How simple the architecture of the pyramids seems, if we have to understand the architecture. These linked data provide opportunities for constraining estimates of the possibilities for life in the history of Earth and elsewhere in the universe. LMW conceived the project, designed the study, prepared the data and analyses, and wrote the manuscript.
Draft genome sequence of Ardenticatena maritima 110S, a thermophilic nitrate- and iron-reducing member of the Chloroflexi class Ardenticatenia. LMW helped to conceive the study, analyze the data and write the manuscript. One of the most significant innovations in the history of life was the invention of oxygenic photosynthesis by cyanobacteria.
The first part of this dissertation therefore deals with the nature of the early Earth, before the development of oxygenic photosynthesis and the rise of oxygen.

ELECTRON DONOR LIMITATION OF THE BIOSPHERE BEFORE THE ORIGIN OF OXYGENIC PHOTOSYNTHESIS
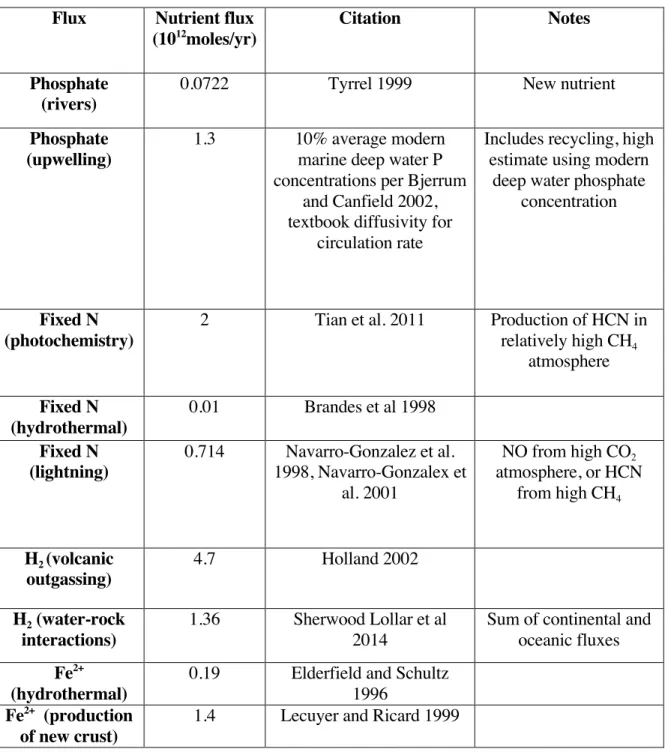
In the modern ocean, fixed nitrogen is supplied primarily by biological nitrogen fixation using the enzyme nitrogenase (Canfield et al. 2010). Prior to the evolution of nitrogenase, and potentially throughout the Archaean, fixed nitrogen may have been provided primarily by abiotic nitrogen fixation processes (Navarro-Gonzalez et al. 2001). 2002, Saito et al. 2003), this has not been demonstrated for anoxygenic organisms, which are expected to promote primary productivity prior to the development of oxygenic photosynthesis.
Although iron oxidation was considered an important energy source for the early biosphere, its relative contribution to primary productivity rates is disputed (e.g. Kharecha et al. 2005, Canfield et al. 2006) and may have been negligible (Box 1). . Methanogenesis and acetogenesis are thought to be among the earliest metabolisms to evolve (Battistuzzi et al. 2004), and thus may have been responsible for promoting productivity in the earliest stages of life on Earth. The Wood-Ljungdahl (reverse acetyl-CoA) pathway is thought to be much more ancient, possibly present in the last universal common ancestor (Lane et al. 2010).
This is consistent with some phylogenetic and molecular clock analyzes of the evolution of the nitrogenase family (e.g., Boyd et al. 2011), but further evidence will be needed for a reliable assessment. This seems an unlikely situation—photopherotrophy appears to have been an ecologically acceptable niche in the Archean ocean and is biochemically feasible given its presence in modern organisms (e.g., Widdel et al. 1993). In the modern ocean, dissolved silica concentrations are low due to the precipitation of silica biominerals, mainly by eukaryotic algae; the Archean ocean would have been dominated by abiotic precipitation of silica prior to the development of these organisms, resulting in a more silica-saturated ocean (Stefurak et al. 2014).
TIMESCALES OF OXYGENATION FOLLOWING THE EVOLUTION OF OXYGENIC PHOTOSYNTHESIS
Recent trace metal data from strata nearly three billion years old have been interpreted to indicate oxygenic photosynthesis (Planavsky et al. 2014, Crowe et al. 2013). The greatest control on the preservation of organic carbon in sediments is the time of exposure to oxygen (Hartnett et al. 1998). This has been observed in Lake Matano, where methanogenesis appears to be responsible for ~50% of organic carbon remineralization (Crowe et al. 2011).
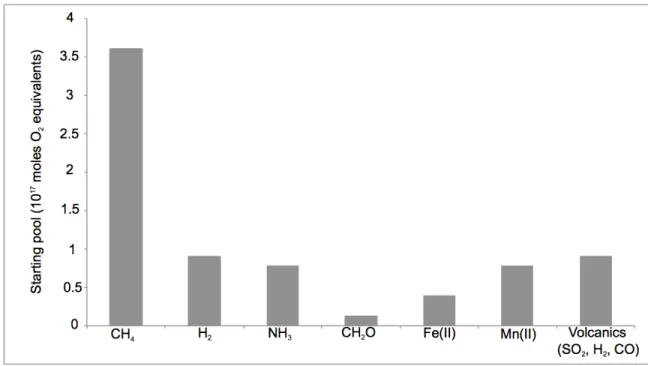
These represent upper limits for the concentrations of each of these species to satisfy the Faint Young Sun paradox (Pavlov et al. 2000). Based on Williams et al. 2012), mantle redox – and thus the Fe2+/Fe3+ ratio of new crustal material – is believed to have been constant since accretion. Estimates for methanogenic rates can also be made by analogy with Lake Matano (Crowe et al. 2008).
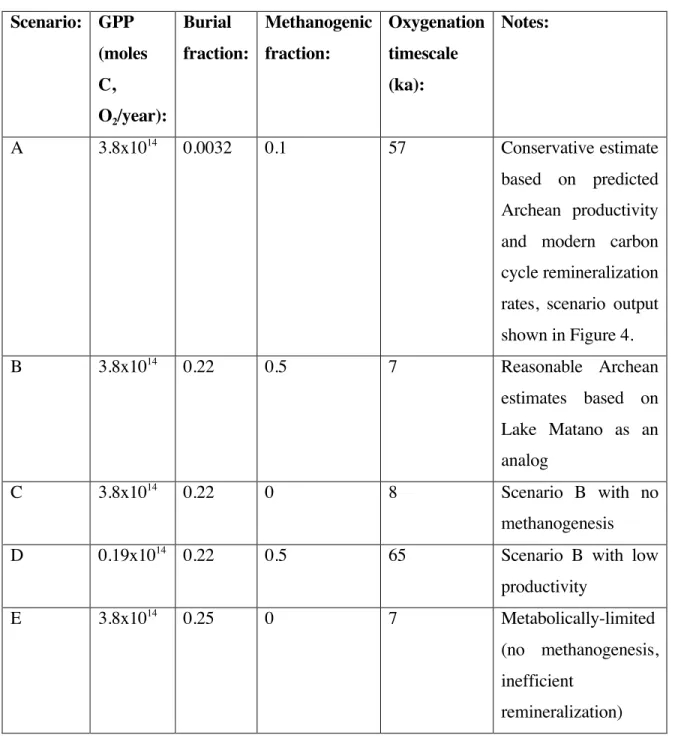
In Lake Matano, 50% or more of the fixed organic carbon has been shown to be remineralized via methanogenesis (Crowe et al. 2011). Dissolved inorganic carbon is not thought to be limiting for oxygen photosynthesis as DIC concentrations are thought to have been as high or higher than modern (Fischer et al. 2009). 2% in the Black Sea (Arthur et al. 1994), and 22% in Lake Matano (Crowe et al. 2011), and therefore this approach provides a very conservative estimate because this parameter exerts a strong control on the degree of oxygenation from the sea. oceans and atmosphere.
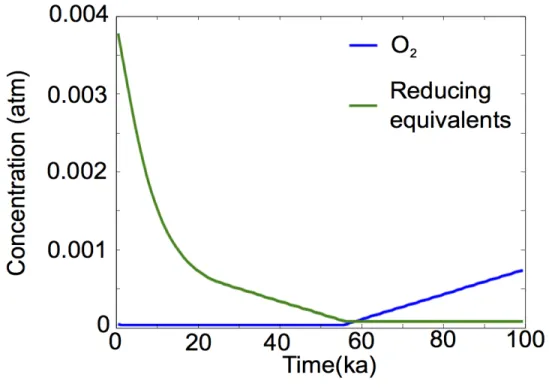
10%, well below that for Lake Matano, but comparable to that estimated for the pre-iltic photosynthetic Archean Earth system (Kharecha et al. 2005) results in oxygenation for about 57 kyr. Similar logic underlies a class of interpretations of the large C isotope excursions observed in carbonate rocks (δ13C values about 10 permil higher than Archean and Paleoproterozoic background values) associated with the oxygen rise (Martin et al. 2013). The aforementioned interpretations of 13C-enriched isotope ratio data in many Paleoproterozoic carbonates imply a greater relative flux of sedimentary organic matter associated with the oxygen increase (Karhu and Holland 1996, Rasmussen et al. 2013)—.
This is consistent with other proxies for a transition in the oxygenation state of the atmosphere at this time, such as the disappearance of redox-sensitive detrital grains (Johnson et al. 2014), the behavior of iron in paleosols and red beds (Holland 1984 ). , and the occurrence of evaporite sulfate mineral deposits (Chandler 1988). Data plotted are for Scenario 1 (summarized in Table 1), a conservative estimate of oxygenation time based on best estimates for Archean NPP (10% of modern marine values based on phosphorus limitation, per Bjerrum and Canfield, 2002), Archean methanogenic rates (0.1 of NPP, per Kharecha et al. 2005), and modern burial rates (0.32% of NPP, per Berner 1982).

A COMPLEX HISTORY OF PHOTOTROPHY REVEALED BY NOVEL CHLOROFLEXI LINEAGES
Although it has been suggested that photosynthesis was present in the last common ancestor of bacteria (Woese et al. 1985, Woese 1987), followed by widespread loss in most lineages, this idea remains controversial. While the structure of the bacterial tree of life is debated (e.g., Woese 1987, Williams et al. 2013, McInerney et al. 2014), intra-phylum organismal relationships are generally robust despite uncertainty in relationships between phyla (Pace 2009). Phylogenetic analysis of Chloroflexis phototrophy genes has suggested that anoxygenic photosynthesis in this group predates the evolution of oxygenic photosynthesis in cyanobacteria (Xiong et al. 2000), suggesting that this group is ancient and therefore a good candidate to investigate questions about early Earth history.
Recent culture- and sequence-based efforts have expanded the known taxonomic and metabolic diversity of Chloroflexi (e.g., the class Ardenticatenia capable of nitrate and iron oxide reduction, Hemp et al. Shikinejima is part of the Izu Islands, a chain of volcanic islands that formed in during the last 2-3 million years along the northern margin of the Izu-Bonin-Mariana arc (Kaneoka et al. 1970). Organ phylogenies of Chloroflexi, including the new phototrophs and other genomes described here, were constructed using the RpoB protein sequence (Fig. 2 ).
The incongruence between organismal and gene trees for the new phototrophic Chloroflexi described here suggests that photosynthesis genes were not vertically inherited from the last common ancestor of the phototrophic Chloroflexi. The genome bins for CP2_42A and JP3_7 restore most, but not all, of the bacteriochlorophyll synthesis pathway expected for phototrophic Chloroflexi. Although this may be a consequence of the incomplete nature of these genomes, the same bacteriochlorophyll synthesis gene complement has been described in the "Anaerolineae-like" phototrophic genome bin recovered from a Yellowstone National Park metagenome (Klatt et al. 2011).
Our group has previously sequenced several representatives of the Chloroflexi, filling gaps in the tree (Ward et al.). A history of horizontal gene transfer of phototrophy within the Chloroflexi is consistent with that of the Proteobacteria, which exhibit extensive intra-phylum HGT (Igarashi et al. 2001, Nagashima & Nagashima 2013. A single clear case of interphylum HGT is also recorded in the presence of a Proteobacteria-derived RC2 in a member of the Gemmatimonadetes (Zeng et al. 2014).
Kouleothrix aurantiaca, a member of the group previously known as 'Eikenboom morphotype 1851' (Seviour and Burkall 1999), was isolated from activated sludge in an industrial wastewater treatment plant (Kohno et al. 2002). It is closely related to members of the genus Roseiflexus (Beer et al. 2002), but phototrophy has not been observed in these organisms. The bacteriochlorophyll synthesis genes present in JP3_7 and CP2_42A may record a hybrid history of the pathway.
Draft genomes of two members of the Armatimonadetes group recovered from Japanese hot spring metagenomes.

MICROBIAL DIVERSITY AND IRON OXIDATION AT OKUOKU- HACHIKUROU ONSEN, A JAPANESE HOT SPRING ANALOG OF
PRECAMBRIAN IRON FORMATION
In Lake Matano, iron oxide production is thought to be largely driven by photoferrotrophy (Crowe et al. 2008), while at Chocolate Pots Hot Spring, iron oxidation is driven primarily by the abiotic reaction of iron with O2. Early crystalline iron oxide phases at OHK consist of ferrihydrite (Takashima et al. 2011), although this can age into more ordered iron oxide minerals such as hematite, goethite or lepidocrosite in travertines. Then, the most abundant sequence was selected as representative of each de novo OTU (Wang et al., 2007).
16S sequences were aligned using MAFFT (Katoh et al. 2002) and a phylogeny constructed using FastTree (Price et al. 2010). Iron-oxidizing Zetaproteobacteria are more commonly found in marine environments, particularly in deep ocean basins associated with hydrothermal iron sources (Emerson et al. 2010). Despite a similar physiology, Mariprofundaceae are not closely related to Gallionellaceae or other aerobic iron oxidizers, but instead form a distinct class within Proteobacteria (Emerson et al. 2007).
While iron oxide coats are typically associated with the betaproteobacterial iron-oxidizing genus Leptothrix (Emerson et al. 2010), they can also be produced by various iron oxidizers, including some strains of Zetaproteobacteria (Fleming et al. 2013). The abundance of amorphous iron oxide particles also supports the assignment of Gallionellaceae OTUs at OHK to Sideroxydans rather than stem-forming Gallionella (Emerson et al. 2010). It has been shown that in iron-rich systems where cyanobacteria are abundant and productive, only ~1% of released O2 oxidizes ferrous iron, while the rest escapes to the atmosphere (Rantamäki et al. 2016).
This includes anoxygenic iron-oxidizing phototrophs such as Chlorobium ferrooxidans (Heising et al. 1999), which use a metabolism thought to be important for archaeal BIF deposition (e.g., Kappler et al. 2005). Anaerolineae have generally been isolated as obligately anaerobic heterotrophs (e.g., Sekiguchi et al. 2003, Yamada et al. 2006), but the genom. Additionally, the genome for an organism closely related to Anaerolineae with photosynthesis genes was assembled from the Yellowstone National Park metagenome (Klatt et al. 2011).
For example, in Lake Matano, iron oxidation is thought to be driven primarily by photoferrotrophs (Crowe et al. 2008). It has been suggested that high iron concentrations are toxic to cyanobacteria, and that this may have played a role in slowing the oxygenation of the Archean atmosphere (Swanner et al. 2015). However, measured yields of aerobic iron oxidizers are typically much lower, on the order of 1 mol of CO2 fixed for every 40 mol of Fe(II) oxidized (Neubauer et al. 2002).
The lack of significant iron reduction in OHK is consistent with the predominance of ferric iron minerals in OHK deposits (Takashima et al. 2011) (Figure 3).