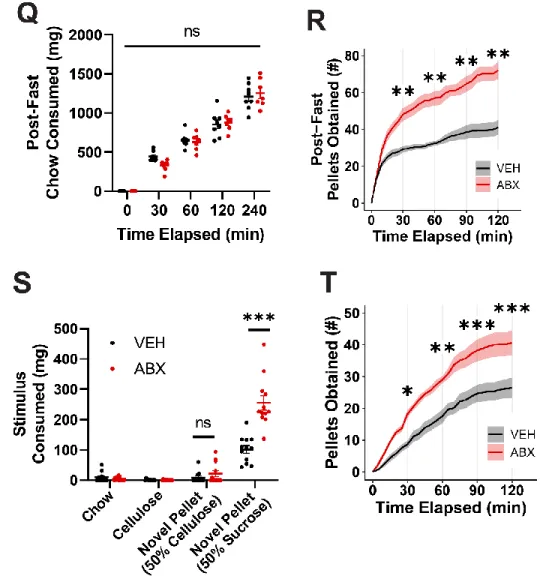
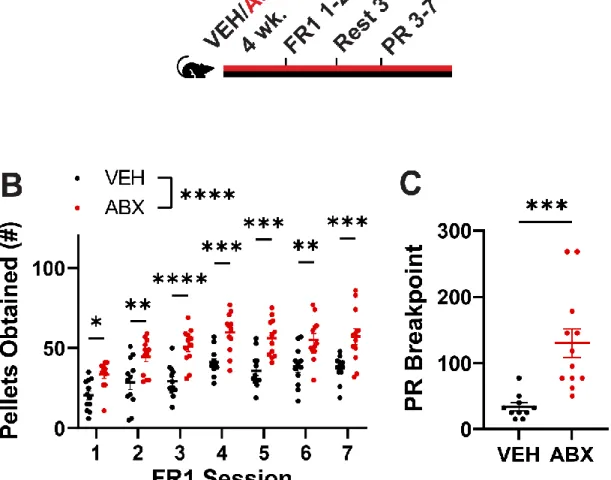
Operant conditioning tests show that the motivation to pursue high-sucrose rewards is enhanced in mice with reduced microbiota. The mesolimbic brain area activity induced by consumption of high sucrose pellets is increased in antibiotic-treated mice. Enteric bacteria from the family S24-7 and the genus Lactobacillus were identified by differential antibiotic treatment and fecal microbiota transplants as correlating with the reduction in high sucrose pellet consumption.
Indeed, colonization of vancomycin-treated mice with a mixture of S24-7 and Lactobacillus johnsonii reduces overconsumption of high-sucrose pellets in a restricted-access binge model.
GUT MICROBIOTA OF MUS MUSCULUS
HOST-GUT MICROBIOTA RELATIONSHIPS IN MUS MUSCULUS
Mouse models of disease that mirror humans often show changes in the composition of the gut community.20,21 However, unraveling whether changes in the microbiota contribute to disease etiology or are a result of the disease state itself is a formidable challenge and an opportunity for further discovery. Among these secreted molecules are short-chain fatty acids (SCFAs), which are reported to reduce food intake through central signaling pathways,26,27 regulate the activation state of microglia,28 and act through fatty acid receptors to control host fat.29 Other small organic molecules produced exclusively by Gut microbes, including 4-ethylphenol and indole sulfate, exacerbate anxiety-like behavior in rodents.30,31 Some pathogenic microbes, such as In addition to the production of active molecules, antigens produced by microbiota constantly induce host immune responses through innate and adaptive pathways.
Finally, the vagus nerve represents a direct line of communication between the gut microbiota and activity in the central nervous system.
EXPERIMENTAL METHODS TO STUDY THE GUT MICROBIOTA
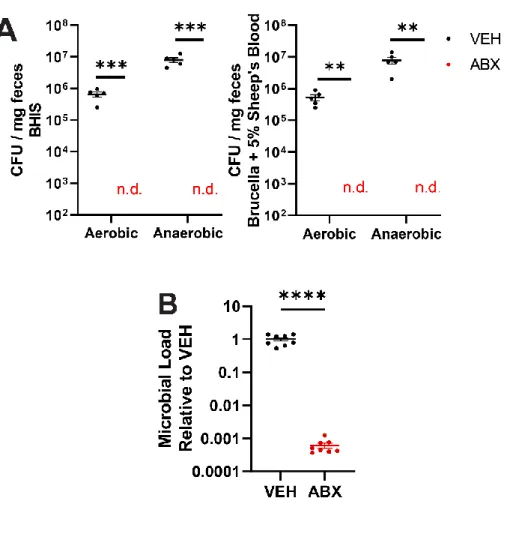
Broad-spectrum oral antibiotics significantly reduce gut microbial loads and produce systemic changes that broadly reflect the GF state.18 Similar to GF mice, ABX depletion can be followed by FMT to determine whether the gut microbiota is sufficient to transfer donor phenotypes. Advances in high-throughput sequencing and taxonomic classification technology have advanced the study of the gut microbiome. Absence of gut microbiota increases anxiety-like behavior and neuroendocrine response to acute stress in rats.
The role of gut microbiota in host appetite control: bacterial growth to animal feeding.
REGULATION OF FEEDING AND FOOD REWARD IN MUS MUSCULUS
- INTRISIC AND EXTRINSIC INFLUENCES ON FEEDING BEHAVIOR All life requires energy. Animals have developed strategies to seek and consume food
- HEDONIC FEEDING AND CONSUMPTION INDUCED BY PALATABLE FOOD Foods which provide pleasure upon consumption will be readily eaten by mice in the
- EXPERIMENTAL METHODS TO STUDY FEEDING AND FOOD-MOTIVATED BEHAVIOR
- GUT MICROBIOTA EFFECTS ON FEEDING BEHAVIOR
Other peripheral mechanisms that regulate short-term intake include mechanosensitive neurons in the gastrointestinal tract and gut-produced cholecystokinin (CCK) and glucagon-like peptide 1 (GLP-1), all of which are reported to act through vagal pathways to reduce intake. of food. For some research questions, it is necessary to induce a strong food intake response in the absence of a palatable stimulus. Genetic differences in the behavioral organization of binge eating, conditioned food reward and compulsive eating in C57BL/6J and DBA/2J strains.
High-fat overeating increases evoked dopamine release and decreases dopamine uptake in the Nucleus Accumbens.
GUT MICROBIOTA SUPPRESS FEEDING INDUCED BY PALATABLE FOODS
Gut microbiota suppress feeding induced by palatable foods
SUMMARY
RESULTS
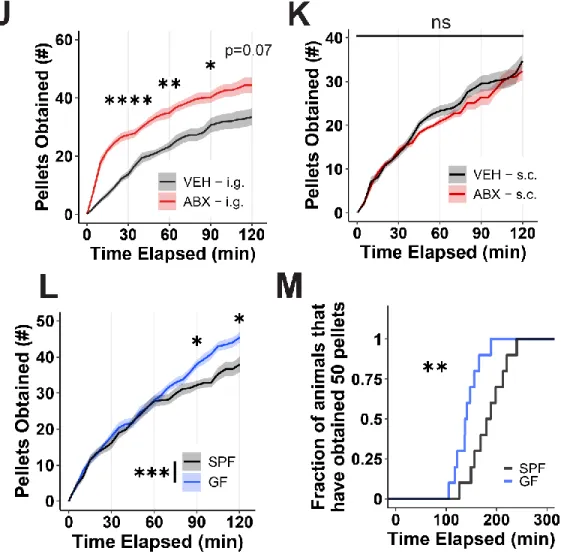
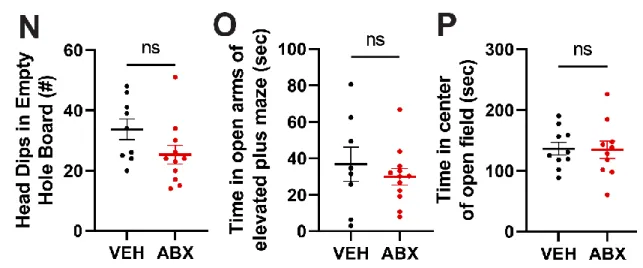
Our data show that the absence of a gut microbiota in mice results in overconsumption of the high-sucrose pellet. Taken together, a complex gut microbiota is sufficient to suppress feeding induced by a high-sucrose diet in mice. VEH and ABX rats that did not receive high-sucrose pellets showed no differences in neural activity in the same regions (Figure S3A-B).
Loss of function suggests that A and V-sensitive microbial taxa inhibit high-sucrose pellet consumption in mice.
DISCUSSION
Significance calculated via two-way repeated measures ANOVA using 30-min time points followed by Šidák's multiple comparisons test. Significance calculated via two-way repeated measures ANOVA using 30-min time points followed by Tukey's multiple comparisons test (within time points). Significance calculated via two-way repeated measures ANOVA using 30-min time points followed by Tukey's multiple comparisons test (within time points).
Time × Treatment interaction effect significance calculated via two-way repeated measures ANOVA using 30-min time points. Treatment effect significance calculated via two-way repeated measures ANOVA at 30-min time points followed by Šidák's multiple comparison test. Significance calculated via two-way repeated measures ANOVA at 30-min time points followed by Šidák's multiple comparison test.
Significance calculated via two-way repeated measures ANOVA at 30 minutes. time points followed by Šidák's test of multiple comparisons. Significance calculated via two-way repeated measures ANOVA using 30-min time points followed by Tukey's multiple. Significance calculated via two-way repeated measures ANOVA using 30-min time points followed by Dunnett's multiple comparison test for VEH (within time points).
Significance calculated by Kruskal-Wallis one-way ANOVA followed by Dunn's multiple comparisons test with VEH. Significance calculated by two-way repeated measure ANOVA using 30-min time points followed by Dunnett's multiple. Significance calculated by Kruskal-Wallis one-way ANOVA followed by Dunn's multiple comparisons test with VEH-FMT.
Significance calculated via one-way ANOVA followed by Dunnett's multiple comparison test with VEH - FMT.

ACKNOWLEDGEMENTS
Resource availability
For c-Fos analysis of NPY+ neurons in the ARC, VEH, ABX, and overnight fasted mice were removed from their home cages and euthanized. To reduce the effects of neophobia, mice were housed overnight with an automated pellet dispenser (Feeding Experimentation Device 2.0, (FED2)) in the "off" condition and acclimated to 100 mg (five pellets) of the high-dose pellets. sucrose content (5TUL, Test Diets, Cat#1811142) in their home cage. On four occasions, representing less than 1% of all retrieval events recorded for this cohort (4/493, 2 VEH and 2 ABX), mice were observed to retrieve a pellet from the cell.
To reduce the effects of neophobia, rats were placed on 200 mg of high-fat diet (HFD). To reduce the effects of neophobia, rats were placed with 1 ml of Ensure® chocolate (Abbott Nutrition, Cat#53623) provided in a glass container. Experimental rats were housed in a home and adapted to 100 mg of high-sucrose pellets the day before the start of FR1 training.
Non-fixed experimental mice were habituated to the test chamber (an empty cage without bedding) for 10 min prior to the introduction of a bolus of pre-weighed edible stimuli in a glass dish – standard chow, pure microcrystalline cellulose pellets (5TUW, Test Diets, Cat# 1812939), a modified formulation of the high sucrose pellets in which the sucrose was replaced by microcrystalline cellulose (Test Diets), or the high sucrose pellets used during the study (5TUL, Test Diets, Cat#1811142) . To reduce neophobia, mice were given a small amount of each edible stimulus in their cage the day before testing (100 mg/mouse) and housed in a glass dish overnight. For behavioral testing, habituated mice were placed in the testing chamber for 10 min before being given a pre-weighed bolus of approximately 1.5 grams of high-sucrose pellets in a glass dish and allowed to consume freely for 1 h.
Experimental mice were housed in a single house and given ad libitum food during VEH/ABX treatment. In the unbaited test, holes were empty, mice were videotaped, and the number of spontaneous head dips was measured by an investigator blind to treatment group as a metric for exploratory behavior.
Quantification and statistical analysis
Supplemental Information
Germ-free C57BL/6J mice are resistant to high-fat diet-induced insulin resistance and have altered cholesterol metabolism. Up-regulation of intestinal type 1 taste receptor 3 and sodium glucose luminal transporter-1 expression and increased sucrose intake in gut microbiota-null mice. In Mood- and anxiety-related phenotypes in mice Neuromethods., T. Use of the elevated plus maze as a test of anxiety-related behavior in rodents.
In mood and anxiety-related phenotypes in mice: characterization using behavioral tests Neuromethods., T. Behavioral and neurochemical consequences of chronic gut microbiota depletion during adulthood in the rat. Comparative evaluation of microbiota engraftment after faecal microbiota transfer in mouse models: age, kinetics and microbial status matter. An open source device for measuring food intake and operant behavior in rodent home cages.
Differential effects of fasting and leptin on proopiomelanocortin peptides in the arcuate nucleus and nucleus of the solitary tract. Gut Microbiota Influences Central Levels of Brain-Derived Neurotropic Factor and Behavior in Mice. Nutrient Sensing in CD11c Cells Alters Gut Microbiota to Regulate Food Intake and Body Mass.
Bacterial community monitoring of human gut microbiota reveals increased lactobacilli in obese patients and methanogens in anorexic patients. Gut microbiota and metabolites in patients with anorexia nervosa. Microbial, biological and psychiatric profiling in relation to binge eating disorders: A cross-sectional study in obese patients.
S upplemental Tables
CONTEXTUALIZATION AND CONCLUSIONS
The biological processes that regulate food intake and the perception of pleasure are vast and complex. The results in this thesis demonstrate that the gut microbiota has a significant influence on the expression of eating behavior that occurs in response to palatable food, providing a new frontier for exploring peripheral influences on feeding and possibly generalized reward. Our findings are supported and extended by a recently published study comparing the intake profiles of antibiotic-induced microbiota-depleted mice given concurrent access to standard chow and a high-fat, high-sugar (HFHS) diet.1 Consistent with our data, the authors argue , that antibiotic-treated mice overconsume the HFHS diet compared with vehicle-treated animals.
However, our findings provide broad innovation in the scientific field beyond reporting simple binge eating behavior in ABX mice. Additionally, operant conditioning experiments provide evidence that intrinsic motivation to obtain a high-sucrose reward varies based on microbiota status, and we performed differential antibiotic treatment and 16S RNA profiling followed by targeting experiments of microbial reconstitution to identify candidate bacterial species that directly mediate the change in host behavior. Furthermore, we tested foods beyond high-sugar stimuli and found a persistent hyperphagic response of ABX mice to multiple palatable foods, including a high-fat diet and Ensure®.
Indeed, microbiota influences have been reported in animal tests of depression, anxiety, autism, addiction, and reward processing.2–7 Curiously, many psychiatric and metabolic diseases in humans are accompanied by consistent shifts in gut microbiota community composition,8,9 suggesting that disturbances in the microbiota may influence disease etiology or one day be used in a diagnostic manner. Prebiotics, probiotics, fecal transplants, genetically engineered microbial strains designed to deliver therapeutic agents, and molecules designed to remove toxic microbial byproducts have been investigated for mood disorders, psychiatric diseases, metabolic syndromes, local intestinal inflammation and for the elimination of antibiotic-resistant opportunistic pathogens.10 While initial findings in this space appear promising, and federal regulatory agencies have begun to grant approval of microbiota consortia for the treatment of disease, additional basic research is needed to understand the fundamental relationships that the gut microbiota linked to the health of the host. . This thesis provides ground evidence of a direct link between the gut microbiota and the behavioral response to palatable food, the dysregulation of which can contribute to disordered eating habits, obesity and additional adverse effects on human health.