!
" #$%&###'((#
#" )
Semua upaya para ilmuwan pada awalnya merupakan impian para ahli filsafat yang mustahil bisa diciptakan pada saat itu. Sebagai contoh: ada satu anekdot yang diungkapkan oleh Ratu Christina dari Swedia pada saat menyampaikan pidato kematian seorang ahli filsafat, matematika dan ilmuwan
* +, - + tahun 1650, yang meninggal akibat cuaca buruk musim salju:
“Sangat disayangkan Descartes meninggal, kenapa dengan kepandaiannya dia tidak dapat memperpanjang hidupnya sendiri”. Kenapa anekdot itu disampaikan dan apakah mungkin masa hidup dapat diperpanjang?.
Rene Descartes merupakan salah seorang ilmuwan yang dianggap mempunyai impian'impian yang jauh kedepan dan mempunyai ambisi untuk ikut mengontrol proses kehidupan, ide'idenya dianggap kontroversial pada saat itu.
Dia beranggapan bahwa hidup ini merupakan bagian dari alam. Semua yang ada di alam dapat diteliti dengan suatu metode dan hasil penelitian itu nantinya menjadi hukum alam yang dapat diekspresikan ke dalam bentuk bahasa dan matematika. Pada saat itu pembagian kelompok'kelompok keilmuan belum banyak sehingga hanya dikatakan bahwa hasil penelitian diungkapkan dalam bentuk bahasa. Dia juga mengatakan bahwa manusia dapat mengontrol dan memanipulasi proses kehidupan untuk memperoleh keuntungan. Apa yang dikemukakan Descartes mendapat sambutan para ilmuwan lain. Proses penelitian pada saat itu tidak diekspos kepermukaan karena para ilmuwan bekerja secara individu. Tetapi dalam kurun waktu 20 tahun ke depan penelitian yang mengarah pada modifikasi proses kehidupan berkembang dengan pesat. Penemuan Hukum Mendel mulai mengawali perwujudan impian itu pada abad ke 20 dan sejak tahun 1930 teori ini digunakan seluruh dunia karena dianggap efisien dan sistimatik di bidang pemuliaan organisme untuk
tujuan'tujuan tertentu bagi kepentingan manusia (Christiansen dan Sandoe, 2000).
Keberhasilan pengembangan budidaya perikanan sangat ditentukan, antara lain: oleh pasok benih yang meliputi faktor kualitas dan kuantitas benih.
Kualitas benih dipengaruhi ole kualitas induk (faktor genetis) dan faktor lingkungan (kualitas air, makanan dan penyakit). Kualitas induk sebagai faktor kualitas genetik mempunyai pengaruh terhadap kualitas benih dalam pengertian meningkatkan jumlah benih, pertumbuhan yang cepat, dan ketahanan terhadap perubahan lingkungan (penyakit dan kualitas air).
Peningkatan kualitas induk memlaui peningkatan sifat'sifat genetis baik kualitatif maupun kuantitatif yang dapat dilakukan melelui seleksi sifat atau karakter dari calon'calon induk. Selain itu, pengelolaan induk untuk menghindari terjadinya silang dalam (in breeding) yang menyebabkan menurunnya daya waris (heritabiltas) induk tersebut.
Tujuan dari penulisan makalah ini untuk meninjau konsep genetika dan analisis pewarisan sifat'sifat kuantitatif atau genetika kuantitatif di bidang ilmu budidaya perikanan khusus untuk kepentingan pemuliaan ikan. Pada makalah ini akan dijelaskan aplikasi pemahaman mengenai falsafah sain khususnya mengenai Ontologi, Epistemologi dan Aksiologi. Secara ontologi akan membahas eksistensi (keberadaan) ilmu genetika dan esensi atau keberartiannya dalam bidang budidaya perikanan. Secara epistomologi akan membahas pendekatan' pendekatan teoritis dan metode sebagai pemikiran dasar pengembangan ilmu genetika kuantitatif. Sedangkan secara aksiologi akan membahas bagaimana memanfaatkan kaidah'kaidah atau analisis genetika kuantitatif untuk kepentingan pemuliaan dan heritabilitas dalam suatu kegiatan budidaya perikanan.
."
/ 0"
."#" + 1 * + 2 * *3 *3 4 * - *- - - 1
Genetika berasal dari bahasa Yunani γέννω atau genno yang berarti
"melahirkan". merupakan cabang biologi yang penting saat ini. Ilmu ini mempelajari berbagai aspek yang menyangkut pewarisan sifat dan variasi sifat pada organisme maupun suborganisme (seperti virus dan prion). Ada pula yang
dengan singkat mengatakan, genetika adalah ilmu tentang gen. Nama
"genetika" diperkenalkan oleh William Bateson pada suatu surat pribadi kepada Adam Chadwick dan ia menggunakannya pada Konferensi Internasional tentang genetika ke'3 pada tahun 1906. Bidang kajian genetika dimulai dari wilayah molekular hingga populasi. Secara lebih rinci, genetika berusaha menjelaskan:
material pembawa informasi untuk diwariskan (bahan genetik), bagaimana informasi itu diekspresikan (ekspresi genetik), bagaimana informasi itu dipindahkan dari satu individu ke individu yang lain (pewarisan genetik) dan mengapa terjadinya variasi antar individu dalam suatu populasi (Ragam genetik).
Genetika kuantitatif adalah cabang genetika yang membahas pewarisan sifat'sifat terukur (kuantitatif atau metrik), yang tidak bisa dijelaskan secara langsung melalui hukum pewarisan Mendel. Sifat'sifat yang tergolong sifat kuantitatif bidang perikanan misalnya panjang tubuh ikan, pertambahan berat, hasil panen, jumlah telur (fekunditas), kandungan biokimia, dan lain sebagainya.
Genetika kuantitatif menerapkan hukum pewarisan Mendel untuk gen dengan pengaruh yang kecil/lemah (minor gene). Selain itu, diasumsikan pula bahwa tidak hanya sedikit gen yang mengendalikan suatu sifat melainkan banyak gen. Karena itu, sifat kuantitatif sering dasamakan dengan sifat poligenik. Ilmu ini banyak menggunakan matematika dan statistika dalam menjelaskan prinsip'prinsip yang dipakai maupun dalam metodologinya. Namun demikian, penerapan ilmu ini dalam ilmu pemuliaan sangat bermanfaat dalam bidang pertanian dan perikanan.
Pewarisan genetik (genetic inheritance) adalah aspek pertama yang dipelajari orang dalam genetika karena berkaitan langsung dengan fenotipe.
Sebagai contoh, Gregor Johann Mendel mempelajari peawarisan tujuh sifat pada tanaman kapri, atau Karl Pearson (salah satu pelopor genetika kuantitatif) mempelajari pewarisan ukuran tubuh orang tua dan anaknya. Kajian genetika yang mempelajari aspek ini dijuluki genetika transmisional, karena membahas bagaimana bahan genetik dialihkan dari satu generasi ke geneasi lainnya beserta implikasi yang menyertainya.
Genetika kuantitatif dimulai pada tahun 1918 oleh Ronald Fisher (ahli biostatistika dari Inggris) yang menerbitkan On the correlation between relatives on the supposition of Mendelian inheritance (secara bebas berarti "Keterkaitan antar kerabat berdasarkan pewarisan Mendel"), yang mengakhiri perseteruan
antara teori biometri (Pearson dkk.) dan teori Mendel sekaligus mengawali sintesis keduanya.
."." 5 *3 * * -
Meskipun orang biasanya menetapkan genetika dimulai dengan ditemukannya kembali naskah artikel yang ditulis Gregor Mendel pada tahun 1900, sebetulnya genetika sebagai "ilmu pewarisan" atau 2 - + sudah dikenal sejak masa prasejarah, seperti domestikasi dan pengembangan berbagai ras ternak dan kultivar tanaman. Orang juga sudah mengenal efek persilangan dan perkawinan sekerabat serta membuat sejumlah prosedur dan peraturan mengenai hal tersebut sejak sebelum genetika berdiri sebagai ilmu yang mandiri. Silsilah tentang penyakit pada keluarga, misalnya, sudah dikaji orang sebelum itu. Namun demikian, pengetahuan praktis ini tidak memberikan penjelasan penyebab dari gejala'gejala itu.
Teori populer mengenai pewarisan yang dianut pada masa itu adalah teori pewarisan campur: seseorang mewariskan campuran rata dari sifat'sifat yang dibawa tetuanya, terutama dari pejantan karena membawa sperma. Hasil penelitian Mendel menunjukkan bahwa teori ini tidak berlaku karena sifat'sifat dibawa dalam kombinasi yang dibawa alel'alel khas, bukannya campuran rata.
Pendapat terkait lainnya adalah teori Lamarck: sifat yang diperoleh tetua dalam hidupnya diwariskan kepada anaknya. Teori ini juga patah dengan penjelasan Mendel bahwa sifat yang dibawa oleh gen tidak dipengaruhi pengalaman individu yang mewariskan sifat itu. Charles Darwin juga memberikan penjelasan dengan hipotesis pangenesis dan kemudian dimodifikasi oleh Francis Galton. Dalam pendapat ini, sel'sel tubuh menghasilkan partikel'partikel yang disebut gemmula yang akan dikumpulkan di organ reproduksi sebelum pembuahan terjadi. Jadi, setiap sel dalam tubuh memiliki sumbangan bagi sifat'sifat yang akan dibawa zuriat (keturunan).
Pada masa pra'Mendel, orang belum mengenal gen dan kromosom (meskipun DNA sudah diekstraksi namun pada abad ke'19 belum diketahui fungsinya). Saat itu orang masih beranggapan bahwa sifat diwariskan lewat sperma (tetua betina tidak menyumbang apa pun terhadap sifat anaknya).
Peletakan dasar ilmiah melalui percobaan sistematik baru dilakukan pada paruh akhir abad ke'19 oleh Gregor Johann Mendel. Ia adalah seorang biarawan dari Brno (Brünn dalam bahasa Jerman), Kekaisaran Austro'Hungaria (sekarang bagian dari Republik Ceko). Mendel disepakati umum sebagai 'pendiri genetika' setelah karyanya "Versuche über Pflanzenhybriden" atau Percobaan mengenai Persilangan Tanaman (dipublikasi cetak pada tahun 1866) ditemukan kembali secara terpisah oleh Hugo de Vries, Carl Correns, dan Erich von Tschermak pada tahun 1900. Dalam karyanya itu, Mendel pertama kali menemukan bahwa pewarisan sifat pada tanaman (ia menggunakan tujuh sifat pada tanaman kapri, Pisum sativum) mengikuti sejumlah nisbah matematika yang sederhana. Yang lebih penting, ia dapat menjelaskan bagaimana nisbah'nisbah ini terjadi, melalui apa yang dikenal sebagai 'Hukum Pewarisan Mendel'.
Dari karya ini, orang mulai mengenal konsep gen (Mendel menyebutnya 'faktor'). Gen adalah pembawa sifat. Alel adalah ekspresi alternatif dari gen dalam kaitan dengan suatu sifat. Setiap individu disomik selalu memiliki sepasang alel, yang berkaitan dengan suatu sifat yang khas, masing'masing berasal dari tetuanya. Status dari pasangan alel ini dinamakan genotipe. Apabila suatu individu memiliki pasangan alel sama, genotipe individu itu bergenotipe homozigot, apabila pasangannya berbeda, genotipe individu yang bersangkutan dalam keadaan heterozigot. Genotipe terkait dengan sifat yang teramati. Sifat yang terkait dengan suatu genotipe disebut fenotipe.
Setelah penemuan ulang karya Mendel, genetika berkembang sangat pesat. Perkembangan genetika sering kali menjadi contoh klasik mengenai penggunaan metode ilmiah dalam ilmu pengetahuan atau sains.
Berikut adalah tahapan'tahapan perkembangan genetika:
• 1859 Charles Darwin menerbitkan The Origin of Species, sebagai dasar variasi genetik.;
• 1865 Gregor Mendel menyerahkan naskah Percobaan mengenai Persilangan Tanaman;
• 1878 E. Strassburger memberikan penjelasan mengenai pembuahan berganda;
• 1900 Penemuan kembali hasil karya Mendel secara terpisah oleh Hugo de Vries (Belgia), Carl Correns (Jerman), dan Erich von Tschermak (Austro' Hungaria) ==> awal genetika klasik;
• 1903 Kromosom diketahui menjadi unit pewarisan genetik;
• 1905 Pakar biologi Inggris William Bateson mengkoinekan istilah 'genetika';
• 1908 dan 1909 Peletakan dasar teori genetika populasi oleh Weinberg (dokter dari Jerman) dan secara terpisah oleh James W. Hardy (ahli matematika Inggris) ==> awal genetika populasi;
• 1910 Thomas Hunt Morgan menunjukkan bahwa gen'gen berada pada kromosom, menggunakan lalat buah (Drosophila melanogaster) ==> awal sitogenetika;
• 1913 Alfred Sturtevant membuat peta genetik pertama dari suatu kromosom;
• 1918 Ronald Fisher (ahli biostatistika dari Inggris) menerbitkan On the correlation between relatives on the supposition of Mendelian inheritance (secara bebas berarti "Keterkaitan antarkerabat berdasarkan pewarisan Mendel"), yang mengakhiri perseteruan antara teori biometri (Pearson dkk.) dan teori Mendel sekaligus mengawali sintesis keduanya ==> awal genetika kuantitatif;
• 1927 Perubahan fisik pada gen disebut mutasi;
• 1928 Frederick Griffith menemukan suatu molekul pembawa sifat yang dapat dipindahkan antarbakteri (konjugasi);
• 1931 Pindah silang menyebabkan terjadinya rekombinasi;
• 1941 Edward Lawrie Tatum and George Wells Beadle menunjukkan bahwa gen'gen menyandi protein, ==> awal dogma pokok genetika;
• 1944 Oswald Theodore Avery, Colin McLeod and Maclyn McCarty mengisolasi DNA sebagai bahan genetik (mereka menyebutnya prinsip transformasi);
• 1950 Erwin Chargaff menunjukkan adanya aturan umum yang berlaku untuk empat nukleotida pada asam nukleat, misalnya adenin cenderung sama banyak dengan timin;
• 1950 Barbara McClintock menemukan transposon pada jagung;
• 1952 Hershey dan Chase membuktikan kalau informasi genetik bakteriofag (dan semua organisme lain) adalah DNA;
• 1953 Teka'teki struktur DNA dijawab oleh James D. Watson dan Francis Crick berupa pilin ganda (double helix), berdasarkan gambar'gambar difraksi sinar X DNA dari Rosalind Franklin ==> awal genetika molekular;
• 1956 Jo Hin Tjio dan Albert Levan memastikan bahwa kromosom manusia berjumlah 46;
• 1958 Eksperimen Meselson'Stahl menunjukkan bahwa DNA digandakan (direplikasi) secara semikonservatif;
• 1961 Kode genetik tersusun secara triplet;
• 1964 Howard Temin menunjukkan dengan virusRNA bahwa dogma pokok dari tidak selalu berlaku;
• 1970 Enzim restriksi ditemukan pada bakteri Haemophilus influenzae, memungkinan dilakukannya pemotongan dan penyambungan DNA oleh peneliti (lihat juga RFLP) ==> awal bioteknologi modern;
• 1977 Sekuensing DNA pertama kali oleh Fred Sanger, Walter Gilbert, dan Allan Maxam yang bekerja secara terpisah. Tim Sanger berhasil melakukan sekuensing seluruh genom Bakteriofag Φ'X174;, suatu virus
==> awal genomika;
• 1983 Perbanyakan (amplifikasi) DNA dapat dilakukan dengan mudah setelah Kary Banks Mullis menemukan Reaksi Berantai Polymerase (PCR);
• 1985 Alec Jeffreys menemukan teknik sidik jari genetik.
• 1989 Sekuensing pertama kali terhadap gen manusia pengkode protein CFTR penyebab cystic fibrosis;
• 1989 Peletakan landasan statistika yang kuat bagi analisis lokus sifat kuantitatif (analisis QTL) ;
• 1995 Sekuensing genom Haemophilus influenzae, yang menjadi sekuensing genom pertama terhadap organisme yang hidup bebas;
• 1996 Sekuensing pertama terhadap eukariota: khamir Saccharomyces cerevisiae;
• 1998 Hasil sekuensing pertama terhadap eukariota multiselular, nematoda Caenorhabditis elegans, diumumkan;
• 2001 Draf awal urutan genom manusia dirilis bersamaan dengan mulainya Human Genome Project;
• 2003 Proyek Genom Manusia (Human Genome Project) menyelesaikan 99% pekerjaannya pada tanggal (14 April) dengan akurasi 99.99%
."$" 6 5 *37, 5 *3 3 * -
Genetika berkembang baik sebagai ilmu murni maupun ilmu terapan.
Cabang'cabang ilmu ini terbentuk terutama sebagai akibat pendalaman terhadap suatu aspek tertentu dari objek kajiannya. Cabang'cabang murni genetika: genetika molekular, genetika sel (sitogenetika), genetika populasi, genetika kuantitatif dan genetika perkembangan. Sedangkan cabang'cabang terapan genetika:genetika kedokteran, ilmu pemuliaan, rekayasa genetika atau rekayasa gen.
Kajian genetika klasik dimulai dari gejala fenotipe (yang tampak oleh pengamatan manusia) lalu dicarikan penjelasan genotipiknya hingga ke aras gen. Berkembangnya teknik'teknik dalam genetika molekular secara cepat dan efisien memunculkan filosofi baru dalam metodologi genetika, dengan membalik arah kajian. Karena banyak gen yang sudah diidentifikasi sekuensnya, orang memasukkan atau mengubah suatu gen dalam kromosom lalu melihat implikasi fenotipik yang terjadi. Teknik'teknik analisis yang menggunakan filosofi ini dikelompokkan dalam kajian genetika arah'balik atau reverse genetics, sementara teknik kajian genetika klasik dijuluki genetika arah'maju atau forward genetics.
$"
/ 0"
$"#" 8*+ 4 + ) 2 - +
Meskipun orang biasanya menetapkan genetika dimulai dengan ditemukannya kembali naskah artikel yang ditulis Gregor Mendel pada tahun 1900, sebetulnya genetika sebagai "ilmu pewarisan" atau 2 - + sudah dikenal sejak masa prasejarah, seperti domestikasi dan pengembangan berbagai ras ternak dan kultivar tanaman. Orang juga sudah mengenal efek persilangan dan perkawinan sekerabat serta membuat sejumlah prosedur dan peraturan mengenai hal tersebut sejak sebelum genetika berdiri sebagai ilmu yang mandiri. Silsilah tentang penyakit pada keluarga, misalnya, sudah dikaji orang sebelum itu. Namun demikian, pengetahuan praktis ini tidak memberikan penjelasan penyebab dari gejala'gejala itu.
Teori populer mengenai pewarisan yang dianut pada masa itu adalah teori pewarisan campur: seseorang mewariskan campuran rata dari sifat'sifat yang
dibawa tetuanya, terutama dari pejantan karena membawa sperma. Hasil penelitian Mendel menunjukkan bahwa teori ini tidak berlaku karena sifat'sifat dibawa dalam kombinasi yang dibawa alel'alel khas, bukannya campuran rata.
Pendapat terkait lainnya adalah teori Lamarck: sifat yang diperoleh tetua dalam hidupnya diwariskan kepada anaknya. Teori ini juga patah dengan penjelasan Mendel bahwa sifat yang dibawa oleh gen tidak dipengaruhi pengalaman individu yang mewariskan sifat itu. Charles Darwin juga memberikan penjelasan dengan hipotesis pangenesis dan kemudian dimodifikasi oleh Francis Galton. Dalam pendapat ini, sel'sel tubuh menghasilkan partikel'partikel yang disebut gemmula yang akan dikumpulkan di organ reproduksi sebelum pembuahan terjadi. Jadi, setiap sel dalam tubuh memiliki sumbangan bagi sifat'sifat yang akan dibawa zuriat (keturunan).
Pada masa pra'Mendel, orang belum mengenal gen dan kromosom (meskipun DNA sudah diekstraksi namun pada abad ke'19 belum diketahui fungsinya). Saat itu orang masih beranggapan bahwa sifat diwariskan lewat sperma (tetua betina tidak menyumbang apa pun terhadap sifat anaknya).
$"." 8*+ 4 2 * 8 + *
* (dari bahasa Belanda: gen) adalah unit pewarisan sifat bagi organisme hidup. Bentuk fisiknya adalah urutan DNA yang menyandi suatu protein, polipeptida, atau seuntai RNA yang memiliki fungsi bagi organisme yang memilikinya. Batasan modern gen adalah suatu lokasi tertentu pada genom yang berhubungan dengan pewarisan sifat dan dapat dihubungkan dengan fungsi sebagai regulator (pengendali), sasaran transkripsi, atau peran+peran fungsional lainnya[1][2]. Penggunaan "gen" dalam percakapan sehari'hari (misalnya "gen cerdas" atau "gen warna rambut") sering kali dimaksudkan untuk alel: pilihan variasi yang tersedia oleh suatu gen. Meskipun ekspresi alel dapat serupa, orang lebih sering menggunakan istilah alel untuk ekspresi gen yang secara fenotipik berbeda. Gen diwariskan oleh satu individu kepada keturunannya melalui suatu proses reproduksi, bersama'sama dengan DNA yang membawanya. Dengan demikian, informasi yang menjaga keutuhan bentuk dan fungsi kehidupan suatu organisme dapat terjaga.
Pada saat itu DNA sudah ditemukan dan diketahui hanya berada pada kromosom (1869), tetapi orang belum menyadari bahwa DNA terkait dengan gen. Melalui penelitian Oswald Avery terhadap bakteri Pneumococcus (1943), serta Alfred Hershey dan Martha Chase (publikasi 1953) dengan virus bakteriofag T2, barulah orang mengetahui bahwa DNA adalah bahan genetik.
Pada tahun 1940an, George Beadle dan Edward Tatum mengadakan percobaan dengan Neurospora crassa. Dari percobaan tersebut, Beadle dan Tatum dapat menarik hipotesis bahwa gen mengkode enzim, dan mereka menyimpulkan bahwa satu gen menyintesis satu enzim (one gene+one enzyme theory). Beberapa puluh tahun kemudian, ditemukan bahwa gen mengkode protein yang tidak hanya berfungsi sebagai enzim saja, dan beberapa protein tersusun dari dua atau lebih polipeptida. Dengan adanya penemuan'penemuan tersebut, pendapat Beadle dan Tatum, one gene+one enzyme theory, telah dimodifikasi menjadi teori satu gen'satu polipeptida (one gene+one polypetide theory).
Pada sel eukariot, gen terdiri dari: domain regulasi inisiasi transkripsi, yang terdiri antara lain dari: deret GCCACACCC, ATGCAAAT, kotak GC, kotak CCAAT dan kotak TATA, intron dan ekson yang merupakan area kodikasi protein yang dapat ditranskripsi secara overlapping atau nonoverlapping. Sebagai contoh, pada kode dengan tiga deret nukleotida (kodon triplet) AUU GCU CAG,
dapat secara dibaca nonoverlapping sebagai AUU GCU CAG atau dibaca secara overlapping sebagai AUU UUG UGC GCU CUC CAG. Walaupun pada sekitar tahun 1961, telah diketahui bahwa asam amino dikodikasi oleh kodon secara nonoverlapping, telah ditemukan protein berbeda hasil transkripsi dengan pergeseran overlapping kodon, serta domain regulasi akhir transkripsi
Ekspresi gen adalah proses dimana kode'kode informasi yang ada pada gen diubah menjadi protein'protein yang beroperasi hanya di dalam sel. kspresi gen terdiri dari dua tahap:
1. Transkripsi, proses pembuatan salinan RNA.
2. Translasi, proses sintesis polipeptida yang spesifik di dalam ribosom.
Proses transkripsi DNA menjadi mRNA dan translasi mRNA menjadi sebuah polipeptida disebut dogma sentral (central dogma). Dogma sentral berlaku pada prokariot dan eukariot. Namun, pada eukariot ada tahap tambahan yang terjadi di antara transkripsi dan translasi yang disebut tahap pre'mRNA.
Tahap pre'mRNA adalah untuk menyeleksi mRNA yang akan dikirim keluar nukleus untuk ditranslasikan di ribosom. Ekson merupakan mRNA yang akan dikirim keluar nukleus untuk ditranslasikan, sedangkan intron merupakan mRNA yang akan tetap berada di dalam nukleus karena kemungkinan mRNA tersebut akan membentuk protein yang tidak fungsional (tidak berguna) jika ditranslasikan. Intron kemudian akan terurai kembali untuk membentuk rantai mRNA baru. Diketahui pula bahwa beberapa kesalahan yang disebut mutasi dapat terjadi pada proses ekspresi gen ini.
$"$" 8*+ 4 2 * 8 +
Dalam genetika, (dari bahasa Belanda, allel, dibentuk dari kata bahasa Yunani, αλλήλων atau allélon, "saling berhadapan") merupakan bentuk' bentuk alternatif dari gen pada suatu lokus. Alel terbentuk karena adanya variasi pada urutan basa nitrogen akibat peristiwa mutasi. Istilah ini muncul akibat penggunaan allelomorph oleh William Bateson pada buku karangannya Mendel's Principles of Heredity (1902).
Lokus dikatakan bersifat polimorfik apabila memiliki variasi alel dalam suatu populasi dan, sebaliknya, dikatakan bersifat monomorfik ("satu bentuk") apabila tidak memiliki variasi. Individu yang memiliki alel sama pada suatu lokus dikatakan memiliki genotipe yang homozigot sedangkan yang memiliki alel berbeda dikatakan heterozigot. Karena genotipe diekspresikan menjadi suatu fenotipe, alel dapat menyebabkan perbedaan penampilan di antara individu' individu dalam suatu populasi.
Konsep alel dapat sedikit berbeda tergantung dari apa yang dilihat. Dua atau lebih alel dikatakan "identik karena keadaan" (identical by state, IBS) apabila mereka memberikan efek fenotipe yang sama, meskipun ada kemungkinan memiliki sekuens basa yang berbeda. Dalam tingkat DNA, khususnya dalam penggunaan SNP, artinya lebih ketat karena mereka harus memiliki sekuens basa yang sama.
Apabila berada pada lokus yang sama pada dua kromosom yang homolog, sepasang alel dari satu individu dikatakan "identik karena segaris keturunan"
(identical by descent, IBD) jika dan hanya jika keduanya merupakan salinan sempurna dari satu alel milik satu individu pada generasi pendahulunya. Apabila terjadi mutasi terhadap salah satu salinan pada transmisi di antara generasi, alel mutan ini tidak dianggap identik lagi.
Identitas alel dipakai dalam genetika populasi untuk menentukan tingkat kedekatan kekerabatan antara dua individu yang memiliki moyang yang sama, seperti pada anggota generasi hasil perkawinan sekerabat.
$"'" 8*+ 4 2 * - 8 2 + *8- 4
*8- 4 (harafiah berarti "tipe gen") adalah istilah yang dipakai untuk menyatakan keadaan genetik dari suatu individu atau sekumpulan individu populasi. Genotipe dapat merujuk pada keadaan genetik suatu lokus maupun keseluruhan bahan genetik yang dibawa oleh kromosom (genom). Genotipe
dapat berupa homozigot atau heterozigot. Setelah orang dapat melakukan transfer gen, muncul pula penggunaan istilah hemizigot.
Dalam genetika Mendel (genetika klasik), genotipe sering dilambangkan dengan huruf yang berpasangan; misalnya AA, Aa, atau B1B1. Pasangan huruf yang sama menunjukkan bahwa individu yang dilambangkan adalah homozigot (AA dan B1B1), sedangkan pasangan huruf yang berbeda melambangkan individu heterozigot. Sepasang huruf menunjukkan bahwa individu yang dilambangkan ini adalah diploid (2n). Sebagai konsekuensi, individu tetraploid (4n) homozigot dilambangkan dengan AAAA, misalnya.
$"9" 8*+ 4 2 * 8 2 + *8- 4
*8- 4 adalah suatu karakteristik (baik struktural, biokimiawi, fisiologis, dan perilaku) yang dapat diamati dari suatu organisme yang diatur oleh genotipe dan lingkungan serta interaksi keduanya. Pengertian fenotipe mencakup berbagai tingkat dalam ekspresi gen dari suatu organisme. Pada tingkat organisme, fenotipe adalah sesuatu yang dapat dilihat/diamati/diukur, sesuatu sifat atau karakter. Dalam tingkatan ini, contoh fenotipe misalnya warna mata, berat badan, atau ketahanan terhadap suatu penyakit tertentu. Pada tingkat biokimiawi, fenotipe dapat berupa kandungan substansi kimiawi tertentu di dalam tubuh. Sebagai misal, kadar gula darah atau kandungan protein dalam beras. Pada taraf molekular, fenotipe dapat berupa jumlah RNA yang diproduksi atau terdeteksinya pita DNA atau RNA pada elektroforesis.
Fenotipe ditentukan sebagian oleh genotipe individu, sebagian oleh lingkungan tempat individu itu hidup, waktu, dan, pada sejumlah sifat, interaksi antara genotipe dan lingkungan. Pengamatan fenotipe dapat sederhana (masalnya warna bunga) atau sangat rumit hingga memerlukan alat dan metode khusus. Namun demikian, karena ekspresi genetik suatu genotipe bertahap dari tingkat molekular hingga tingkat individu, seringkali ditemukan keterkaitan antara sejumlah fenotipe dalam berbagai tingkatan yang berbeda'beda.
Perfomans atau penampilan (ekspresi) suatu karakter atau sifat atau ciri atau variasi suatu organisme memiliki pola keteraturan yang unik atau spesifik atau khusus. Dengan melihat, mengetahui, mengenali dan menganalisis pola keteraturan yang unik atau spesifik ini, maka pemuliaan, seleksi, manajemen dan konservasi organisme tersebut dapat dilakukan dengan lebih mudah, baik dan cepat serta efektif dan efisien.
Performans sifat/karakter/ciri fenotip dapat bersifat kualitatif dan kuantitatif. Sifat atau ciri kualitatif adalah ciri/sifat/karakter yang dikategorikan atau dideskripsikan, sedangkan sifat atau karakter kuantitatif adalah ciri yang diukur ketimbang dideskripsikan. Ciri'ciri ini tidak memisahkan individu'individu atas kategori fenotipe yang berbeda, seperti warna hitam atau albino. Malah, setiap karakter atau sifat kuantitatif memperlihatkan suatu sebaran atau distribusi yang kontinyu dalam suatu populasi. Perbedaan fenotipe antar individu adalah masalah tingkatan atau perbedaan derajat ketimbang macamnya.
Fenotipe kuantitatif adalah performans ciri/karakter/traits produksi dan pertumbuhan organisme yang penting seperti: panjang, berat, kelulusan hidup, kandungan biokimia, rasio RNA/DNA, kesuburan reproduksi, biomassa, rasio tulang dan daging serta jumlah/kualitas atau produksi telur. Biasanya diukur dalam mikron, milimeter, cm, inchi, miligram, gram, kilogram, persentasi, butir, diameter dan lain'lain. Sebagai contoh berat merupakan suatu fenotipe kuantitatif tunggal dan perbedaannya diukur sebagai gram, kilogram atau dalam unit atau satuan yang dikehendaki.
Karena fenotipe kuantitatif memperlihatkan karakter/variasi yang kontinyu, maka satu'satunya cara untuk mempelajarinya adalah menganlisa varian atau ragam yang ada dalam satu populasi dan memilah ke dalam komponen baian'bagiannya.
Varian atau ragam fenotipe (VP) yang dianalisa untuk suatu ciri/karakter kuantitatif merupakan penjumlahan atau gabungan dari varian genetik atau genotipe (VG), varian atau ragam lingkungan (VE) serta interaksi antara varian genetik dan lingkungan (VGxE). Secara matematik dapat ditulis:
GxE E G
P V V V
V = + +
Varian atau ragam genetik merupakan gabungan atau penjumlahan atau resultan dari varian genetik yang aditif (VA), varian atau ragam genetis yang dominan (VD) dan varian genetis yang epistatis (VI). Secara matematis dapat ditulis:
I D A
G V V V
V = + +
Oleh karena VI sangat sulit diukur bahkan sangat jarang terjadi pada biota seperti pada ikan, maka kadang dianggap nol, sehingga, dapat ditulis:
D A
G V V
V = +
$"%" 8*+ 4 + * - - - 1 2 * *- - - 1
Sejauh ini pembicaraan tentang suatu fenotipe diasumsikan menggambarkan fenotipenya. Fenotipe sifat'sifat demikian mudah dibedakan, misalnya wama kulit buncis bersifat altematif yaitu putih atau abu'abu, tanamannya tinggi atau rendah, sehingga mudah dibedakan. Sifat'sifat demikian dikenal dengan istilah sifat diskontinyu (discontinue traits). Pada sifat'sifat demikian terdapat hubungan yang sederhana antara genotipe dengan fenotipenya. Dalam banyak kasus suatu gnotipe hanya menghasilkan satu macam fenotipe, sebaliknya suatu fenotipe merupakan hasil aktivitas suatu genotipe. Namun demikian telah kita maklumi bahwa hubungan antara genotipe dengan fenotipe dipengaruhi oleh fenomena'fenomena, misalnya ekspresivitas, penetrasi dan pleiotropi. Dengan kata lain suatu genotipe dapat menghasilkan beragam fenotipe karena genotipe berinteraksi dengan lingkungannya selama proses pertumbuha/perkembangan. Sifat'sifat seperti bobot bayi fahir, tinggi orang dewasa, kandungan protein jagung, menunjukkan kisaran fenotipe yang luas.
Sifat'sifat yang menunjukkan kisaran keragaman fenotipe demikian sering disebut sifat kontinyu (continues traits). Pada sifat diskontinyu memiliki distribusi diskrtit, sedangkan sifat kontinyu memiliki distribusi kontinyu, sehingga periu dinyatakan secara kuantitatif. Sifat'sifat demikian sering disebut sifat kuantitatif.
Sifat kontinyu memiliki kisaran fenotipe yang kontinyu pula. Untuk mengkaji pewarisan sifat kontinyu/kuantitatif perlu dilihat mengapa suatu sifat memiliki beragam fenotipe. Sifat kuantitatif dapat terjadi dalam berbagai cara.
Umumnya kisaran fenotipe terjadi karena berbagai macam genotipe ada dalam suatu kelompok individu (populasi). Hal ini biasa terjadi apabila suatu karakter dikendalikan oleh banyak lokus. Misalnya, apabila suatu lokus dengan dua alel per lokus mengendalikan suatu sifat, maka ada tiga kemungkinan genotipe yakni AA, Aa, aa. Untuk dua lokus dengan dua alel per lokus akan diperoleh 9 genotipe (32), misal AABB, AABb, Aabb, AaBB, AaBb, Aabb, aaBB, aaBb, dan aabb.
Demikian pula dengan pertambahan jumlah lokus sebanyak n, maka genotipe yang terbentuk sebanyak 3n. jika terdapat lebih dari dua alel untuk setiap lokus maka akan dihasilkan macam genotipe jauh lebih banyak.
Dengan demikian semakin banyak lokus yang mengendalikan suatu sifat, maka akan semakin banyak macam genotipe yang mungkin terbentuk. Sifat
yang dikendalikan oleh banyak lokus seperti ini disebut sifat poligenik atau sifat kuantitatif. poligenik atau sifat kuantitatif.
Perbedaan dasar antara sifat kualitatif dan sifat kuantitatif melibatkan jumlah gen yang berkontribusi pada variabilitas fenotip dan derajat di mana fenotip itu dapat dimodifikasi oleh faktor'faktor lingkungan. Sifat'sifat kuantitatif dapat diatur oleh banyak gen (mungkin 100 sampai 100 atau lebih), masing' masing berkontribusi terhadap fenotip begitu sedikit sehingga pengaruh' pengaruh individunya tidak dapat dideteksi dengan metode'metode Mendel.
Gen'gen yang bersifat demikian disebut poligen (Stansfield, 1991).
Di bawah ini disajikan ringkasan beberapa perbedaan utama antara genetika kuantitatif dan kualitatif (Stansfield, 1991):
No. Genetika Kuantitatif Genetika Kualitatif
1. Ciri'ciri dari derajat. Ciri'ciri dari jenis.
2.
Variasi kontinu; pengukuran fenotip merupakan suatu spektrum.
Variasi diskontinu; kelas'kelas fenotip yang jelas.
3.
Pengendalian poligenik; pengaruh gen' gen tunggal terlalu kecil untuk dapat dideteksi.
Gen tunggal memberikan pengaruh yang jelas dapat dibedakan.
4.
Mempersoalkan suatu populasi organisme yang terdiri dari segala macam perkawinan yang dapat terjadi.
Mempersoalkan perkawinan+
perkawinan individu dan keturunannya.
5.
Analisis statistik memberikan estimasi (perkiraan) parameter+parameter populasi seperti rata'rata dan deviasi standar.
Dianalisis dengan membuat penghitungan+penghitungan dan rasio+rasio.
Dalam genetika kuantitatif, konsep poligen (polygenes, berarti “banyak gen“) digunakan untuk menjelaskan terbentuknya sifat kuantitatif. Ronald Fisher (1918) dapat menjelaskan bahwa sifat kuantitatif terbentuk dari banyak gen dengan pengaruh kecil, yang masing'masing bersegregasi menuruti teori Mendel. Karena pengaruhnya kecil, fenotipe yang diatur oleh gen'gen ini dapat dipengaruhi oleh lingkungan. Meskipun demikian, penjelasan Fisher ini tetap menempatkan “gen'gen” yang mengatur sifat kuantitatif sebagai sesuatu yang abstrak karena hanya merupakan konsep. Langkah pembuktian mengenai
adanya gen'gen yang mengatur sifat kuantitatif mulai terbuka setelah tersedianya banyak penanda genetik sehingga memungkinkan orang membuat peta pautan genetik yang dapat menjangkau sebagian besar kromosom.
Penanda'penanda genetik digunakan untuk menunjukkan situasi alelik pada bagian kromosom tertentu. Variasi alel pada suatu penanda menjadi genotipe bagi kromosom atau kelompok pautan (apabila kromosomnya belum teridentifikasi) (Rohmad, 2012).
$":" 8*+ 4 + ; + - ) - 5 - +
Menurut Nasir (2001) heritabilitas adalah proporsi ragam genetik terhadap besaran total ragam genetik ditambah dengan ragam lingkungan, dengan kata lain heritabilitas merupakan proporsi besaran ragam genetik terhadap besaran ragam fenotipe untuk suatu karakter tertentu. Ada dua nilai heritabilitas yang dikenal dalam pemuliaan tanaman yaitu heritabilitas dalam arti luas dan heritabilitas dalam arti sempit (Alif, 2008).
Nilai heritabilitas dalam arti luas memperhatikan ragam genetik total dalam kaitannya dengan keragaman fenotipe. Dalam hal ini genotipe dianggap sebagai unit dalam kaitannya dengan lingkungan. Sementara itu heritabilitas dalam arti sempit yang menjadi fokus perhatian adalah keragaman yang diakibatkan oleh peran gen aditif merupakan bagian dari keragaman genetik total. Berdasarkan penjelasan ini dapat dipahami bahwa nilai heritabilitas dalam arti sempit tidak akan pernah lebih besar dibandingkan dengan nilai heritabilitas dalam arti luas untuk suatu karakter tertentu (Alif, 2008).
Parameter heritabilitas melibatkan semua tipe aksi gen dan oleh karena itu membentuk suatu perkiraan heritabilitas yang luas. Pada kasus dominansi sempurna, bila suatu gamet yang mengandung alel dominan aktif A2 berpadu dengan suatu gamet yang mangandung alel A1 nol, fenotip yang dihasilkan bisa terdiri atas dua unit. Bila dua gamet A2 berpadu, hasil fenotipnya tetap akan terdiri atas dua unit. Sebaliknya, jika gen'gen yang tidak mempunyai dominansi (gen'gen aditif) terlibat, maka gamet'gamet A2 akan menambah satu unit kepada fenotip dari zigot yang dihasilkan, dengan tidak memandang kontribusi alel dari gamet yang berpadu dengannya. Jadi hanya komponen aditif genetik dari variansi yang mempunyai kualitas dapat diramalkan, yang perlua pada formulasi renca'renca pemuliaan. Heritabilitas dalam arti yang lebih sempit ini adalah rasio variansi aditif genetik terhadap variansi fenotip (Stansfield, 1991).
Harus ditekankan bahwa heritabilitas suatu sifat hanya berlaku pada populasi tertentu yang hidup dalam suatu lingkungan khusus. Suatu populasi yang secara genetik berbeda (mungkin suatu varietas bangsa, ras yang berbeda atau subspesies dari spesies sama) yang hidup dalam lingkungan yang identik, kemungkinan besar mempunyai heritabilitas yang berbeda bagi sifat yang sama.
Begitu pula, populasi yang sama kemungkinan besar memperlihatkan heritabilitas yang berbeda bagi sifat yang sama bila diukur dalam lingkungan' lingkungan yang berbeda, karena suatu genotip tertentu tidak selalu memberikan respon terhadap lingkungan'lingkungan yang berbeda dengan cara yang sama. Tidak ada satu genotippun yang mempunyai daya adaptasi yang superior dalam segala macam lingkungan. Hal inilah yang menyebabkan mengapa seleksi alam cenderung menimbulkan populasi'populasi yang secara genetik berbeda dalam suatu spesies, suatu populasi beradaptasi secara khas terhadap kondisi'kondisi setempat dan tidak secara umum beradaptasi terhadap semua lingkungan dimana spesies itu ditemukan (Stansfield, 1991).
Persoalan yang selalu muncul pada sifat'sifat kuantitatif adalah pertanyaan tentang seberapa jauh suatu sifat dikendalikan secara genetik dan seberapa jauh porsi pengendalian oleh lingkungan. Dengan demikian pertanyaan yang selalu berulang adalah bagaiman imbangan pengaruh alami (genetik) versus pemeliharaan (nature versus nurture) atau pengaruh gen versus lingkungan. Hal inilah yang menjadi fokus pembahasan genetika kuantitatif.
Dengan demikian persoalannya dapat dinyatakan sbb. Seberapa besar variasi suatu karakter (fenotipe) yang teramati (Vp) disebabkan oleh variasi genetik (Vg) dan variasi lingkungan (Ve) atau sering dituliskan dengan formulasi yang sederhana sbb. Vp = Vg + Ve. Untuk mengukur suatu variasi fenotipe dan memilahnya menjadi variasi genetik dan variasi lingkungan dipertukan metode statistik.
Heritabilitas adalah proporsi variasi fenotipik populasi yang disebabkan oleh faktor genetik. lsitilah ini seringkali disalah gunakan. Misalnya apabila individuindividu dalam suatu famili memiliki kemiripan dalam suatu aspek fenotipe, misalnya perawakan atau intelgensia, hal ini sering dianggap disebabkan oleh faktor genetik. Akan tetapi sering tejadi kemiripan tersebut semata'mata sebagai konsekuensi dari interaksi genotipe dengan lingkungan yang sama. Konsep heritabilitas dimaksudkan untuk menilai suatu karakter kuantitatif dalam kaitannya dengan kontribusi relatif faktor genetik dan
lingkungan pada suatu sifat tertentu. Misalnya seberapa jauh efek gen mempengaruhi kebiasaan, misal kebiasaan minum alkohol, hal ini akan bermanfaat dalam membuat kebijakan sosial tetapi hal ini perlu ditafsirkan dengan sangat hati'hati karena besar kemungkinan terjadi penyalahgunaan tafsir. Untuk menilai suatu heritabilitas, langkah pertama yang harus dilakukan adalah mengukur variasi fenotipe suatu sifat, kemudian dilajutkan dengan pemerian varians tersebut kedalam berbagai sumber ragam penyebabnya.
Varians fenotipe dapat diperikan menjadi varians genetik, varians lingkungan, kovarians genetik x lingkungan dan varians interaksi genetik x lingkungan yang dapat diformulasikan dalam persamaan sedehana sbb.:
Vp = Vg + Ve + COVgxe + Vg+e.
Oleh karena kovarians g x e biasanya bemilai kecil maka biasa diabaikan dan dianggap nol oleh para ahli genetika. Pada komponen Vg dapat diperikan menjadi dianggap nol oleh para ahli genetika. Pada komponen Vg dapat diperikan menjadi varians aditif (VA) dan varians dominan (VD) dengan memperhatikan interaksi tindak gen dalam lokus. Apabila terjadi inteaksi tindak gen antarlokus, biasa dikenal dengan isitilah epistasis, maka komponen varians genetik bertambah lagi dengan varians epistasis, V1. Dengan demikian varians fenotipe menjadi :
Vp=VA+VD+VI+Ve+COVgxe+Vg+e.
Untuk memerikan varians fenotipe menjadi berbagai komponen varias seperti di atas diperlukan rancangan percobaan yang lebih khusus.
Secara umum ada dua jenis heritabilitas yang dikenal orang, yaitu heritabilitas arti luas (broad'sense heritability) dan heritabilitas arti sempit (narrow'sense heritability), yang masing'masing disimbulkan sbb.: heritabilitas arti luas (broad'sense heritability) = H = h2B = Vg/Vp, dan heritabilitas arti sempit (narrow'sense heritability) = h =h2N = VA/Vp.
Lepas dari manfaatnya, parameter heritabilitas memiliki berbagai keterbatasan yang biasanya diabaikan orang, sehingga terjadi misused. Oleh karena itu konsep heritabilitas sering disebut sebagai suatu parameter yang paling sering disalah mengerti (misunderstood) dan disalahgunakan (misused) di bidang genetika. Di bawah ini dikemukan beberapa kualifiaasi dan keterbatasan konsep heritabilitas yang perlu diperhatikan:
1. heritabilitas tidak mengisyaratkan bahwa suatu sifat dikendalikan secara genetik. Apa yang terukur dari parameter heritabilitas hanyalah proporsi
varians fenotipe diantara individu dalam suatu populasi karena perbedaan genotipenya. Hal inilah yang sering disalah mengerti dan dianggap sebagai suatu sifat dikendalikan genetik. Suatu gen sering mempengaruhi perkembangan suatu sifat, sehingga dapat dikatakan karakter tersebut bersifat genetik. Namun demikian perbedaan fenotipe antar individu dalam suatu populasi yang terukur pada parameter heritabilitas sama sekali bukan bersifat genetik.
2. heritabilitas diukur berdasarkan nilai varians yang hanya dapat dihitung untuk suatu kelompok individu, sehingga heritabilitas adalah karakteristik suatu populasi bukan karakter suatu individu.
3. heritabilitas tidak bersifat tetap untuk suatu karakter. Tidak ada nilai tertentu yang beriaku umum bagi suatu karakter. Nilai heritabilitas suatu karakter sangat tergantung pada perubaan genotipe dan lingkungan dimana genotipe tersebut tumbuh.
4. bahkan apabila ada dua populasi dimana masing'masing populasi memiliki nilai heritailitas tinggi untuk karakter yang sangat berbeda, maka tidak dapat dikatakan bahwa kedua populasi tersebut berbeda secara genetik.
5. suatu karakter yang dimiliki oleh individu'individu dalam suatu famili tidak selalu memiliki nilai heritabilitas tinggi. Karakter demikian disebut karakter famili (familial trait). Karakter famili bisa disebabkan oleh gen yang sama ataupun oleh lingkungan yang sama, sehingga familiality tidak identik dengan heritability.
$"<" 8*+ 4 + ! + * + 1 -7+ 1 - *- - - 1
Pada awalnya para ahli kesulitan memahami bagaimana suatu karakter kuantitatif diwariskan, kecuali mereka sekedar memahami bahwa pola pewarisan karakter kuantitatif berbeda dari karakter kualitatif (karakter diskontinyu).
Kajian yang pernah dilakukan para ahli tentang pewarisan kuantitatif adalah karakter panjang tongkol pada jagung. Pada tahun 1913, Rollins Emerson dan Edward East membuat persilangan jagung varietas jagung manis yang memilkii tongkol pendek dengan varietas jagung popcorn (jagung brondong) yang memiliki tongkol panjang, kemudian membuat sating silang sesama individu F1.
Dari percobaan tersebut mereka melaporkan bahwa sifat'sifat kuantitaif, dengan mengabaikan pengaruh lingkungan, memiliki karakteristik sbb.:
1. nilai rerata sifat kuantitatif pada populasi F1 mendekati nilai tengah antar kedua tetua.
2. nilai rerata sifat kuantitatif pada populasi F2 mendekati nilai rerata pada populasi F1'nya.
3. pada populasi F2 menunjukkan kisaran yang lebih luas dibanding populasi F1.
4. nilai'nilai ekstrim pada populasi F2 lebih jauh dari nilai ekstrim pada populasi F1, mendekati nilai'nilai ekstrim kedua tetuanya.
Dari data tersebut tidak dapat diterangkan menggunakan prinsip'prinsip genetika Mendel yaitu kaidah `satu gen'satu lokus' yang mengendalikan pewarisan karakter diskontinyu. Dengan demikian Emerson dan East mengemukankan hipotesis bahwa karakter tersebut dikendalikan oleh banyak gen/lokus. Hipotesis ini dikenal dengan sebutan hipotesis poligenik atau multiple gen untuk pewarisan kuantitatif (polygene or multiple'gene hypothesis for quantitative inheritance). Pada karakter kuantitatif tidak ada dominansi antar alel dalam suatu lokus, sehingga ekspresi suatu karakter tergantung pada berapa banyak alel yang berpengaruh terhadap fenotipe dikandung oleh genotipe yang bersangkutan. Dalam perspektif ini dikenal istilah alel berefek (contributing alleles) dan alel tidak berefek(noncontributing alleles). Misalnya pada karakter warna biji gandum, untuk genotipe RRCC (memiliki 4 alel berefek), maka fenotipenya akan berwarna merah gelap, sedang genotipe rrcc (tidak memiliki alel berefek) maka fenotipenya akan berwarna putih. Fenotipe wama dari suatu genotipe merupakan fungsi dari seberapa banyak genotipe tersebut memiliki alel berefek (dalam hal ini R dan C). antung pada berapa banyak alel yang berpengaruh terhadap fenotipe dikandung oleh genotipe yang bersangkutan. Dalam perspektif ini dikenal istilah alel berefek (contributing alleles) dan alel tidak berefek(noncontributing alleles). Misalnya pada karakter warna biji gandum, untuk genotipe RRCC (memiliki 4 alel berefek), maka fenotipenya akan berwarna merah gelap, sedang genotipe rrcc (tidak memiliki alel berefek) maka fenotipenya akan berwarna putih. Fenotipe wama dari suatu genotipe merupakan fungsi dari seberapa banyak genotipe tersebut memiliki alel berefek (dalam hal ini R dan C).
$"#(" 8*+ 4 + *
Pemuliaan (breeding) merupakan penerapan biologi, terutama genetika, dalam bidang peternakan untuk memperbaiki produksi atau kualitas. Ilmu ini relatif baru dan lahir sebagai implikasi berkembangnya pemahaman manusia atas asas'asas pewarisan sifat secara genetik. Secara umum, ilmu ini berusaha menjelaskan dan menerapkan prinsip'prinsip genetika (dengan bantuan cabang' cabang biologi lain) dalam kegiatan pemuliaan. Dalam prakteknya pemulian ternak menerapkan ilmu genetika, statistika dan biometrika serta reproduksi ternak, dengan tujuan untuk memperbaiki mutu genetik ternak, sehingga dapat meningkatkan produksi atau memberikan nilai tamba dalam pelaksanaannya.
Dari ilmu genetika, terkait dengan aspek penurunan sifat dari tetua kepada keturunannya. Termasuk dalam hal ini adalah konsep'konsep hokum Mendel. Statistika dan biometrika berperan dalam pengukuran keragaman sifat dan penyebarannya, hubungan antara dua sifat atau lebih, serta analisis untuk pendugaan parameter'parameter genetik. Reproduksi terkait dengan aspek fertilitas, kebuntungan, jarak beranak dan kelahiran
Berbagai cabang genetika menemukan aspek praktis dalam ilmu pemuliaan, seperti: genetika populasi (misalnya dalam strategi persilangan), genetika kuantitatif (misalnya dalam teori seleksi), sitogenetika (misalnya dalam penggunaan organisme poliploid atau mutan), dan genetika molekular (misalnya dalam penggunaan penanda molekular atau transfer gen).
Kegiatan pemuliaan lebih merupakan kombinasi antara ilmu dan seni yang telah dilakukan manusia ribuan tahun lalu, misalnya: seleksi dan konservasi jagung oleh orang Indian di Meksiko dari teosinte, pembiakan ulat sutera di daratan Tiongkok yang menghasilkan serat sutera yang panjang, pemurnian berbagai ras anjing melalui seleksi terhadap serigala, persilangan kuda dengan keledai yang menghasilkan bagal, atau persilangan itik dengan itik manila (mentok) yang menghasilkanbrati.
$"##" -82 *2 - * * + + * - *- - - 1
$"##"#" -82 * + + - 8 18 -
Data morfometrik yang diukur antara lain: berat, panjang, lebar, tinggi, tebal cangkang, luas, keliling, dst. Analisis rasio/indeks morfomterik dilakukan untuk mendapatkan koefisien keragaan antar individu dalam satu populasi
dengan menghitung koefisien jarak (nilai Eucleudian), Koefisien kemiripan (SSm) dan Koefisien keragaman (KK). Nilai'nilai koefisien tersebut dipetakan dalam dendrogram untuk melihat ada tidaknya kemiripan dan perbedaan antar individu dalam populasi tiram mutiara.
$"##"." -82 * + + 3 * 8
Keragaan biokimia yang diukur adalah analisis enzym (allozyme), protein, Asam Amino dengan menggunakan starch gel electrophoresis. Analisis laju migrasi buffer dan prosedur stainning mengikuti metode Sugama et. all (1988). Fitness test untuk kesetimbangan Hardy'Weinberg dan perbedaan antar populasi menggunakan softwares GENEPOP dan GenStat (Raymond and Roussell, 1995). Interpretasi penamaan lokus dan alel mengikuti metode Allendorf dan Utler (1979). Penamaan alel didasarkan atas jarak relatif migrasi alel tersebut terhadap alel yang paling sering muncul (modus) yang diberi kode 100. Data diinterpretasi sehingga didapatkan frekwensi alel, derajat polimorfisme, heterozigositas dan jarak genetik dengan menggunakan formula Nei (1987)
di mana:
= Nilai statistik mahalonobis sebagai ukuran jarak kuadrat genetik antar dua populasi
= Kebalikan matriks gabungan ragam peragam antar peubah = Vektor nilai rataan pengamatan dari populasi i masing'masing
peubah
= Vektor nilai rataan pengamatan dari populasi i masing'masing peubah
Analisis diskriminan digunakan untuk melihat perbedaan yang signifikan antar kelompok yang ditentukan oleh karkter morfometrik yang diamati. Hasil pengukuran direlatifkan dengan menggunakan standar deviasi untuk menghindari keragaman ukuran dan kemungkinan umur yang berbeda.
Selanjutnya akan dikelompokan karakter morfologi yang paling mencirikan perbedaan tersebut. Adanya pengaruh interaksi sesama karakter yang diukur mengakibatkan penyusutan karakter'karakter tersebut. Komponen ini juga digunakan untuk melihat pengaruh yang paling kuat terhadap pembentukan
) (
)
(
1) 2(
j i j
ij
X
iX C X X
D = −
−−
) ( 2
D
ij−1
C X
iX
jsuatu populasi. Analisis ini menggunakan program Minitab versi 17 dan program SPSS versi 20 yang berdasarkan Analisis Komponen Utama (Principal Component Analysis/PCA). Analisis PCA bertujuan untuk menginterpretasikan sebagian besar informasi yang terdapat dalam suatu matriks data ke dalam bentuk grafik dan untuk mendapatkan peta penyebaran populasi serta nilai'nilai kesamaan di dalam maupun di luar kelompok. Hasil yang diperoleh ditampilkan dalam bentuk dendrogram.
$"##" $ -82 * + + - 3 * 8
Ekstrak DNA dan RNA total mengikuti metode baku yang dikembangkan oleh Farajallah et. all (1999). Amplifikasi dan visualisasi DNA dan RNA menggunakan metode PCR dan elektroforesis dengan primer yang disesuaikan dengan sampel. Sekuensing DNA dan RNA hasil PCR dengan menggunakan mesin Sequenzer ABI377A Applied Biosystem. Hasil runutan DNA dan RNA diedit secara manual menggunakan software Genetyx versi 4.01, kemudian dijajarkan dengan menggunakan software Clustal W.Versi 1.7 (Higgins et. all, 1997).
Karakterisasi runutan DNA serta analisisnya menggunakan metode maximum parsimony (MP) dan Neighbor Joint (NJ) yang dilakukan dengan menggunakan software MEGA versi 2.0. (Kumar et. all, 2001). Untuk mengetahui konsentrasi DNA dan RNA dilakukan pengukuran dengan menggunakan program Gene quant. Hasil konsentrasi genom DNA dan RNA masing'masing sampel dikalkulasi lebih lanjut untuk penentuan rasio RNA – DNA.
$"##" '" -82 * *2
Beberapa penanda DNA yang dapat digunakan untuk menganalisis variasi genetika, evolusi dari suatu tanaman, keterpautan gen tertentu terhadap karakter spesifik, penelusuran tetua, analisis lokus'lokus karakter kuantitatif, dan revisi klasifikasi tanaman adalah Restriction Fragment Length Polymorphism (RFLP), Random Amplified Polymorphic DNA (RAPD),Amplified Fragment Length Polymorphism (AFLP), dan mikrosatelit atau Simple Sequence Repeat (SSR).
! " ! # $
Penelitian untuk mengeksploitasi polimorfisme DNA pada genom tanaman dengan memanfaatkan teknologi marka molekuler jumlahnya terus meningkat.
Restriction Fragmen Length Polymorphism (RFLP) adalah teknik molekular yang
didasarkan pada polimorfisme yang diakibatkan oleh adanya substitusi basa (nukleotida), insersi, dilesi, ataupun translokasi yang kemungkina terjadi pada masa lalu. Teknik ini memanfaatkan situs'situs restriksi spesifik yang terdapat pada genom suatu organisme.
Enzim restriksi atau endonuklease restriksi adalah enzim yang memotong untaian DNA pada rangka gula'fosfat tanpa merusak basa, sesuai urutan yang dikenalinya Sekuen pengenalan sering disebut situs pengenalan merupakan sekuen DNA yang menjadi tempat menempelnya enzim restriksi dan melakukan pemotongan pada sekuen tersebut. Enzim'enzim tersebut dapat dipesan melalui perusahaan'perusahaan yang bergerak di bidang bioteknologi dan biologi molekuler. Hasil pemotongan genom menggunakan enzim restriksi tertentu akan menghasilkan perbedaan panjang fragmen DNA, yang menunjukkan jarak dari situs'situs restriksi enzim tersebut dalam suatu genom organisme. Genom yang telah terfragmentasi kemudian dapat dianalisis sesuai tujuan penelitian. Melalui analisis selanjutnya dapat diketahui apakah pada sekuen target telah terjadi perubahan akibat adanya substitusi basa (nukleotida), insersi, dilesi, ataupun translokasi.
% & " ! '(% # % '$
RAPD banyak digunakan untuk menganalisis keanekaragaman karakter genetik dalam berbagai penelitian dengan pertimbangan antara lain tidak membutuhkan latar belakang pengetahuan tentang genom yang akan dianalisis, primer yang digunakan bersifat universal (dapat digunakan untuk prokariot maupun eukariot), mampu menghasilkan karakter yang relatif tidak terbatas jumlahnya, bahan'bahan yang digunakan relatif lebih murah, preparasi lebih mudah,dan memberikan hasil lebih cepat dibandingkan dengan analisis molekular lainnya.
Metode RAPD mampu mendetekasi sekuen nukleotida dengan hanya menggunakan satu primer. Primer tersebut akan berikatan dengan utas tunggal genom yang satu dan pada utas DNA pasangannya dengan arah berlawanan. Selama situs penempelan primer masih berada pada jarak yang dapat diamplifikasi pada umumnya tidak lebih dari 5000 pasangan basa (pb), maka akan diperoleh produk DNA amplifikasi. Dengan penggunaan RAPD yang relatif mudah, murah, dan menganalisis dalam tingkat DNA, maka seleksi dini dapat dilakukan terhadap sifat'sifat ketahanan terhadap cekaman biotik dan
abiotik, karakter'karakter yang bersifat kuantitatif, dan duplikasi tanaman koleksi dapat dihindari.
# ) * $
DNA ruas berulang yang memiliki variasi paling tinggi dalam genom tanaman adalah sekuen berulang dengan fragmen berulang sederhana atau pendek. Fragmen ini dikenal dengan nama minisatelit dan mikrosatelit.
Minisatelit adalah DNA yang memiliki pengulangan biasanya antara 10'60 pasang basa, yang pada awal penemuannya banyak diaplikasikan pada genom manusia. Sedangkan Mikrosatelit (SSR) memiliki unit berulang lebih sedikit berkisar antara1–6 pasang basa, terdapat dalam jumlah sangat banyak dan menyebar di dalam genom, dan banyak digunakan pada tanaman. Variasi fragmen'fragmen ini biasanya merupakan hasil perubahan dalam jumlah kopi dari perulangan asal dan sering dikategorikan sebagai Variable number of tandem repeats (VNTR). Karena level polimorfisme yang sangat tinggi dapat dideteksi dengan fragmen ini, VNTR diakui sebagai alat yang manjur untuk finger printing dan identifikasi kultivar tanaman. Fragmen ini juga dapat digunakan untuk mempelajari keragaman antar dan intra populasi, studi ekologi, menghitung jarak genetik, dan mempelajari evolusi tanaman. Sekuen mikrosatelit DNA yang pendek dengan sekuen DNA pengapit bersifat conserved, memungkinkan mendesain primer untuk mengamplifikasi situs'situs spesifik menggunakan PCR. Jika primer'primer tersebut digunakan mengamplifikasi lokus'lokus SSR tertentu, maka setiap primer akan menghasilkan polimorfisme dalam bentuk perbedaan panjang hasil amplifikasi yang dikenal dengan SSLP (Simple Sequence Length Polymorphism). Setiap panjang mewakili satu alel dari suatu lokus. Perbedaan panjang terjadi karena perbedaan jumlah unit pengulangan pada lokus'lokus SSR tertentu Keanekaragaman jumlah ulangan pada mikrosatelit dapat dideteksi dengan mengelektroforesis produk DNA yang sudah diamplifikasi di dalam sekuen gel standar, yang dapat memisahkan fragmen'fragmen yang membedakan tiap'tiap nukleotida. Mikrosatelit DNA terdapat dalam jumlah banyak dan menyebar di dalam genom. Bentuk umum pengulangan Mikrosatelit DNA (SSR) adalah pengulangan dua basa secara sederhana. Mikrosatelit DNA (SSR) dengan kemudahan dan kecepatan menggunakan teknologi PCR, bersifat kodominan, dan mudah diinterpretasikan membuat mikrosatelit menjadi penanda paling baik dalam pemetaan gen.
% '
Data yang dihasilkan dari analisis molekular sering digunakan sebagai dasar untuk klasifikasi individu pada suatu populasi, untuk konstruksi filogenetik, penelusuran tetua, identifikasi penanda suatu sifat tertentu, revisi taksonomi dan lain'lain. Dendogram yang dikonstruksi dalam analisis tersebut umumnya didasarkan pada tingkat kesamaan antara individu yang digunakan. Namun, batas kepercayaan untuk pengelompokan yang dihasilkan melalui dendogram, biasanya tidak dapat diterima untuk perhitungan menggunakan prosedur statistik yang umum. Data molecular untuk dapat dianalisis harus diubah ke dalam bentuk data biner berdasarkan ada dan tidaknya pita hasil amplifikasi.
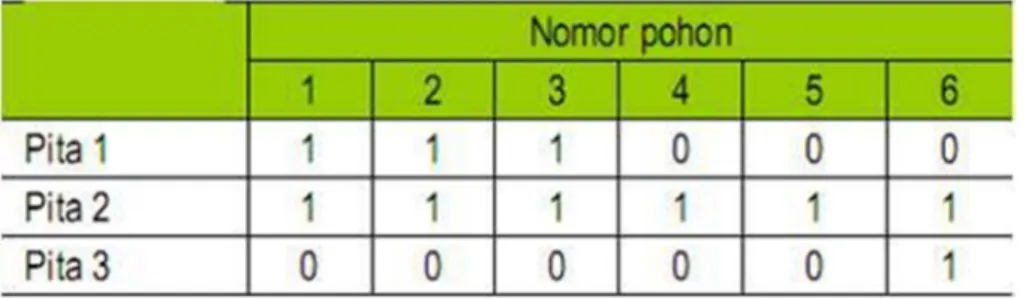
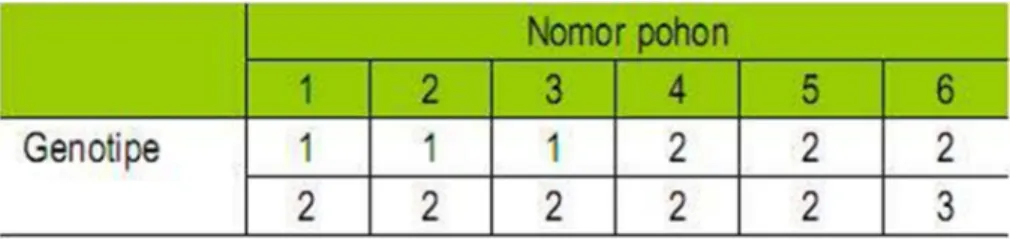
Jika terdapat pita hasil amplifikasi diberi skor 1, sedangkan jika tidak ada diberi skor 0. Data biner yang diperoleh dapat digunakan untuk menyusun dendogram atau filogenetik dari orgsnisme yang dianalisis. Untuk penelusuran tetua, pemetaan genetik, dan indentifikasi fragmen spesifik data biner diubah ke dalam data genotipe, dan dengan program khusus dianalisis sesuai tujuan. Contoh persiapan pengolahan data menggunakan penanda molekular, disajikan pada Gambar berikut.
+ , ! '(%
Tabel 2. Contoh Data biner yang diturunkan dari gambar 3 pada turunannya.
Tabel 3. Contoh Data genotipe yang diturunkan dari Tabel 1.
Hasil pola pita molekular yang telah diubah ke dalam bentuk data biner dapat diolah menggunakan Program computer NTsys ver. 2.01 untuk mendapatkan matriks jarak genetik dan dendogram atau filogenetik dari sampel.
Untuk menganalisis data sesuai tujuan penelitian, saat ini tersedia sangat banyak program/software yang dapat digunakan untuk mengolah data molekular.Sebagai contoh, Data genotipe yang disusun dari data biner (Tabel 2) dapat digunakan untuk menganalisis hubungan tetua dengan zuriatnya (pelacakan tetua), peta keterpautan (linkage map), dan korelasi antar lokus dengan lokus dan lokus dengan sifat morfologi. Pelacakan tetua menggunakan program Cervus 2.0 (Marshall,1998), depresi silangdalam berdasarkan penanda molekular menggunakan program POPGENE ver. 1.32 (Yeah et al., 2001), pembuatan peta keterpautan menggunakan MAPMAKER dengan nilai LOD 3, dan korelasi antar lokus dengan lokus dan antara lokus dengan sifat morfologi menggunakan Program Minitab 14.
'" =
/ 0"
'"#" * 2 * *3+ * + + 2 *3 - 4 18 *+ 5 8- -
Dengan menganalisis fenotipe kualitatif dan atau kuantitatif, diharapkan dapat menemukan pola'pola keteraturan dalam suatu performans sifat/karakter kuantitatif yang belum ditemukan sebelumnya.
Dalam satu populasi, fenotipe'fenotipe kuantitatif digambarkan oleh kecenderungan sentral serta sebaran atau distribusinya di sekitar kecenderungan tersebut. Misalnya, rata'rata (mean), ragam (varian), simpangan baku (standard devisasi), koefisien varian (co'varian), serta sebaran (range atau interval).
Pewarisan sifat/karakter kuantitaif sangat rumit, tidak sama halnya dengan fenotipe kualitatif yang dapat dikontrol oleh satu atau sepasang gen tunggal . Fenotipe kuantitatif dikontrol oleh 20 atau 50 bahkan lebih dari 100 gen. Bahkan jumlah gen yang mengontrol fenotipe kuantitatif kadang tidak diketahui dan cara ekspresi gennya pun tidak diketahui.
Padahal pewarisan fenotipe kuantitatif, seperti produksi telur, ukuran tubuh, bobot tubuh, pertumbuhan dan kelulusan hidup, sangat besar pentingnya dalam upaya pemuliaan ikan, baik ikan konsumsi maupun ikan umpan. Hal ini berbanding terbalik dengan pemuliaan ikan hias yang lebih mengutamakan fenotipe kualitatifnya seperti warna dan bentuk tubuh ikan.
'"." * 2 * *3+ * + + 2 + + 5 8- -
Seleksi merupakan suatu program yang dengannya individu'individu atau keluarga yang dipilih, dalam usaha untuk mengubah rata'rata populasi pada generasi berikutnya atau dengan kata lain, seleksi adalah memilih individu' individu untuk dijadikan calon tetua dari generasi berikutnya.
Seleksi biasanya didasarkan pada tingkat performan atau indeks indeks yang minimal. Ikan atau individu yang melampaui performans atau indeks minimal akan diseleksi sebagai stok induk sedangkan individu yang berada di bawah indeks atau performans minimal akan dibuang atau diabaikan. Artinya
bahwa rasio atau indeks'indeks yang diukur atau dihitung merupakan nilai performans individu'individu dalam satu populasi.
Dalam suatu program seleksi karakter/sifat kuantitatif yang dinilai atau dianalisis adalah varian genetis apa yang bertanggung jawab untuk memproduksi karakter atau sifat'sifat kuantitatif tersebut yang harus ditransmisikan dari tetua ke anaknya dalam cara yang dapat diprediksi dan dipercaya. Artinya, prediksi varian genetis yang diproduksi dalam bentuk rasio' rasio atau indeks'indeks di antara sifat/karakter kuantitatif yang muncul.
Misalnya, untuk memprediksi pewarisan sifat'sifat seperti:
a. Jumlah produksi telur (fekunditas) seekor ikan, maka salah satu sifat/karakter kuantitatif yang dikur adalah performans atau indeks ukuran gonad dan ukuran telur. Karenanya sifat/ragam genetis apa yang memproduksi atau menyebabkan atau mengontrol perkembangan ukuran gonad dan telur ikan tersebut perlu diukur atau dianalisis.
b. Laju pertumbuhan, maka salah satu karakter kuantitatif yang dilihat atau diukur adalah performans atau indeks'indeks ukuran lambung, panjang dan diameter usus, ukuran lebaran mulut, dan lain'lain, karena untuk menyebabkan atau memproduksi sifat'sifat tersebut ada ragam gen yang mengontrol atau mengaturnya.
c. Kelulusan hidup atau peka terhadap kondisi lingkungan atau kemampuan homeostatis, maka salah satu sifat kuantitatif yang diukur adalah performans jantung dan alat respirasi lainnya, karena ada ragam genetis yang memproduksi atau mengontrolnya.
d. Kemampuan atau efisiensi konversi makanan, maka sifat kuantitatif yang diprediksi adalah alat'alat atau saluran pencernaan yang terlibat dalam proses pencernaan ini, dan semua ini diontrol oleh sejumlah bahkan banyak ragam gen yang terlibat.
Artinya ragam atau varian ukuran lambung, gonad, jantung, usus, dan lain'lain merupakan ekspresi dari ragam genetis yang mengontrol dan memproduksinya.
Dari ketiga varian atau ragam genetis yakni varian aditif (VA), varian dominan (VD dan varian epistatis (VI), hanya varian atau ragam aditif (VA) yang dapat ditransmisikan dalam cara yang dapat diramalkan/dipredikasi dan
dipercaya, karena VA adalah fungsi allela, sedangkan VD merupakan fungsi genotipe yakni interaksi antara dan antar alella pada tiap lokus. Sehingga VA inilah yang kadang dianggap sebagai faktor penentu dalam breeding values (nilai pemuliaan) suatu individu. Artinya nilai performans atau indeks'indeks yang dipakai adalah varian lebar (range) yang dianggap sebagai VA. Apakah sebaran minimum atau maksimum yang akan dijadikan sebagai patokan sangat bergantung pada tujuan yang ingin dicapai misalnya apakah untuk ikan hias atau ikan konsumsi, sedangkan nilai rata'rata (mean) atau bahkan modus merupakan frekwensi sifat/karakter yang paling banyak atau sering muncul dapat digunakan untuk memprediksi atau meramal varian genetis yang dimiliki individu tersebut.
Deviasi individu yang dilihat dari performans indeks'indeks jantung, lambung, daging, tulang, berat tubuh, tinggi tubuh, dan lain'lain dapat dijadikan sebagai varian aditif yang selanjutnya dibandingkan atau dikonversi dengan ukuran/sifat/karakter ideal yang dimiliki individu yang besar dan sehat dapat dijadikan sebagai paramater untuk program seleksi yang selanjutnya dicari jarak genetik dan clusternya.
Selain itu, sebaiknya performans indeks'indeks untuk seleksi diupayakan pada sifat/karakter kuantitatif yang bisa diukur pada saat ikan atai individu hidup dan bukan saat dimatikan. Misalnya, pengukuran indeks'indeks sirip, diameter mata, panjang kepala, ukuran tubuh, berat tubuh, dan lain'lain.
5 *3 * *
Penerapan ilmu pemuliaan (dan cabang'cabang ilmu peternakan lainnya) telah mengubah peta peternakan pada abad ke'20 jauh berbeda dari masa' masa sebelumnya; dari peternakan yang rentan terhadap lingkungan menjadi peternakan yang lebih terkendali dan bisa dikalkulasi hasilnya. Sebagai contoh:
• penggunaan varietas hibrida dengan memanfaatkan gejala heterosis, yang melipatgandakan hasil tanaman pangan (dikembangkan teknologinya oleh J.W. Shull)
• revolusi hijau pada gandum (1950'an) dan diikuti dengan padi, yang berhasil mengembangkan varietas yang berumur pendek, tanggap terhadap pupuk namun berdaya hasil tinggi
• seleksi berbasis prinsip genetika yang menghasilkan sapi dengan produksi susu atau daging yang meningkat tajam.