Most diaphragms common to trepostomic autozooecia apparently formed floors for the living chambers of successive functioning bodies in the degeneration-regeneration cycle. Perhaps these more closely spaced structures reflect an increase in the length of the soft parts during a single functional phase of the degeneration-regeneration cycle. Investigation of the nature of the soft parts which are included in the Paleozoic autozooecia is necessary if useful conclusions are to be drawn as to mode of growth.
However, the weight of the evidence indicates that the only reasonable assumption at this time places the trepostomes in the Bryozoa. Unfortunately, an indeterminate portion of the end of an exposed autozooecium is usually removed by abrasion. The width of the soft parts perpendicular to the zooid axis was slightly increased to fill the living room.
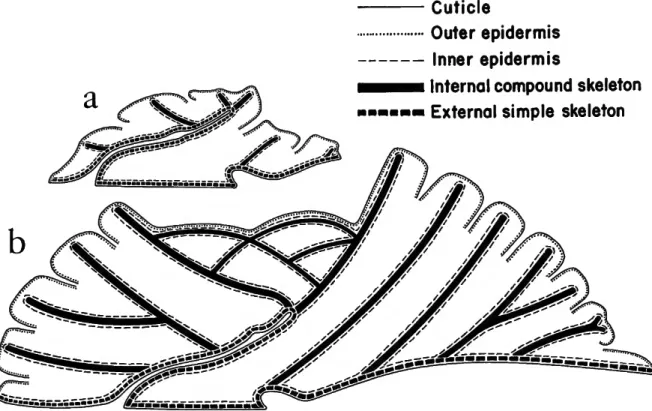
T he exoskeleton of the ancestral zoo is extended distally beyond the fold in the outer wall by composite internal walls (Figure 2. In addition, new zoos arise from the basal layer of the colony and their exoskeletal walls are also composite and internal.
Cuticle
Outer epidermis - Inner epidermis
Internal compound skeleton - External simple skeleton
The most obvious interpretation of the outer tubes of the pairs in the Devonian forms seems to be. Note the change in outline of the sphincter muscle mass (Plate 1: level B of Figures 5b–d) as the tentacles were extruded. The thickness of most of the organic material (Plate 4: figures 2a, b, d) is significantly greater than the thickness of the membranes of the structures in the last studied cyclostomes.
These measurement comparisons indicate that the funnels in the single funnel samples correspond to the inner funnels of the more complex samples. T he walls of most of the funnels do not show the brown or yellow color commonly found in the Devonian Antarctic species. Structural continuity with autozooecial walls requires funnel-cystiphragms to be deposited on the inner wall of the coelom.
The shape of intrazooecial structures (Plate 8: Figures 1–4) found in diaphragm genera are practically identical to those (Plate 6) of the examined monticuliporid genera. Two intact living chambers (Plate 8: figures 3, 4) protected by incrusting overgrowth have double and single funnel-cystiphragma in the same general position as those in living chambers (Plate 5: figures 4, 5; Plate 6) of the monticuliporids.
Calcareous skeleton Inner epidermis
Coelomic space
The general impression is that the layers of a basal diaphragm extend outward along the zooecial wall to form at least a partial lining of the living chamber. The outward extension of this diaphragm would not be as the lining of the living chamber, but as part of the adjacent cortex. The subsequent growth of the zooecial wall would necessarily have begun without a suitable chamber to hold the food organs.
In species of apertured trepostomes, it is common for the distance between successive apertures in the exozone of an autozooecium to be considerably less than the length of the living room at the end of the autozooecium. The outer position of the new basal diaphragm would then apparently be determined by the new outer position of the membranous organs. The relative size and inward slope of the recent heteroporid ligament (Plate 2: Figure 36) are similar to those of the halloporid spines.
In the several thin sections examined of this species, the spacing of the two structures appears to be similar. Their position suggests that they may have been part of the lophophore retraction mechanism. The inter-zooecial correlation of cystifragm placement shown in the figure is typical of the species (Ross 1967: pi.
A few plates extended from the hemiphragm to the wall to complete the separation from the living room and probably acted as basal structures. Perhaps the elongated, cylindrical shape characterized the membranous organs of the Late Paleozoic stenoporids, with or without ring septa. In a recent lichenoporid (Plate 2: Figures 1, 2), the lophophore is located on the proximal side of the autozooecium.
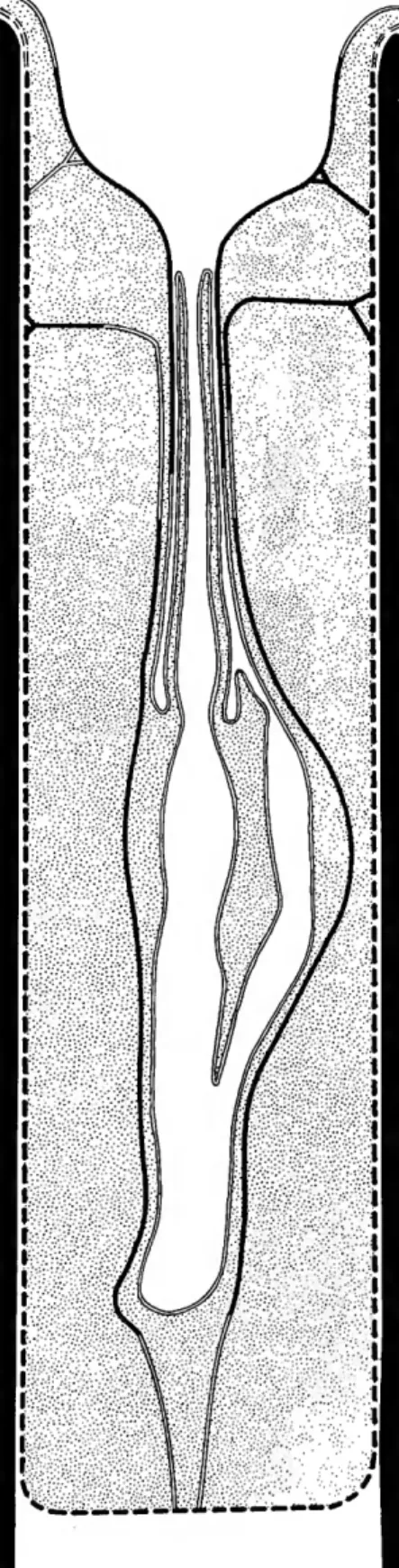
Attempts to correlate cycles would only be meaningful in the distal parts of ramose zoaria growing at the time of colony death. In the vast majority of flask-shaped chambers in Paleozoic trepostomes, funnel cystipragms have a skeletal microstructure and are thought to be formed by subparallel depositional epidermis of the inner membrane. It is believed that the occurrence of fossilized brown deposits indicates the presence of organic matter during the life of the colony.

KNOWN
COELOMIC SPACE
CALCAREOUS SKELETON INNER EPIDERMIS
UNKNOWN
INNER EPIDERMIS
OUTER MEMBRANE
The presence of cyclic patterns of chamber distribution in several species and the overwhelming number of species for which balloon-shaped chambers are not known argue against the accidental preservation of a common structure. It is assumed here that the general nature of the balloon-shaped distribution of skeletal chambers is approximated in the fossil record. No more than two or three lichenoporid tentacles may have passed through the neck in some of the heterothripids (Plate 8: figures 1-4, X 100).
The model in Figure 1 was created at the beginning of the study, when the flask-shaped chamber of a diaphragm trepostome looked as if it reflected the shape of enclosed feeding organs. The flask-shaped chamber in early Paleozoic monticuliporids and diaphragmatic repostomes seems best explained as part of the mechanism of sexual reproduction. A lophophore with few tentacles appears consistent with the shape and constriction of the funnel cystiphragms of the flask-shaped cham-.
If the chambers were parts of zooids, those zooids, regardless of their function, were polymorphic with the zooids of the consistently present and less restrictive chambers that alternated with the flask-shaped chambers in the same autozoa and that occurred in other autozooea throughout the zoarium. Regeneration from the male sexual stage to the asexual or feeding stage was subsequently reported in the same species (Powell. Cook, British Museum (Natural History), for her encouragement and help in the study of soft parts of recent cyclostomes and for arranging the loan of specimens from the British Museum.
Longitudinal thin section of a segment of an autozooid, X 150, showing the ends of the tentacles drawn back to level A of the lateral ligaments and a dark mass of U-shaped sphincter muscle cells (level B) in closed position. The terminal vestibular membrane is missing and the base of the tentacles is at the bottom of the figure. Note the membrane of the tentacular sheath, which passes upward through the opening in the sphincter muscle and appears to continue as the vestibular membrane.
The mesentery extends inwards and surrounds the long intestine in the lower left corner of the figure. Longitudinal thin section of segment of autozoid, X150, showing tentacles projecting outward past lateral ligaments (at level A) and through sphincter (at level B), greatly altering the shape of the profile of that muscle. Note double membranes of the tentacular sheath and membranous sac visible inwards from lateral ligaments.
31 PLATE 1
Longitudinal section, X 100, one funnel centered just out of plane of section at neck and complex of lateral membranous cystiphragms. Longitudinal section, USNM 167681, X100, shows a scalloped edge (arrow) of an incomplete cystiphragm due to the lateral position of the cystiphragm in the accommodation space. Longitudinal section, USNM 167686, X100, showing the flask-shaped chamber at the stage of development of the double funnel and cystiphragm.
Longitudinal section, X30, showing a more or less uniform cyclic distribution of the flask-shaped chambers in the zoarium. Longitudinal section, USNM 167684, X100, showing a complex of funnel-shaped cystiphragms that apparently developed without outward movement. Longitudinal section, USNM 167692, X30, showing concentration of brown deposits and incompletely formed chambers at level of zooecial wall thickening.
Longitudinal section, USNM 167695, XlOO, living chamber under protective overgrowth in a double funnel-cystiphragm stage of development. Longitudinal section, USNM 167696, XlOO, living chamber under protective overgrowth plus a covering diaphragm in a single funnel-cystiphragm stage of development. Longitudinal section, USNM 133901, XlOO, flask-shaped chamber filled with debris and covered at neck by subsequent skeletal growth.
Longitudinal section, USNM 167696, XlOO, two successive flask-shaped chambers, the outer of which shows extension of thin calcified chamber lining (arrow) beyond the normal autozooecial wall beneath a protective overgrowth. Longitudinal section, USNM 167702, XlOO, flask-shaped concentration of brown deposits bending around shelf-like hemifragments. Longitudinal section, USNM 167703, XlOO, flask-shaped concentration of brown deposit resting on the hemiphragma at the inner end and extended outward by a curved skeletal septum at the outer end.
Longitudinal section, X100, shows a funnel-shaped skeletal structure attached to the outer end of the brown deposit. Longitudinal section, USNM 167704, X100, showing a concentration of brown sediment that preserves an almost perfect flask shape, although it is indented by two hemiphragms. Longitudinal section, USNM 167705, X100, reddish brown membranous tube in transverse section passing through foramen of annular septum.
Longitudinal section, USNM 167708, Longitudinal section, USNM 167711, X50, showing the outward extent of living spaces with ring partitions under protective vegetation.