MORPHOLOGY OF THE GONAD AND GENITAL TACTUS The embryonic genital ducts of insects are simple mesodermal tubes that are continuous with the mesodermal walls of the gonads. In most other insects in which the origin of reproductive organs has been studied, the gonads have been found to be formed as mesodermal ridges that contain germ cells from the beginning.

14 SMITHSONIAN MISCELLANEOUS COLLECTIONS 89 of the genital ridges each gonad in the female comes to be suspended
THE OVIPOSITOR OF THYSANURA
Thethysanuran ovipositor consists of four processes arising from the styloid lobes of the eighth and ninth abdominal segments. That the stylus-bearing plates of the Machilidae are the bases of the abdominal appendages, which are taken for granted, and are.
28 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 89 basis for all subsequent discussions in this paper on the morphology
GENERAL STRUCTURE OF THE PTERYGOTE OVIPOSITOR AND ASSOCIATED ORGANS
There seems little doubt that the pterygote ovipositoris is an organ formed from the appendages of the eighth and ninth abdominal segments, and that it is therefore homologous with the ovipositor of the Thysanura. Associated with the base of the ovipositor are typically three internal organs formed by median invaginations of the ectoderm.
THE GENITAL INVAGINATIONS
We may therefore assume that the first of the three median genital invaginations took place primitively on the posterior part of the seventh venter, and that, by union with the mesodermal oviducts (fig. 8 A, Odl), it became the primary median outlet tube of the ovaries , or oviductiiscommunis { Ode'). This second mid-length vaginal vagina is formed on the posterior part of the vent of the eighth abdominal segment (fig.8 A, Spt), and gives rise to a pouch for receiving the spermatozoa.
34 SMITHSONIAN MISCELLANEOUS
The oviduct of all Lepidoptera opens at the end of the ninth abdominal segment, or on a terminal segment, which is evidently the united ninth and tenth somite. It is therefore somewhat surprising to find that in the Lepidoptera the more primitive, anterior opening has been suppressed in the more generalized families, but the explanation lies in the fact that the posterior position of the oviporousis correlated with the tubular elongation of the abdomento.
THE GENITAL CHAMBER AND THE VESTIBULUM
With the transposition of the genital opening to the ninth segment, the ovipositor is in all cases suppressed (except in Cicadidae which have two genital openings, in which the passage runs directly into the canal of the ovipositor; and it must also be noted that the female exit apparatus of the Homopteraeodinger not well understood). INSECT BELLY SNODGRASS 39 always takes its original origin on the eighth gum (fig.9); the dorsal muscles of the second valvifers arise on the ninth tergum.
42 SMITHSONIAN MISCELLANEOUS 89
The second valvifers belong to the ninth abdominal segment, and are provided with muscles arising on the ninth tergum. They are
In some of the lower insects the third valvulae bear small terminal styli, which are usually lost in the adult stage. First valvulae (iVl). – The gonaphyses of the first gonopods, which form the ventral leaves of the ovipositor.
THE OVIPOSITOR OF ORTHOPTERA
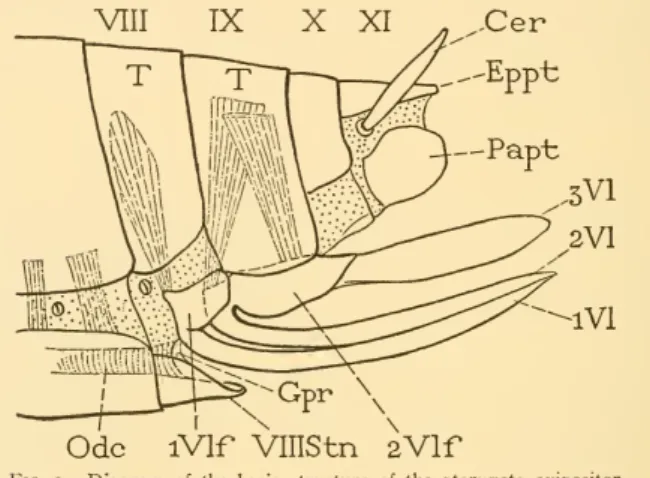
In Gryllidae and Acrididae, the inner or other flaps are rudimentary, and the functional part of the egg thus consists of only two pairs of flaps. The spiracles lie in the laterodorsal or paratergal region of the membrane {patg) above the dorso-pleural line. Each of the first, or ventral, valve flaps (iVl) is attached to a small, oval lateral plate on the base of the oviduct (iVlf), which is the first valve flap ("pileolus" of Chopard, 1920).
The upper end of the valvifer is connected, also by a flexible joint (&), to the basal part of the broad third or dorsal valve (sVl), the side of which overlaps ventrally. 10 C, 31B), in which each basal element of the arm of the ninth segment is divided into a valve (2Vlf) and a distal lobe (3VI).

50 SMITHSONIAN MISCELLANEOUS COLLECTIONS
Their thickened bases are united with each other medially (B,2VI) and laterally with the inner surfaces of the basal valve areas {sVlf) of the third valvulae. This Y-shaped dorsal sclerotization in the base of the inner valvulaeWalker (1919) calls the pons valvidarum (pnvl). The musculature of the ovipositor in Tettigoniidae and Gryllidae is more complex than in Hemiptera and Hymenoptera.
The ovipositor muscle and the terminal segments of the abdomen will be fully discussed in the description of Grylliisassiniilis. The oviduct of Gryllohlatta is, judging from the descriptions of Crampton and Walker (1919), very obvious.
52 SMITHSONIAN MISCELLANEOUS COLLECTIONS
The first (^) is attached ventrally to the pleural margin of the same segment; the second (j) is intersegmental because it is attached to the pleural margin of the next segment. The shortened eighth tergum (B, VIII) ends below a point separated by the dorso-pleural fold from the small eighth sternum (VlllStn), which projects like a small subgenital plate below the base of the ovipositor. The remaining lateral muscles in both genital segments are highly developed as muscles of the ovipositor (Fig. 17E).
The first, or anterior intervalvula (Fig. 17H, aiv), is a transverse bar between the anterior processes (g) of the secondvalvifers. The muscles of the ovipositor.—The musculature of the cricket's ovipositor is highly developed, but is not very complex.
VIII T IXT XT
It includes the ventral muscles of the seventh abdominal segment and the principal lateral and ventral muscles of the eighth and ninth segments. Ventrolateral dilators of the rectum.—A group of slender, branched fibers arising on the inside of each first valvifer behind the articulation with the ninth tergum; inserted ventrolaterally on the rectum.

62 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 89
The opposite movement of the first valvifer (indicated by the curved arrow on the left) reverses the movement of the valvulae. The distal end of the ovipositor is somewhat enlarged and forms a lancehead-like termination of the shaft with the acute tip slightly depressed (Fig.17B). There are no muscles in the terminal parts of the ovipositor; the opening of the distal extremities of leaves.
The venter of the tenth segment is mostly membranous, but it contains a narrow, transversely elongated sternal sclerotization (fig. 22C, XStn). 8 INSECT BELLY SNODGRASS 69 behind and below the bases of the cerci and the lower ends of the lateral plates of the tenth tergum (fig. i8).

BLATTA ORIENTALIS LINNAEUS
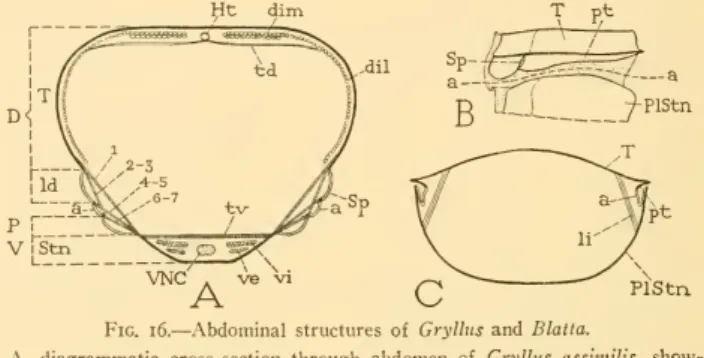
5 INSECT ABDOMEN SNODGRASS 7I 21, 22.– Two lateral muscles on each side, arising together on the anterior lateral part of the midplate of epiproct; diverging downwards. 2]^.—Origin anterior on inner side of paraproct; insertion posterior to the circumanal membrane above the posterior end of paraproct. However, the genital chamber of the cockroach appears to be a sac in the anterior wall of the great vestibulum (Vst) overlying the seventh sternum (VIIS) and the terminal lobes of the latter (VIISL).
The accessory glands (AcGl) open into the dorsal wall of the vestibular chamber in the region of the ninth venter.
VIIIT KT
THE OVIPOSITOR OF HEMIPTERA
The shaft of the ovipositor consists of the first and second valves, the first being external and ventral, and the second internal and dorsal. The first valve vifers have a pleural position below the gingiva on the sides of the eighth segment, although their posterior corners may be flexibly attached to the ninth gingiva. This basal sclerite (m) of the inner ramus of the first valve can be considered part of the valve.
Basally, as we have already noticed, the first valve is externally connected to the lower corner of the first valve (C, c). The ventral parts of the ninth segment are hidden, and the proctiger that Tower mistakes for the ninth segment is numerically the tenth segment, although anatomically it may represent segments X and XI.

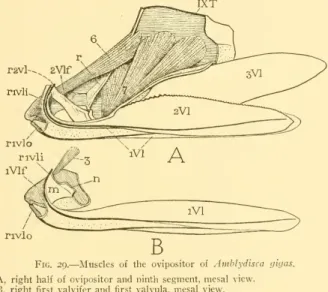
AMBLYDISCA GIGAS FOWLER
The sternal plates of the first two abdominal segments are narrow and are hidden in a ventral depression at the base of the abdomen. The distal part of the abdomen (fig. 28A) is constricted, and ends with the broad ends of the third valve (3VI) which usually encloses the end of the ovipositor (Ovp). The seventh segment of the abdomen is the first to be modified in relation to the sexual functions.
The lower anterior corner of the paratergite is connected to the dorsal posterior corner of the first valve valve (B, iVlf), which is normally hidden within the lower part of the seventh segment by the invagination of the lower parts of the eighth and ninth segments within the seventh (A). The intersegmental membrane (B, Mb) between the seventh and eighth segments is reflected from the dorsal (anterior) margin of the second valvifer.
SMITHSONIAN MISCELLANEOUS VOL
The first is an anterior, median, triangular plate (IS) with its lateral angles extended as a pair of arms (c) fused with the postcoxal bridges (Pcx), but each ending in a lateral extension (d), to which the tergosternal muscles attach is of the first abdominal segment. The sternum of this contra-abdominal segment, like that of the first segment, consists of two parts, one part exposed, the other hidden in the trunk cavity. The anterior portion of the second sternum is a weaker median sclerite (C, f) turned upwards towards the anterior margin of the posterior sclerite in the posterior wall of the tympanic cavity (B, /); its dorsal edge meets the posterior plate of the first sternum(s) in the transverse fold of the roof of the cavity.
The lateral angles of the anterior sclerite of the second sternum (C, /) are produced into thin bands (g) extending outwards to the lateral edges of the posterior clerite, each having a small expansion containing one of these condabdominal spiracles (IISp). The thetergumo of the seventh segment does not differ from the theterga preceding it. The seventh sternum (fig. 30 A, VIIS), however, is shortened, but has a deep edge at the lower end, the base of the ovipositor (Ovp).
96 SMITHSONIAN MISCELLANEOUS
THE OVIPOSITOR OF HYMENOPTERA
The first valves are completely separated from the other parts of the eighth segment and the formal intimate part of the basal mechanism of the ovary or bite. On the other hand, the terminal abdominal appendicular processes of adult Mecoptera and Dipterama may be true. The shaft of the ovipositor and the third valve project from above the seventh sternum (fig. 36 B).
The first valvifer is a small plate (Fig. 36E, iVlf) that articulates between the anterior ventral angle of the ninth tergum (a) and the dorsal margin of these condvalvifers (&); it gives attachment to the ramus of the first valvula (ribl) at its upper edge (c). The anterior end of the second valvifer is expanded into a dorsal lobe (D), to which the ramus of the second valvula (r2vl) is attached.

MEGARHYSSA ATRATA (PABRICIUS)
The sting of the bee has been the subject of many anatomical studies and the structural details of the skeletal parts are well known. The general musculature of the abdomen of the bee has been described by Betts (1923) and by Morison (1927). Its front wall is formed by the ventri of the eighth segment and contains the genital opening (fig. 44 A, c).
These parts include the spiral bearing plates of the eighth tergum (Lsp), the lateral plates of the ninth tergum (IXT), the first second valvifera {iVlf, 2Vlf) and the valves, which form the frame and shaft of the spike (Stng). Students of beeanatomy have adopted special names for the parts of the stinging apparatus which are convenient for descriptive purposes.

124 SMITHSONIAN MISCELLANEOUS COLLECTIONS
The quadrangular plates, or lateral tergal sclerites of the ninth dorsum. Fig. 41, IXT), are the mounting plates of the plug-in mechanism. The shaft of the sting consists of the united second valves (fig. 41, 2VI), and of the slender first valves or lancelets (iVl). The lancets slide freely over the lateral ventral edges of the balloon and stylet (Fig. 42E); their lower edges contact each other.
The shaft of the sting is supported at its base on the rami of the valves (fig. 41, rivl, r2vl). The basal surface of the balloon slopes significantly forward before the thermotherapy connections (Fig. 42 C), and a membrane fold extends dorsally into the body cavity.
INSECT ABDOMEN — SNODGRASS 127
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 89 merely a movable apodeme contained in a membranous fold at the
Short, thick, conical muscle, which is created anteriorlionante-costa of the seventh sternum {VIIS) in the position of the common middle ventricles (fig. The relative changes in the position of the various parts of the stinging apparatus before and after the elongation of the axis are shown above The axis of revolution in the basal parts passes up the edges (/) of the quadrate plates (IXT), which overlap the spiracular plates (Lsp) of the eighth segment.
It is possible that the strong contraction of the seventh abdominal segment, which increases the broadanterior lobes of thesternum, can beat. It is curious that all students of the mechanism of bee stings, since KraepeHn (1873), have regarded the muscles of the fur as angular (Vor-stossmuskeln) of the axis.

138 SMITHSONIAN MISCELLANEOUS
The only muscle inserted into the bee on the first tappet (triangular plate), as already shown, is the slender muscle of the lower edge of the spiral plate of the eighth segment (fig. The second tappets (oblong plates) remain relatively stationary , but there is sometimes a vibration of 8 INSECT ABDOMEN SNODGRASS 141 Due to the double articulation of the first valve on the quadrangular plate and the elongated plate (fig.48A), and due to the difference in length between the two axes (ab and ac ) of the valve, a very small movement of the square valve may occur plate ( B, d ) produces a relative.
Fig. 48.-Diagrams of a honey bee sting pattern to show the mechanism of inhibition and drag of the helantics. When the sting tip of a living bee is examined under the microscope, and no palpable object is allowed to penetrate, only an ulcer (remaining, in the writer's experience) appears to move back and forth over the style and extend beyond this last.

The morphology and development of the genitalia and genital tubes of Homoptera and Zygoptera as shown in the life histories of Philaenus and Agrion. On the development of the ovipositor and the efferent genital ducts of Tenebrio molitor L. Coleoptera), with remarks on the comparison of the latter organs in the two sexes.