95 a seloniko a wangawangan ti immuna a postoral a somite {PMsd), ken dagiti prostomial a diding ket mabalin a mabalin a nalinya babaen ti coelomic a peritoneum (kitaen ti Meyer, 1901; Sokolow, 1911). Dagiti immuna a pudno a postoral a coelomic a supot ti Onychophora ket dagitoy dagiti panga (/); kalpasanna maipapan dayta kadagiti oral papillae (OR).

ORIGIN OF THE GONADS AND THE GONODUCTS
If the generation of arthropod somites is invariably teloblastic, it seems to follow that the numerical order of a segment in the body series of any arthropod will determine the homology of this segment with a segment of the same number in any other arthropod. With the polychaete, the first body somites form directly in the hyposphere of the trochophore; the crustacean nauplius larva has a short body region containing two primary somites (second antennal and mandibular), the continuation of secondary teloblastic somites beginning with the first maxilla segment.
8 SMITHSONIAN MISCELLANEOUS COLLECTIONS 95 in one or several somites near the posterior end of the body (fig. 2 A,
In the arthropods, direct development of the gonads and gonoducts from coelomic sacs is in ways similar to the development of these organs in the Onychophora described by Heymons (1901) in Scolopendra. The dorsal parts of the coelomic sacs of the millipede embryo consist of a double series of closed chambers, which unite on each side to form two long tubes with segmental compartments, the epithelial walls of which contain the germ.

14 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 EVOLUTION OF THE GENITAL EXIT APPARATUS
One of the first steps in the development of ectodermal genital appendages may simply consist of the formation of an external papapilla that carries the opening of each duct (fig.4B, Pin). In female insects, the opening of the oz>i- ductits communis (M, Ode), which is the true gonopore (Gpr), is usually hidden in a large genital chamber (GC), the exposed external opening, or vulva, of which is the gonotrem (Gtr).

20 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95
ONYCHOPHORA
The testicles of Onychophora are distributed in the posterior part of the body; they are tubular (Fig. 5C,D, Tes), but vary in length in different species. The spermatozoa are enclosed in spermatophores formed in the upper part of the ejaculatory duct (D, Sphr).

SMITHSONIAN MISCELLANEOUS COLLECTIONS 95 different species from small podlike capsules (E) to long- tubes or
PYCNOGONIDA (PANTOPODA)
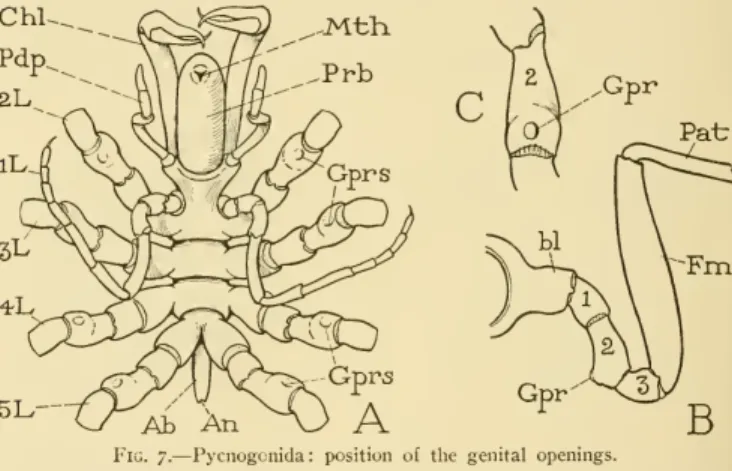
The body of a pycnogonid (fig. 7A) consists mainly of the appendage-bearing region composed of seven or eight segments, some of which are united; anteriorly, however, there is a large proboscis (Prh) with the mouth at its end, and posteriorly a rudimentary, unsegmented abdomen (Ab) bearing the anus. The genital openings of the pycnogonids in each sex are located ventrally on the second segments of the legs (fig. 4 A, Gprs, B, C, Gpr).
24 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 had metamerically repeated gonads with individual segmental outlets
XIPHOSURIDA
The dividing line between the two body parts in Xiphosura appears at first glance to be between segments VII and VIII, as the hilaria apparently belong to the prosoma and operculum, which is associated with the opisthosoma gill plates. Closer examination, however, shows that the first appendages actually borne by theopisthosoma are the first pair of gill-bearing plates, and that the operculum hangs from the amembranous region of the ventertus, which is united laterally with the posterior angles of the prosomal carapace.
SMITHSOXJAX MISCEIXANEOUS COLLECTIONS VOL. 95 responds with that in species of Pycnogonida having eight pairs of
The origin of the female is essentially like that of the male, opening a pair of similar papillae. The genital opening of Xiphosurida is therefore located, not on the base of the appendages, but on the outside of the eighth segment, as in Arachnida.
28 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 The xiphosurids copulate during the breeding season, but there is
EURYPTERIDA (GIGANTOSTRACA)
14 INSECT ABDOMEN SNODGRASS 29 preabclomen, because the extra segment of the scorpions appears to be located in this part of the body. Furthermore, the last gill plates of Xiphosura and the last lung books of the scorpion are located on segment XIII, while the last gills of the eurypterids are on.
30 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 are more closely related to the Arachnida than to the Xiphosurida
ARACHNIDA
In arachnid ovaries, the eggs develop in a large number of small follicular diverticula of the ovarian tubes, a feature that the arachnid share with the Xiphosurida and Onychophora. Live scorpion embryos develop either in the swellings of the ovarian tubes between the egg follicles or within them. The gonoducts, regardless of the shape of the gonads, are always a pair of tubes, usually extending forward from the ovaries or testes.
34 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 mating the male chelonethid attaches to the supporting surface be-
CRUSTACEA
The genital openings of females are always located on the sixth thoracic segment (somite X), and of males on the eighth (somite XII). In some cases, however, the spermatophores are only attached to the lower surface of the female's body. Spermatophores are probably taken from the gonopores of the male into the cavities of the first gonapophysis, and are pushed out of the latter by the second gonapophysis.

46 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 formed as invaginations of the body wall at the gonopores on the
MYRIAPODA PROGONEATA
There can be no doubt about the identity of the first body segment in Symphyla, since, although reduced, it bears a pair of legs. The first three body segments after the neck segment have only one pair of legs each, and the genital openings are always on the segment of the second pair. In most of the diplopods (Proterandria), the eighth or eighth and ninth pairs of legs (those of the third "double segment") are modified in structure to serve as intromitting organs and are converted into gonopods.
50 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 The gnathochilarium has been variously regarded as representing
CHILOPODA (MYRIAPODA OPISTHOGONEATA) The Chilopoda have a single median genital aperture, which in
The number of segments anterior to the genital segment is, however, so variable in the chilopods as a whole that no fixed numerical indication can be given to the genital segment itself. Among the Epimorpha, in which in most forms the definite segmentation is complete at hatching, the number of segments is highly variable in the Geophilomorpha, even in the same species, and may be very large.
54 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 while in Scolopcndroniorpha there are either 26 or 28 l)ody segments,
HEXAPODA
In insects other than Collembola and Protura, the primitive paired gonopores were probably on somite XIV (seventh abdominal segment) in the female and on somite XVII (tenth abdominal) in the male. Considering the fact that the formation of the somite in arthropods is teloblastic, and that the generative zone lies just before the terminal segment, the ortelson, it is clearly impossible that the genital segment could be the same somite in Collembola, Protura, and other insects. With the establishment of the genital canals in Collembola and Protura, the formation of somites has ceased, but the genital segment is present in the former.
58 SMITHSONIAN MISCELLANEOUS COLLECTIONS 95 insects opened originally on segment XVII, which is the tenth abdomi-
However, the subsequent union of the vasa deferentia with the median ejaculatory duct (B) necessitated a median and forward migration of the posterior ends of the lateral ducts. The correlated forward retraction of the posterior part of the ventral nerve cord then pulled the large nerves of the cerci (CerNv), released from the terminal ganglion, over the inwardly curved endings of the vasa deferentia (Vd). By the way, the relationship between the nerve pathways in the theme also shows that the cerci are appendages of the eleventh abdominal segment.
60 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95
14 INSECT BELLY SNODGRAS 61 The male bursa genitalis, or genital chamber, which usually contains the phallic organ or organs, projects into the ninth abdominal segment (fig. 22A, GC) since an invagination of the integument behind the ninth sternum, although its dorsal wall presumably belongs to the chest wall. the tenth segment. AcGId, accessory genital gland; Acd, aedeagus; Ccr, circus; Dej, ductus ejaculatorius; Enph, endophallus: Eppt, epiproct; GC, genital chamber or bursagenitalis; Gpr,gonopory (opening of ejaculatory duct);Gtr,gonotreme (opening of genital chamber);IXS, sternum of intraabdominal segment (male subgenital plate, hypandrium); Pap^t,paraproct; Phb,phallobase;Phi, phallus (median penis); Phtr, phallotreme (distal opening of endophallus); Pmr, paramere; Tcs, testis; Til, titillator, f'J, vas deferens; Virg, virga; VII-XI, seventh to eleven dominal segments; I'lINv,IXNv,XNv, XINv,main lateral nerves of eighth, ninth, tenth, eleventh abdominal segments emerging from the compound last ganglion of ventral nerves. These genital valves (valves, harps, harpagons) are always distinguishable from other similar but immovable or merely flexible lobes or processes born on the ninth or other segments of the genital complex by the fact that they are independently movable by muscles attached to their bases has been installed.

62 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 the basal segments (coxopodites) of the gonopods of pterygote insects
DEFINITIONS OF EXTERNAL GENITAL STRUCTURES OF THE MALE The principal parts of the external genitalia and their commonly
Gonopore (Gpr).— The external opening of the agenital canal, either exposed or concealed in a secondary invagination of the invagination, such as the genital chamber or an endophallic cavity. Gonotreme (Gtr).— The external opening of the genital bursa, orgenital chamber, in both sexes; the woman's vuiva. Phallobase (Phb). – A dilated basal part of the phallus, variable and often indistinguishable from the aedeagus.
COLLEMBOLA
Virga (Vir).— An aterminal phallic spine or slender rod, usually arising from the wall of the endophallus, and therefore protractable by the resistance of the latter. The unusual mating habits of the Sminthuridae, however, have long been known and have recently been described in detail by Falkenhan (1932) and Strebel (1932). Egg laying usually takes place about 14 to 18 days after mating, although the time varies depending on the temperature and age of the female.

70 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 The external genitalia of the Protura consist of a large bifid organ
DIPLURA AND THVSANURA
The coxal plates are also present on the genital segments (F,G), but the sternal plate is absent on the ninth segment of the male (G) and on both genital segments of the female. The gonapophyses of the ninth segment in male Thysanura are closely associated with the penis (G, sGon) and are often called parameres because they are said to correspond to additional genital structures called parameres in other male insects (see Heymons, 1897). With the subsequent reduction in size of the tenth segment during embryonic development, the ampullae of the male are transposed to the tenth segment and attached to the ectoderm at the posterior margin of this segment.
EPHEMEROPTERA
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 The insects are thus prepared to complete the reproductive function
It is suggestive, then, that the stylus-bearing plate, or stylist, is the product of jointed coxopods. The basal part of the stylus can in some cases be confused with the coxopoditis, or vice versa (A, I, J), and only with an examination of the musculature can two parts of the appendage be identified with certainty: the muscles of the letter always take their origin from the coxopodite, whether muscle-containing part of the laterisafreelobe (I,J) or part of the styliger plate (A,H). The pinnae differ in shape and relative size in different species (fig. 25B-H,J, K, Pen) and are often provided with accessory processes (C, D).

78 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 extension of the paraprocts between the lower edges of the tenth
DERMAPTERA
95 the base of the penis from a ductus conjunctus (Dejcn) proceeding from a seminal reservoir (rs) into which the vasareferentia discharge from a common duct (Vdcn) after first being enlarged as a pair of seminal vesicles (see Jordan, 1909, Heymons , 1912). In fact, they are the structures that form the basis of most of the elaboration of the external genital mechanism. Information on the exact function of the phallic organs and accessory genital processes in the order is far from complete.

88 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 95 of the ninth sternum may form but a rounded Hp beneath the geni-
The organ usually projects dorsally or forwards from the end of the epiproct, and may be longitudinally grooved on its back. It was first described by Hagenasa as "penis" in the sense that it is an organ to guide the sperm from the genital opening of the male to that of the female, a function that Hagen claimed was proven by observation, and the organ has since been commonly known as the "sperm conveyor". However, in Nemouravallicularia, Wu (1923) notes that the supraanal process merely serves to support the ninth segment of the female during mating, and Needham and Claassen (1925) describe the organ of Perlodes signatus as a guide for "the large soft penis emerging from the tip of the ninth segment. ." Hagen and Smith apparently did not observe the perpetual penile inPteronarcys, which was shown by Prison (1935) to be present and protracted in this genus.
ORTHOPTERA
95and provided with an extensive muscular system, while the ventral phallolo- and provided with an extensive muscular system, while the ventral phalloluere takes the form of a broad lobe with the gonopore at the base. The principal modifications of the phallus in the Tettigoniidae and Gryllidae are adaptations to the formation of spermatophores and the introduction of the latter into the genital chamber or sperm, the calories of the female; the phallic. The male sexual organs of Orthoptera will be the subject of a separate article to follow, in which the structure and function of the male organs in orthopteroid families other than Acrididae will be discussed.
In Tettigoniidae and Gryllidae a phallic organ has evolved in various ways, including ectophallic and endophalic parts, but in any case there is no close resemblance to the sexual structures of Blattidae, and there is little in the ontogeny of the organ to indicate that the adult structures have changed. evolved from every type of primitive structure common to the Orthoptera. The complex phallic organ of Acrididae is perhaps an extreme development of the tettigoniid-type structure, in which a large endophallic cavity becomes an important part of the phallic apparatus. Entwicklungsgeschichte von Peripatus Edwardsii Blanch, und Peripatus torquatusn.sp.. 1896. t)ber de Geschlechtsteilder Plecopteren, niitbesonderer View on the morphology of the genitalia.