We argued that the receptive psychology paradigm could have a much greater impact on the study of animal signals and communication if it also encouraged research into the actual psychological mechanisms involved in communication and their potential evolutionary diversity. We are deeply grateful to all authors for their contributions to this volume and, more broadly, for their contribution to new discoveries about the psychology of signalers and receivers.
Sights and Sounds of Spring
They may also expose or exaggerate the epaulet and direct it at women or rival men using a number of other visual displays, such as the 'bill-up display', the 'defensive flutter' or a 'crouch' stance. such as different flight displays, such as 'flight song' and 'fluttering flight'. However, many unanswered questions remain about the psychological mechanisms underlying communication in animals such as the red-winged blackbird.

Psychological Mechanisms in Animal Communication
- Variation in Sensory Processing
- Signals and Signalers as Perceptual Objects
- Categorization and Social Cognition
- Referential Signals, Semantics, and Concepts
- Decision-Making
- Learning and Memory
In the second part of their chapter, Farris and Taylor (Chapter 4) address multicomponent signals and multisensory integration. Rubi and Stephens (Chapter 5) briefly discuss learning in the context of their economic model of multicomponent signaling.
Toward a Psychology of Signaling and Receiving
For example, Henry et al. Chapter 2) and Bee (Chapter 7) place their reviews of species differences in phylogenetic frameworks. Farris and Taylor (Chapter 4) and Page and Jones (Chapter 11) illustrate the largely untapped promise of robotics in investigating psychological mechanisms in animals.
Summary and Future Directions
Rubi and Stephens (ch. 5) show how theoretical modeling efforts can bridge the gap between studies of animal communication from psychological and behavioral ecology perspectives. Several chapters show how the workhorse of animal communication research - the credible playback study - can be used to investigate psychological mechanisms in field studies (Ch.. 6-9) and in highly controlled laboratory experiments (Ch.. Ch. 10) review the use of interactive playback experiments designed to to create virtual animals in the laboratory.
Introduction
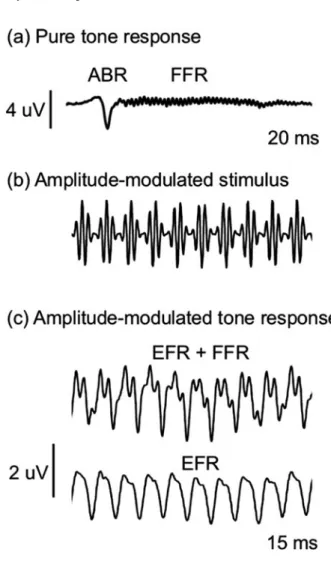
The spectral decomposition of sounds into their frequency components originates through auditory filtering in the inner ear and is maintained to varying degrees in different auditory nuclei along the entire length of the pathway from the brainstem nuclei to the midbrain, thalamus, and forebrain. (eg, Calford et al. 1983). The instantaneous firing rate of auditory neurons varies with the amplitude envelope of acoustic signals (following the envelope; Joris et al. 2004) and, at more peripheral levels of processing, also the fine structure of the pressure waveform (typically for frequencies less than 3 –6 kHz; following frequencies; Johnson1980).
Assessment of Hearing Function with AEPs
Differences in EFR amplitude at high modulation frequencies may reflect differences in the temporal resolution of the auditory system. ABR recovery functions in a variety of taxa, including birds, show a steady decrease in the amplitude of the.
Coevolution Between Signalers and Receivers
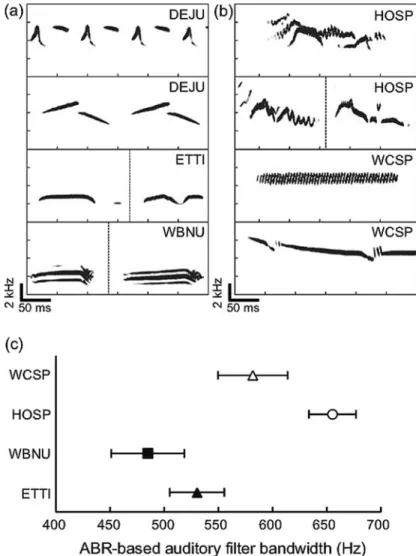
Song elements of brown-headed cowbirds (Molothrus ater) have the fastest rise times, followed by songs of dark-eyed juncos (Junco hyemalis), and, with the slowest rise times, song elements of white-cross sparrows, American tree sparrows (Spizzella arborea), and house finches (Carpodacus mexicanus). As predicted, FFRs for the sustained portion of the sound were poor in brown-headed cowbirds.
![Fig. 2.4 ABR-based audiograms of four songbird species (from Lucas et al. 2015 [Fig. 5]).](https://thumb-ap.123doks.com/thumbv2/123dok/11543257.0/36.659.164.499.533.777/fig-abr-based-audiograms-songbird-species-lucas-2015.webp)
Habitat Effects on Song and Hearing
In contrast, the FFRs for the different spectral components of the complex tones varied little across habitats. Thus, extended processing of individual spectral components and envelope fluctuations may be necessary.
Seasonal Auditory Plasticity
There are several possible mechanisms that may be responsible for plasticity in the songbird auditory periphery. This phenomenon has been seen in the auditory periphery of frogs (Gall and Wilczynski2015) and higher-order auditory processing areas in songbirds (Sockman et al and frogs (Gall and Wilczynski2014).
Sex Differences in Auditory Processing
In the same year, Gall and Lucas (2010) found that frequency resolution of brown-headed beebirds sampled during the breeding season also showed large differences between the sexes, with females having greater frequency resolution than males. During the breeding season, however, females had greater frequency resolution than males, but poorer temporal resolution.
Individual Variation in Auditory Physiology
Frequency sensitivity, when measured as the amplitude of the ABR to the onset of single-frequency sound stimulus, also tends to be greater in females than in males during the breeding season (Gall et al. 2012b). In this same species, for example, there are only small differences in ABR-based auditory thresholds, even during the breeding season (Ve´lez et al. 2015a).
AEP Responses to Natural Vocalizations
Similar to the WBNU song, the call note quank (Fig. 2.8e, f) can be characterized as a series of strong harmonics that produce a strong AM in both call notes (Table 2.1). The difference between WBNU quank and WBNU song elements is that WBNU has a higher AM level than quank calls (Table 2.1), and the gross amplitude envelope increases more slowly (compare Fig. 2.8c, d vs. Fig. 2.8e, f).

Summary and Future Directions
Henry KS, Lucas JR (2010a) Auditory sensitivity and frequency selectivity of auditory filters in the Carolina chickadee, Poecile carolinensis. Henry KS, Lucas JR (2010b) Habitat-related differences in frequency selectivity of auditory filters in songbirds.
Introduction
The Neuronal Substrate of Auditory Processing in Birds
The avian inferior colliculus in the midbrain receives signals from the left and right auditory pathways. Inferior colliculus neurons in the starling's close relative, the myna (Gracula religiosa intermedia), are tuned to modulation frequencies in the sound envelope, providing another cue representing sound sources (Hose et al. 1987).
Principles of Auditory Scene Analysis
While bottom-up processing in the starling brain has been investigated for many stimulus properties, top-down cognitive effects on processing in the auditory pathway have received less attention. The operation of such a heuristic is demonstrated when a deviation from regularity in a previous sequence or previous stimulus provokes the perception of a new object.
Auditory Scene Analysis and Signal Detection
Comodulation Masking Release
Similar to the band-spread paradigm, the masker envelope has greater spiciness in the correlated condition than in the alternative (uncorrelated) condition (cf. waveforms in Figure 3.2a,b). Panels (a) and (b) show the spectrograms (top) and waveforms (bottom) of the stimuli used in the unmodulated and comodulated conditions, respectively.
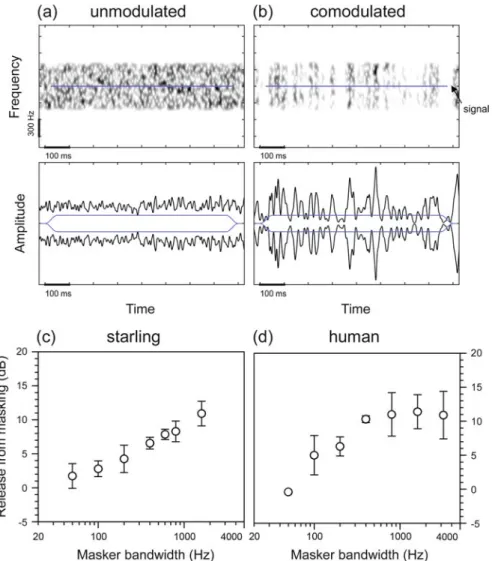
The Comodulation Detection Difference
Panels (d–f) show the magnitude of the CDD effect in response to the stimuli shown in (b) and (c) observed (d) in starling perception, (e) in the rate response of starling auditory forebrain neurons, and (f) as predicted by a model of the auditory periphery. Here, negative values indicate the relative improvement in thresholds in the co-uncorrelated condition, in which the envelopes of the flanking bands are correlated with each other, but uncorrelated with the envelope of the signal.
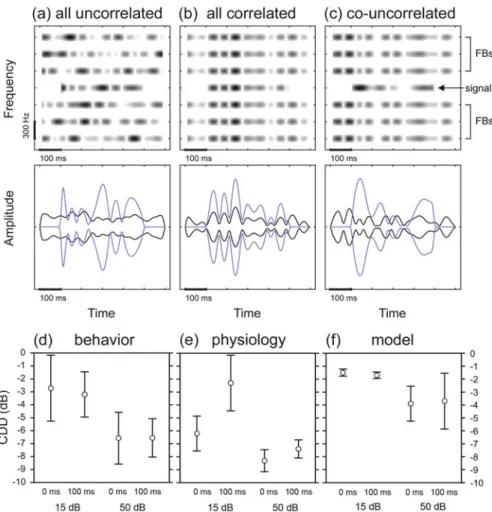
Relevance of CMR and CDD to Natural Listening Environments
Neural signal detection thresholds were about 2–8 dB lower in the co-correlated condition compared to the all-correlated condition (Fig. 3.3e). The laboratory studies reviewed in this section thus provide a representative account of the relevant effects for the perception of sounds in the natural environment.
Auditory Scene Analysis and Perceptual Restoration of Signals
For example, in human subjects, perceptual retrieval of words is enhanced if the words were previously known to the subjects. Schematic spectrograms of the stimuli used to train starlings to test perceptual recovery from pure tones are shown in (a) along with the reward contingencies.

Auditory Streaming of Sound Sequences
- Subjective and Objective Measures of Auditory Streaming in Humans
- Subjective and Objective Measures of Auditory Streaming in Starlings
- Neural Correlates of Auditory Streaming Based on Frequency Differences
- Auditory Streaming Based on Temporal Differences
Here, TRT corresponds to the time period from the onset of the first tone A to tone B in the normal time pattern;. Neurons in the auditory forebrain of the starling showed a descending response to continuous presentation of repeating ABA– triplets (Bee et al. 2010).

Summary and Future Directions
Bee MA, Klump GM (2005) Auditory stream segregation in the songbird forebrain: effects of temporal intervals on responses to interleaved tone sequences. Itatani N, Klump GM (2011) Neural correlates of auditory streaming of harmonic complex sounds with different phase relationships in the songbird forebrain.
Introduction
When the frequency difference is large enough, listeners report hearing two distinct streams, one of only A tones and one of only B tones, which is similar to when sequences of A or B tones are presented alone. Our focus in this chapter is on studies of perceptual grouping during mate choice primarily in anurans (frogs and toads) and, to a lesser extent, orthopterans (e.g. crickets).
Auditory Grouping
Auditory Grouping Cues
Based on Gestalt principles, Bregman (1990) described two categories of group signals: primitive and schema-based. But these Gestalt-based criteria for primitive and schema-based grouping are largely based on work with humans, leading to the question of whether (and to what extent) schema-based grouping can be innate versus learned (Bregman 1990).
Auditory Streaming
The frequency ratio required for perceptual splitting/fusion of two interleaved sequences is related to the frequency resolution of the auditory periphery (Rose and Moore2000). The acoustic structure of tu´ngara frog chirps and croaks allows for testing spectral influences on grouping for more complex stimuli.
Simultaneous Grouping
Cross-Modal Grouping
Anuran Multisensory Communication
Horizontal black rectangles represent the inflation-deflation cycle of the robotic frog vocal sac and its temporal relationship to the call in (d–j) (black, left). Thus, the frogs exhibit sequential cross-modal grouping for at least a short period of time after the call shift.

Beyond Bimodal Grouping
Males call as they float at the edge of the pond; the expansion of the vocal sac and subsequent abdominal movements necessarily generate ripples on the water surface (see Fig. 11.5 in Chapter 11). 2014) showed that male tu´ngara frogs make cross-modal comparisons by attending to these arrival time differences.
Summary and Future Directions
Gridi-Papp M, Rand AS, Ryan MJ (2006) Animal communication: complex call production in the tu'ngara frog. Knudsen EI (1982) Auditory and visual maps of space in the owl optic tectum.
Introduction
A large body of work shows that additional signaling components can increase the effectiveness of signals through a range of perceptual and cognitive mechanisms. Therefore, this is a surprising result, even considering the perceptual and cognitive benefits described above.
The Model
Alternative Actions, Uncertainty About Actions, Payoffs, and Signals
We first consider a “one-signal component” model (i.e., the flag model, McLinn and Stephens 2006 ; Dunlap and Stephens2009 ; McLinn and Stephens2010 ). Consistent with previous economic models, our results show that a single signal component is often sufficient.
Reliabilities
We exclude them for the sake of definition because if q is less than 0.5, we can simply relabel the signal states so that S becomes S+ and S+ becomes S, which has the effect of keeping q in the 0.5–1.0 range. We now define a similar variable representing the reliability of the second signal T such that P(T+| Good)¼P(T| Bad)¼r.
Preliminaries: When Should a Single Signal Be Followed?
We can see that at extreme values of p it is better to ignore the signal. Specifically, if it's small, it's better to ignore the signal and always reject, because the fact that it's small means that the bad condition is quite common and rejection is the best single action.
Following Two Signals
On the other hand, ifpis big it is better to ignore the signal and always accept because the fact thatpis big means that the good condition is quite common and acceptance is the best single action. However, at intermediate values of p, when conditions are neither predominantly good nor bad, it is better to use the signal.

Effects of Environmental Uncertainty and Signal Reliability
In the absence of a default action (ie whenp¼1/2) you should never use multiple signals, but instead follow the most reliable source of information. Specifically, when p>1/2, the animal should either follow the preponderance of negative evidence rule or always accept, depending on the precise level of p; when p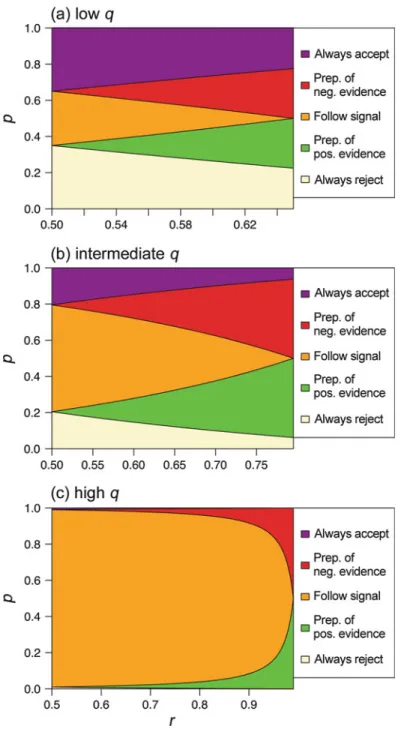
Environmental Uncertainty and Complex Signaling
Somewhat surprisingly, it is difficult to find evidence for "simple signals" in the existing literature. This example illustrates the danger of assuming that receivers will use all apparently important components of a signal.
Economic Benefits Versus Psychological Benefits
For example, Yorzinski et al. 2013) used telemetric gaze trackers to determine that peahens (Pavo cristatus) prioritize certain components of a peacock's courtship and ignore other, highly conspicuous components.
Limitations of this Approach
The consequences of errors will vary greatly between systems; a missed opportunity to mate is not the same as a missed opportunity to find food (Wiley 2015). However, there is good evidence that recipient choice may vary with internal factors, such as recipient need (Brower and Calvert1985; Skelhorn and Rowe2007).
Summary and Future Directions
Dunlap AS, Stephens DW (2009) Components of change in the development of learning and unlearned preference. Johnstone RA (1997) Recognition and the evolution of characteristic signatures: when does it pay to reveal identity.
Sensory Perception and Just-Noticeable Differences
In honey bee (Apis mellifera) dance communication, individuals rely on distance and direction information that is transformed multiple times between physical and perceived values. The dancer actually perceives physical distance and direction when he travels home from the food (Fig.6.1a).
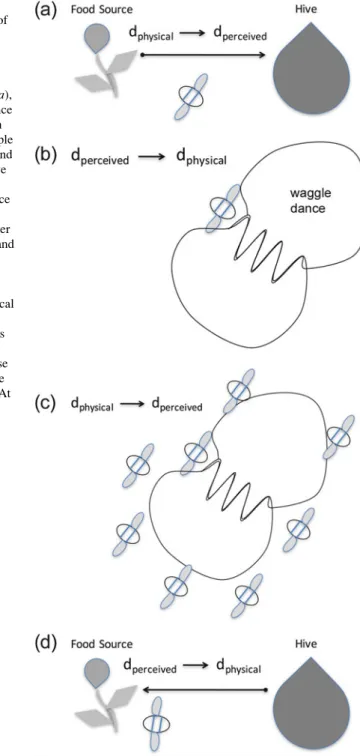
Proportional Processing and Receiver Error
In the remainder of this chapter, we discuss proportional treatment-induced evolutionary changes in communication systems. In section 6.3, we describe how proportional processing can affect the development of physical features of signals.
Impacts on Signals
- Halting the Evolution of Increasing Signal Magnitude
- New Signal Components
- Loss of Original Signal Components
- Directional Predictions
The cost of signal production determines whether proportional processing will limit the evolution of increasing signal size. When this happens, proportional processing by females may be the reason that evolution of increasing signal size stops.

Impacts on Signaler Strategies
Parallels to Evolutionary Consequences of Proportional Processing
Another parallel to evolutionary consequences of proportional processing is the facultative addition of different components to a display when benefits are most likely. Male Tu´ngara frogs (Physalaemus pustulosus) strategically decide when to switch between calls that vary in buckle number in ways expected for proportional treatment of female receivers.
Trade-offs Between Pushing High-Energy Signals and Maintaining Steady Mid-level Signals
Therefore, the distribution of perceived magnitudes for higher-magnitude stimuli is necessarily more variable as a result of proportional processing. According to the joint distribution of perceived stimulus magnitudes (bottom shaded area), animals would perceive the lower-magnitude cue as being more common than the higher-magnitude cue. From Bateson and Kacelnik1995, permission granted by John Wiley and Sons, Inc.).
Impacts on Receiver Responses
Attention to New Sensory Parameters or Signal Components
For example, emission signals are generally produced at magnitudes well above the contrast threshold, making them easy to detect (Cronin et al.2014). For example, females assessing a signal component indicating good genes may prefer expanded versions of this component, causing it to evolve to the point where females cannot distinguish between males.
Evolving a Compensatory Behavior
For example, a bee that perceives a returning forager dancing a dance (Fig. 6.1) may switch to a new strategy for estimating the duration of the movement when the food source being danced for is particularly far away, presenting the possibility of a the absolute biggest mistake. She could switch between estimating movement duration using visual, tactile, or acoustic information if the discrimination threshold for one parameter was better than that for the others.
Proportional Processing and Signal Detection Theory
When olfactory information is withheld from the navigation of honey bees that have observed another forager's dance, they behave exploratoryly and are usually unable to find the exact location of their target (Riley et al. 2005; Tautz and Sandeman 2003).
When JNDs Follow Other Patterns of Change with Variation in Stimulus Magnitude
In fact, proximity can alter phonotaxis behavior in anurans (Akre and Ryan2010) and insects (Mendelson and Shaw2006), and this psychophysical perspective indicates that females will place more weight on non-amplitude features of close-range decision making. However, birds exhibit a “near error” of Weber's law, with increased sensitivity to amplitude at short distances.
Summary and Future Directions
Akre KL, Farris HE, Lea AM, Page RA, Ryan MJ (2011) Signal perception in frogs and bats and the evolution of mating signals. Gerhardt HC (1994) Reproductive character displacement of female mate choice in the gray treefrog, Hyla chrysoscelis.
Introduction
The form of social recognition evidenced by the beloved-enemy effect has been particularly well documented in acoustic playback experiments with songbirds (Falls1982; Lambrechts and Dhondt1995; Stoddard1996). In Section 7.3, I critically review previous studies that have investigated neighbor recognition and the dear-enemy effect in anurans (see also Bee et al. 2016).
Components of Social Recognition Systems
Wiley (2013) described these issues in terms of “specificity” (how many individuals per category) and. Here, one distribution corresponds to differences in a particular signal property that occur between repeated signal deliveries by the same individual (intra-individual differences).
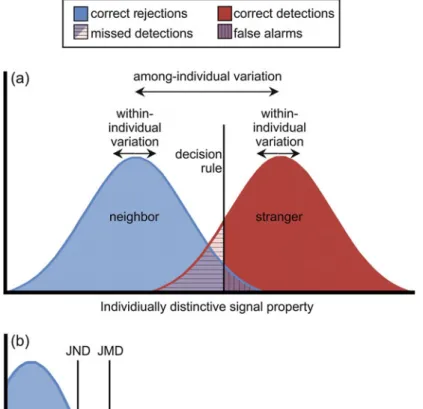
Neighbor Recognition and the Dear Enemy Effect in Anurans
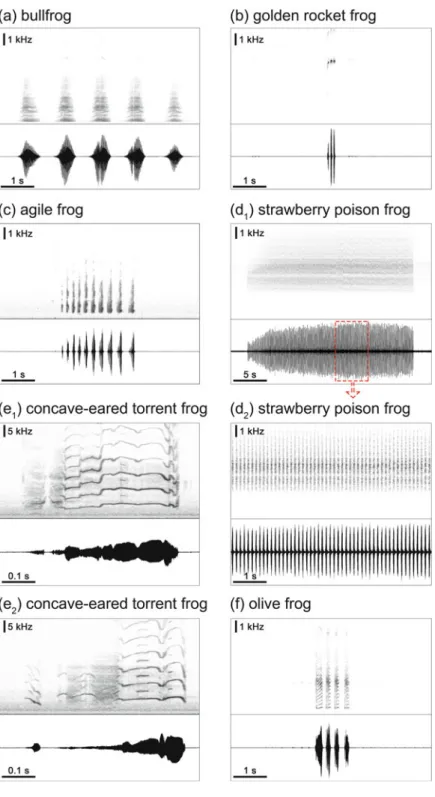
- Bullfrogs (Ranidae)
- Golden Rocket Frogs (Dendrobatidae)
- Agile Frogs (Ranidae)
- Strawberry Poison Frogs (Dendrobatidae)
- Concave-Eared Torrent Frogs (Ranidae)
Pettitt et al. 2012) published the first quantitative description of the vocal repertoire of this species. However, there was no evidence that subjects responded differently to playbacks of neighbors' and strangers' calls (Fig. 7.7a).