The classification of modern lower teleostean fishes has been set up piecemeal, and the interrelationships of many of the groups remain unknown. Often the nature of the change which takes place between the basal members of different orders is of the same kind as that which takes place between the more "primitive" and more. The present paper deals with the caudal skeleton in some of the orders commonly placed between the isospondylous fishes (Clupeiformes) and the percomorph fishes (Perciformes).
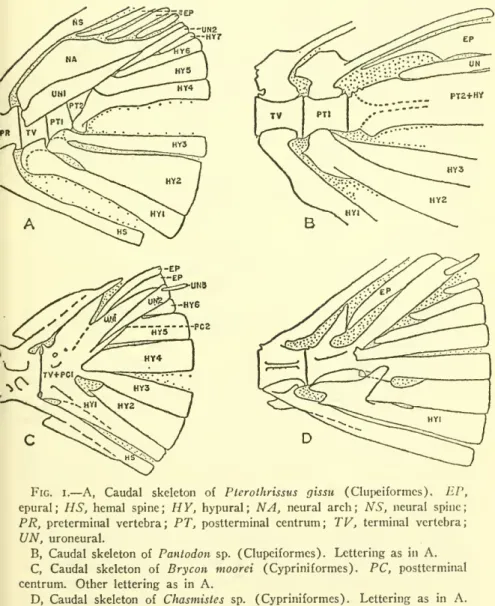
All tail structures to be discussed parallel one of the three types evolved in isospondylous fish. At this point, the three lower hypurals all attach to the terminal vertebra, leaving only some of the upper hypurals to articulate with the remaining postterminal center. Here one or more uroneurals fuse with the terminal vertebra, so that the upward-facing support, also called urostyle in these fish, is actually a composite structure consisting largely of the uroneurals.
Although the basic aim of this article is to detect morphological similarities in the caudal skeleton as possible indicators of phylogenetic evolution, a secondary aim is to examine certain aspects of the relationship between skeletal structure and fin shape. As noted in the previous article, there appears to be an indirect relationship between tail shape, number of tail rays and structure of the tail skeleton.
NO. 3 TELEOSTEAN FISHES — GOSLINE
8 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
An attempt was made to get at this problem by comparing the cyprinids (17 branched rays) with the catostomids.
10 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE II
12 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
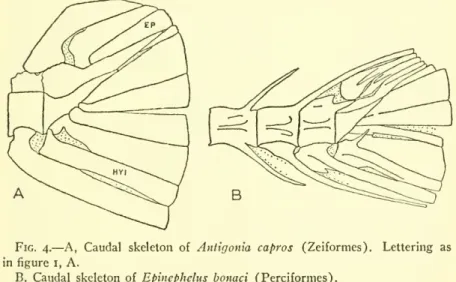
3 TELEOSTEAN FISHES— GOSLINE 13Orders Beryciformes, Zeiformes, and Perciformes.— As to the orders Beryciformes, Zeiformes, and Perciformes.— As to. There seems no reason why one should not be derived from the base stock of the other or, alternatively,. In each of the three orders in the caudal skeleton there is a degree of forms in which this structure is present.

14 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES GOSLINE 15
This secondary simplification of structure developed through a number of steps, and these sequences of these steps appear to have been essentially the same in different groups of fish. Thus, hypurals 2 and 3 appear to fuse before hypural i is added to the group, e.g., in cyprinodonts and apparently in eels; The postterminal centrum joins the terminal vertebra, with postterminal centrum 2 occurring in more advanced forms. When these sequences are altered, other features of the tail skeleton appear to evolve differently from the usual way, or vice versa.
In any case, the different sequences form the basis on which the three tail skeleton types mentioned here are established. In a very general way, there is a connection between the fusion of parts in the tail skeleton and the reduction in the number of tail rays. Perhaps no other meaning should be taken in this than as independent examples of the trend in the evolution of teleostean fishes towards the reduction of the number of parts of the head and body.
In any case, in the case of the cyprinoids discussed above, there was no correlation between the number of caudal rays and hypurals. There also appears to be little direct correlation between .. amount of fusion and fin type or lifestyle. There are numerous round-tailed fishes with a fused, plate-like caudal skeleton (Fundulus) and fork-tailed forms with similar features (Beloniformes); similarly there are slow and weakly moving forms (tube fish) and fast swimming fish (tuna) with a flat tail skeleton.
Returning to the three basic types of caudal structure distinguished at the beginning of this section. In these the uroneural(s) are not fused to the vertebral elements, and a second postterminal centrum is usually visible, although the postterminal centrum is fused to the terminal vertebra. Indeed, Type I appears to be a stage in the normal or at least common path of evolution followed in modern teleosts.
NO. 3 TELEOSTEAN FISHES — GOSLINE 1
SOME FEATURES OF PELVIC STRUCTURE
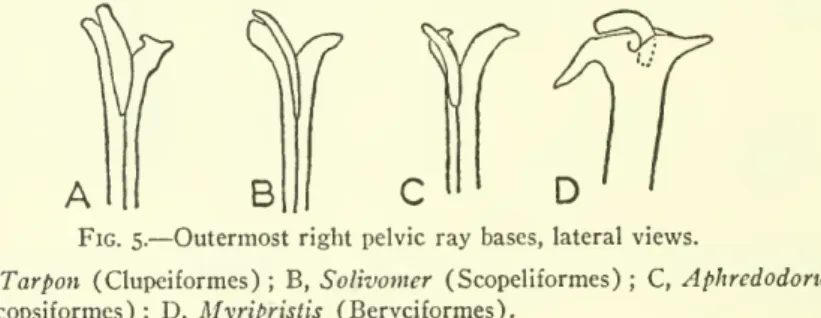
The facets of pelvic structure that will be addressed here are the small, curved bony rail that lies outside the outermost ray in many lower teleosts, and the radial elements. The pelvic fins of most lower teleosts consist of a series of segmented soft rays (lepidotrichia of Goodrich, 1904, Jarvik, 1959 and others), each containing a separate upper and lower half. The only exception is the curved rail that runs for most of its length along the outer surface of the upper half of the outer beam on each side (Figs. 5A to C).
In Lepisosteus there is a small, unpaired, diamond-shaped plate on the outside of the base of the most lateral ray, but whether this is the same element as the curved bar of Amias seems open to question.
NO. 3 TELEOSTEAN FISHES — GOSLINE I9
20 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
SOME BONES OF THE SNOUT REGION IN MODERN TELEOSTEAN FISHES
The bones discussed in this section are the supraorbital bones and certain ossifications that surround the sensory tracts of the brain.
22 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
3 TELEOSTEAN FISHES- GOSLINE 23written on eel by Allis (1903); this will not be discussed here. At least some of the difficulties with the cranial canal bones in Albula and Pterothrissus undoubtedly arise in connection with the cave system of the lateral line canals of these fishes. Many deep-water fishes, but also some shallow-water and even some freshwater forms, have such a system of spoons that replace the usual sensory channels of the head.
First of all, the outer surface of these troughs is covered with a layer of thin, taut skin. Alternatively, such a system may have a large number of small openings on the exterior, but lack the usual array of large (primary) sensory channel pores. Connections can be established between some parts of the cavernous system, and membrane barriers can develop between others.
In Albula, the supraorbital canal on each side passes forward over a nasal and a prenasal ossicle to end blindly at the edge of the snout. Whether this cavity represents Elop's ethmoidal commissure, the peculiar median sensory system of eels described by Allis (1903), or whether it is. In the related Pterothrissus (Fig. 7) the supraorbital canal extends to the tip of the snout, down which it extends for a short distance before ending blindly.
Near its anterior end it is in open connection with its relative on the other side through a hole through the mesethmoid; at the bottom of this transverse opening is a well-developed neuromast.
24 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
One, represented in the fishes studied only of the Beloniformes, has no nostrils and no olfactory laminae, at Another type of nasal structure consists of a series of transverse laminae that lie at the base of a nasal capsule that is closed above except for two rather small, well-separated nostrils. Water is presumably conducted across the nasal epithelium by such an olfactory organ by ciliary action as in Anguilla (Liermann, 1933).
In most living teleosts, by contrast, there are two relatively large, adjacent nasal openings leading to the olfactory capsule, at the bottom of which lie the olfactory layers. The movement of bones around these sacs alternately contracts and expands, thus pumping water in and out across the nasal epithelium (Eaton, 1956). It is with some of the bones involved in this pumping system that the present discussion will deal.
The antorbital (Fig. 7B, AN) is a bone quite commonly represented among the lower leg bones, but apparently incorporated into the lacrimal bone in higher forms. It undoubtedly originated as a sensory canal bone, but in living lower teleosts it serves primarily as part of the nose. Westol, 1937, p. 519)- However, it seems likely that the antorbital of Amia is equivalent to the two lateral rostrals plus the antorbital of Elops (cf. teleosts, the antorbital is associated with the supraorbital bone. Fig. 7B) above it and reduces or loses its sensory canal .
26 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES GOSLINE 27
28 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE 29
30 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE 31
UPPER JAW PROTRUSION IN TELEOSTEAN FISHES The freeing of the maxillary from the cheek is generally considered
32 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
3 TELEOSTEAN FISHES— GOSLINE 33Macruriformes, Gasterosteiformes, Lampridiformes, most Cyprino-Macruriformes, Gasterosteiformes, Lampridiformes, most Cyprino-dontiformes, Phallostethiformes, Stephanoberyci formes, Beryci-formes, Mugiliformes and most higher orders of teleosts. The protrusile maxilla of cyprinoid fishes has been discussed several times (cf. Fiebiger, 1931; Gregory, 1933); its structure is very different from that of percoid fishes and certainly represents an independent development. Regarding the structural elements mentioned in the description of the percoid jaw, there appears to be a basic difference between the projecting upper jaws of cods, sticklebacks, holocentrids, and phalostetids and those of percoids.
But whether the peculiar type of jaw projection found in the Lampridiformes (Regan, 1907) is of basic percoid type or has been independently derived. The distinctive characteristics of mugilid jaw structure can be traced, via the atherinids and sphyraenids, directly back to the percoid type. Among cyprinodont genera with a non-protruding premaxilla, Oryzias has the same descending process at the tip of the premaxilla as Foundtdus and the others mentioned.
34 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
DISCUSSION
In connection with this structural difference, the supraorbital-antorbital and lacrimal pumping mechanisms so characteristic of the Clupei are never present in the Osteoglossa, in which the area normally occupied by these bones is filled with different series of bones, which are rather difficult to explain. Certainly some members of the Clupei have lost their nasal diverticula, e.g. Esox and Umbra, but in them the bones of the snout never progressed as divergently as in Osteoglossa. In the same paper (Gosline, i960), elopids and albulids were placed together in the suborder Elopoidei of the section Clupei.
The presence of sensory canals in the premaxillaries of albulids, unique among recent teleosts, once again points to the divergence between these two groups. Scopeliformes.—The presence of fulcral scales and of well-developed temporal fossae in the posterior part of the skull of the basal iniomous genus Aulopus makes it impossible to derive the Scopeliformes from anything higher in the scale of modern teleosts than the elopoid Clupeiformes. The close resemblance of the caudal skeleton of these fishes to that of the osteoglossoids (Gosline, i960) confirms the interrelationship of these groups assumed on other grounds.
Cypriniformes.—The rather remarkable resemblance between the tail skeleton of the character Bryeon and that of the round herrings has been noted in Part I of this article.
36 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE 37 In conclusion it seems relevant to discuss groupings of modern
38 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE 39
40 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
NO. 3 TELEOSTEAN FISHES — GOSLINE 4I
42 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. I42
WOHLFAHRT, T. A