Introduction
Swimming at low Reynolds number
The linearity and time independence of the Stokes equations (1.2) and (1.3) lead to kinematic reversibility, which is a fundamental limitation associated with motions of bodies at zero Reynolds number. The squirmer model consists of a body that has a prescribed surface activation described by the velocity field u𝑠(𝑡) - the swimming stroke.
Active colloids
- The active Brownian particle model
- Pressure in active colloids
In other words, the instantaneous velocity U =𝑈𝑠q: 𝑈𝑠 is the swimming speed and q is a unit vector in the swimming direction. Therefore, the osmotic pressure at the wall is the sum of the osmotic pressure and the swimming pressure far away in the bulk.
Thesis outline
- Transport of active colloids
- Microrheology of active colloids
- Activity-induced propulsion
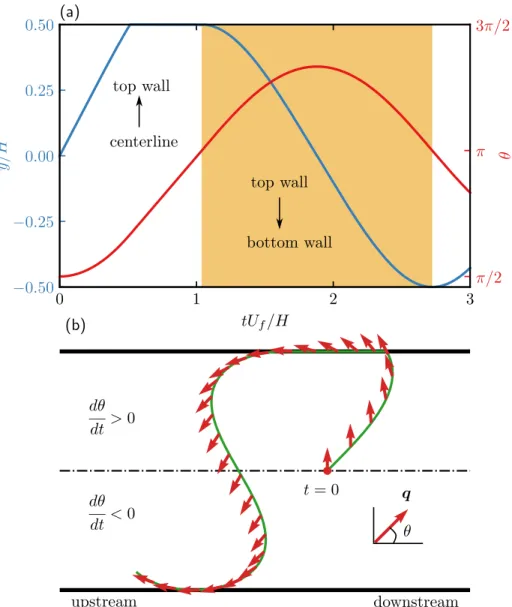
Particles rotate counterclockwise (𝑑𝜃/𝑑 𝑡 >0) in the upper half of the channel and clockwise (𝑑𝜃/𝑑 𝑡 < 0) in the lower half of the channel. When the probe perturbation flow (shown in blue) is included, the density (corresponding to the radial polar order) at the back (𝜃 =±𝜋) becomes higher than that at the front.
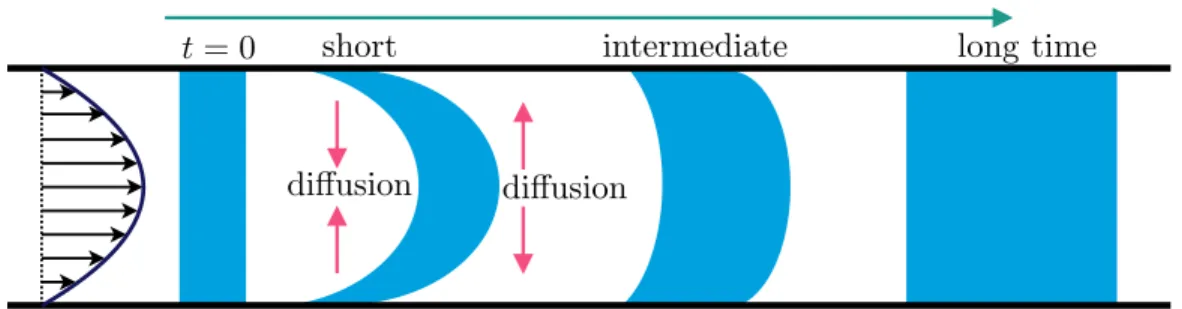
Upstream swimming and Taylor dispersion
Introduction
Matilla et al.(2019) in the context of ABPs in external flow through periodic porous media. The net velocity of ABPs in the longitudinal direction approaches that of passive Brownian particles in the strong flow limit.
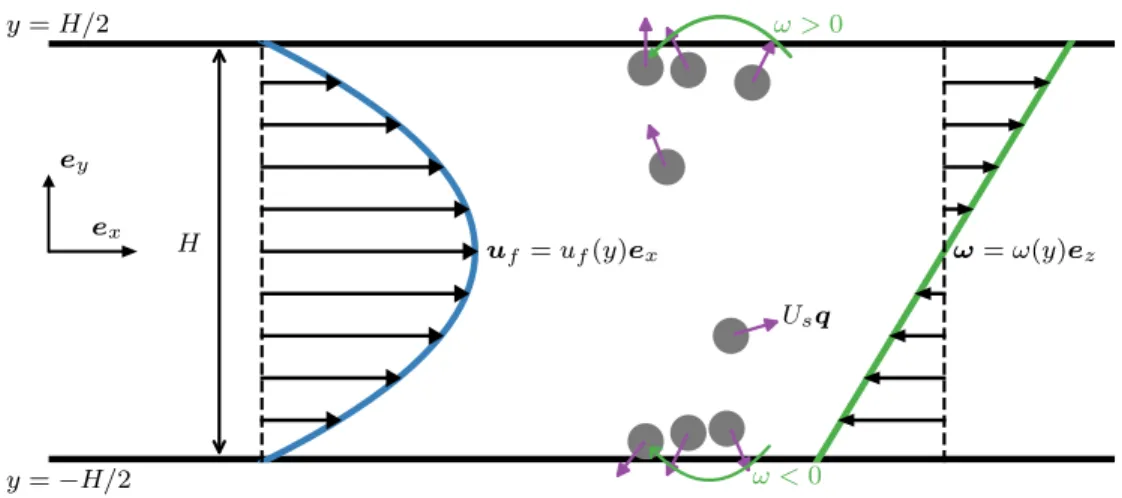
Problem formulation
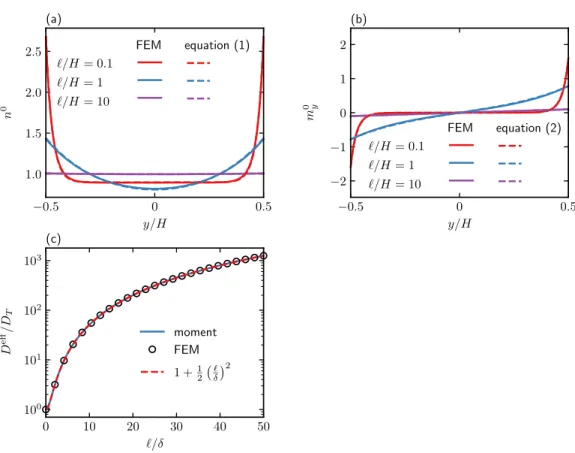
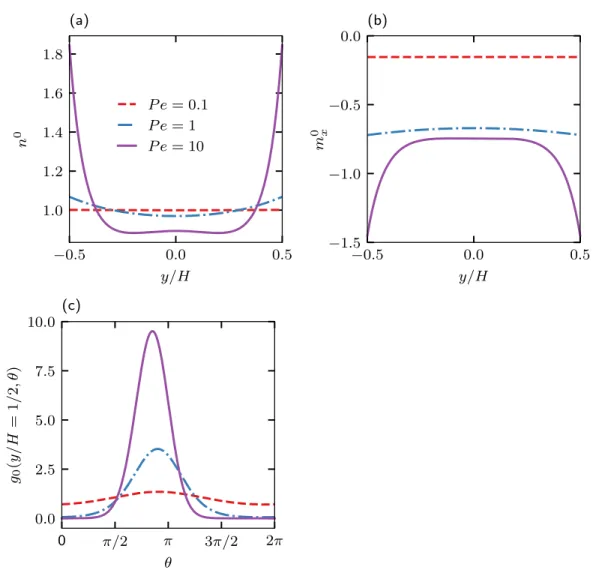
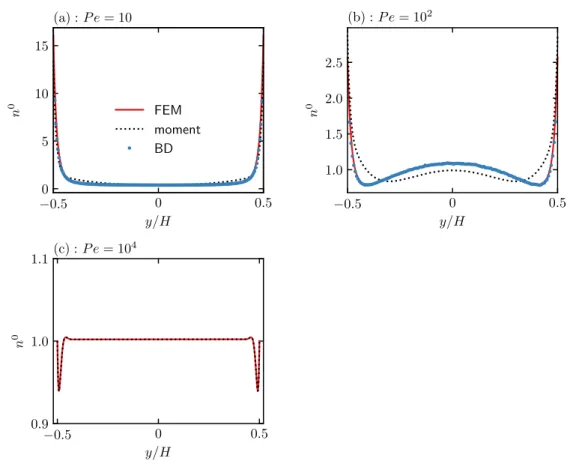
The governing equations are characterized by three dimensionless groups: closing force ℓ/𝐻 = 𝑈𝑠𝜏𝑅/𝐻, activity level ℓ/𝛿 = 𝑈𝑠𝜏𝑅/√. The Péclet number characterizes the relative importance of advection by flow and translational diffusion. We note that the qualitative behavior of the mean displacement and longitudinal distribution does not depend on the dimensionality of the orientation space.
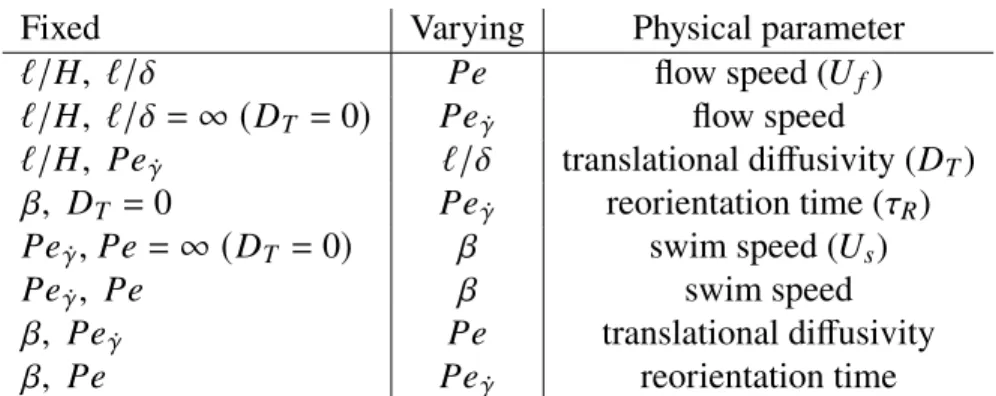
Upstream swimming
- Non-Brownian active particles
- No translational diffusion
- Finite translational and rotational diffusion
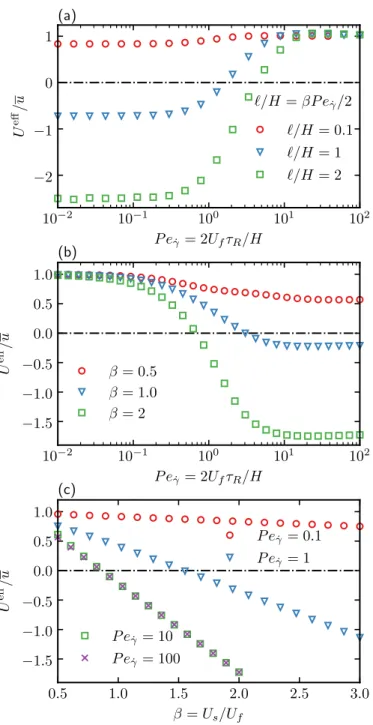
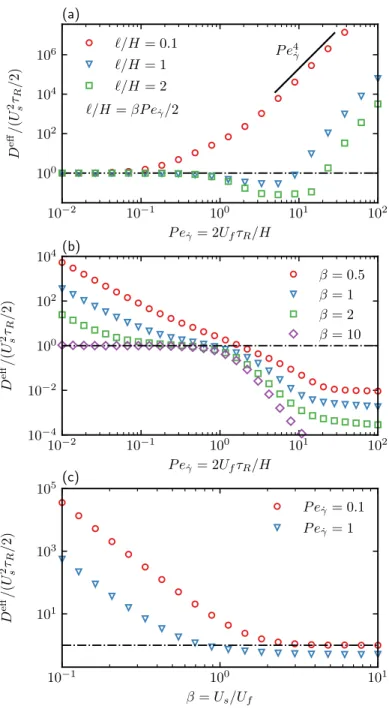
In Figure 2.7(b) we show the average drift 𝑈eff/𝑢 as a function of 𝑃𝑒𝛾¤ for different values of the velocity ratio 𝛽 in the absence of translational diffusion. In Figure 2.8(c), the average field 𝑔0 at the top wall is plotted as a function of the orientation angle 𝜃 for different Péclet numbers.
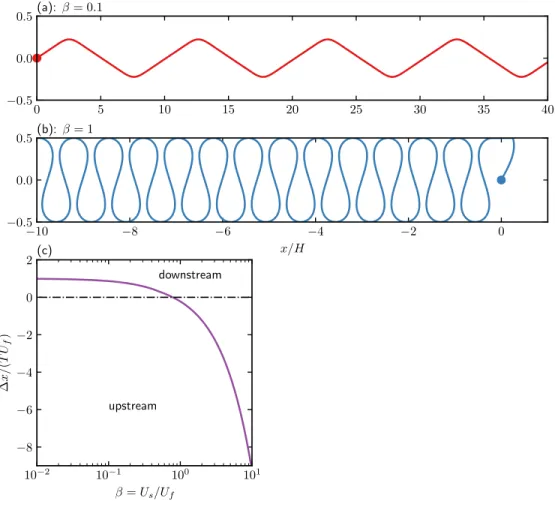
Longitudinal dispersion
- Dispersion in the absence of translational diffusion
- Dispersion with finite translational diffusion
In the classical Taylor dispersion problem, the effective dispersion increases monotonically as a function of the Péclet number. The strong non-monotonic variation of the effective diffusion coefficient as a function of the flow rate (𝑃𝑒𝛾¤) is observed when the translational diffusion is weak (ℓ/𝛿 is large).
Conclusion
As a result, the spherical ABP model provides a basis for understanding the transport of active particles in channel flow. The effect of the alignment is to improve upstream swimming when the flow is weak.
Appendix: The orientational moments
Equation (2.30) together with its boundary condition (2.35) is analytically integrated once to obtain a first-order equation. To solve the average filed moment equations and (2.51), we start from a guess for 𝑛0(−𝐻/2) as a boundary condition and update the guess based on a root finding algorithm so that the normalization condition is satisfied within a tolerance from 10-6.
Appendix: Brownian dynamics simulation
In most simulations, we use a time stepΔ𝑡 =10−3𝑡minku The simulations were run for a duration of 100𝑡maxwu𝑡max is the largest time scale in the problem.
Appendix: Non-spherical particles
The second dimensionless group is the Péclet number of the probe (using the diffusivity of the ABP). When the probe velocity is zero, that is, 𝑃𝑒 = 0, the microstructure (hence the density) is isotropic, which is simply the distribution of ABPs outside a solid sphere (Yan and Brady 2015b).
Dynamics of an active ellipsoid in a Poiseuille flow
Introduction
Disregarding hydrodynamic interactions between the spheroid and the channel walls, the orientational dynamics of a prolate spheroid in pressure-driven flow is often modeled by Jeffery's equation ( Jeffery 1922 ). Compared to a spherical particle rotating with the local angular velocity of the flow, the spheroid experiences the additional effect of alignment from the stress velocity tensor.
Problem formulation
- Constrained equation of motion in 3D
- Constrained equation of motion in 2D
- Time discretization
In particular, the last term represents the Brownian drift due to the gradient of the mobility tensor. Collisions of the particle with a wall are assumed to be inelastic; the particle and the wall can remain in contact with each other after collisions.
Deterministic dynamics
At long times, the particle exhibits a nonzero orientation angle determined by the balance of torque due to flow and contact torque. The ellipsoid then rotates towards the upstream direction and is able to escape into most of the channel.
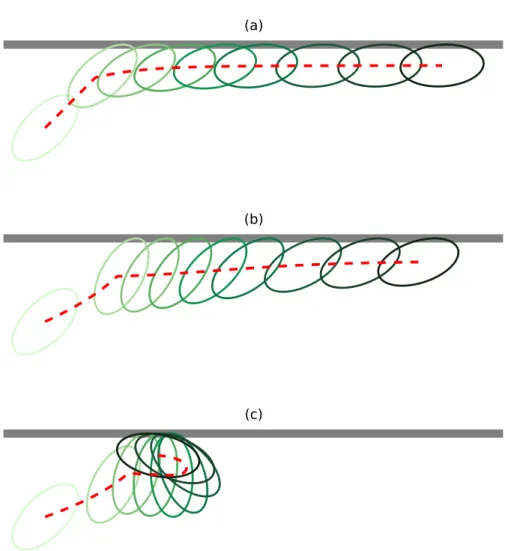
Effective transport
In Figure 3.5, we present the dimensionless effective longitudinal dispersion𝐷eff/𝐷swøm as a function of𝑈𝑓/𝑈𝑠 for different particle shapes. For spherical/point particles, in the small flow limit, the scattering is due to activity and given by the swimming diffusivity𝐷swim =𝑈2.
Appendix: Minimum separation between a spheroid and a planar wall 69
The external vector that is normal to the spheroid surface is in the direction of the vector R𝚲R⊺(y−x). More importantly, the limitation of CF or CV presents a difficulty in meaningfully quantifying probe fluctuations due to a thermodynamic uncertainty bound.
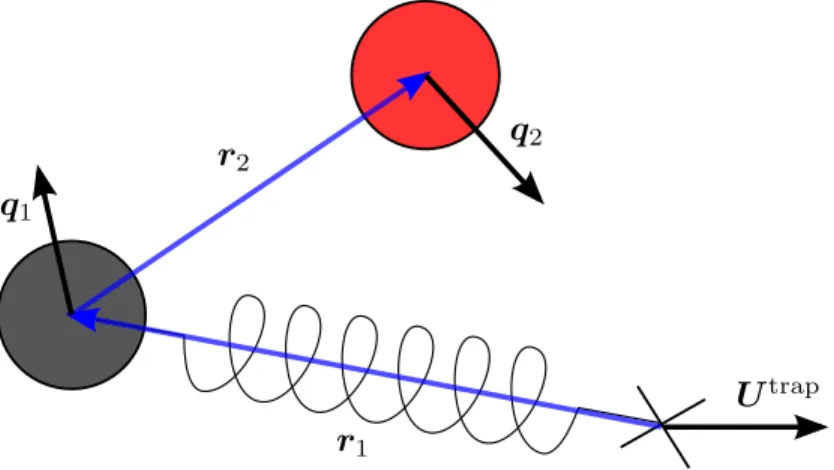
Introduction
In CF mode, the probe is driven by a constant external force Fext and the probe speed fluctuates. For CV mode, the position of the probe is also prescribed and the force fluctuation is infinite.
Mechanics of active Brownian suspensions
The trap force fluctuations and the position from the trap center satisfy an identical relationship (Wang 2017). Equation (4.2) also implies that we must consider the position rather than the velocity of the probe.
Moving trap microrheology
- Mean and fluctuation of the probe position
- The pair problem
- The probe distribution in the absence of bath particles
- A weak trap
- A strong trap
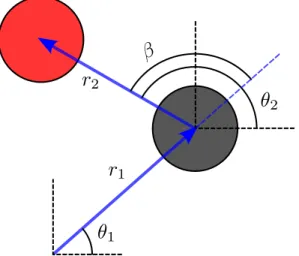
The average position or average displacement of the probe relative to the trap is defined by. The lowest level problem in the above formulation is the single particle problem of the probe interacting with the trap.
Constant-force and constant-velocity microrheology
- Constant-force microrheology
- Force-induced tracer dispersion
- Constant-velocity microrheology
2 = 0 and by integrating out the orientational degrees of freedom of the probe and the bath particle, we obtain the CF microrheology problem of a passive Brownian probe in a passive Brownian suspension. 2 =0 and by integrating the orientational degrees of freedom of both the probe and the bath particles, we recover the equations governing the force-induced spreading of a passive probe in a passive suspension (Zia and Brady 2010).
Fluctuations in a passive suspension
The CV microrheology of an active Brownian suspension governed by (4.80) and (4.81) has been studied by Burkholder and Brady (2020). The pair distribution determined by (4.82) still has high dimensionality even without considering the orientation degrees of freedom, as is the case for a passive probe in a passive suspension.
Derivation of the pair problem
In the dilute limit, we neglect the terms involving the gradients of ln𝑃𝑁−2/2 and use the pair mobility tensor in the absence of other particles instead of ⟨M⟩𝑁−2/2.
Derivation of the variance relations
It can be seen that the net polar order of the probe is not affected by the trap or bath particles. Similar to ⟨q1⟩, the nematic order of the probe regardless of the presence of trap or bath particles is given by.
Asymptotic analysis of the probe in the absence of bath particles
It is clear that in the weak trap limit at this intermediate time scale, the ABP exhibits diffusive behavior with the free space dispersion𝐷𝑇. As long as the trap strength is not identically zero, ABP will eventually (𝑡/𝜏𝑘 ≫ 1) experience the confinement of the trap.
Solution for a passive probe in a passive suspension
A weak density gradient is present in most of the inland areas due to the variation in swimming speed. In the leading order, the probability density in most of the interior is determined by.
Constant-velocity microrheology
Introduction
This is done in the dilute limit, where only the interactions between the probe and one of the bath's ABPs matter. For ABPs in the presence of the probe disturbance current, the force thickening originates from the interaction between swimming and the disturbance current.
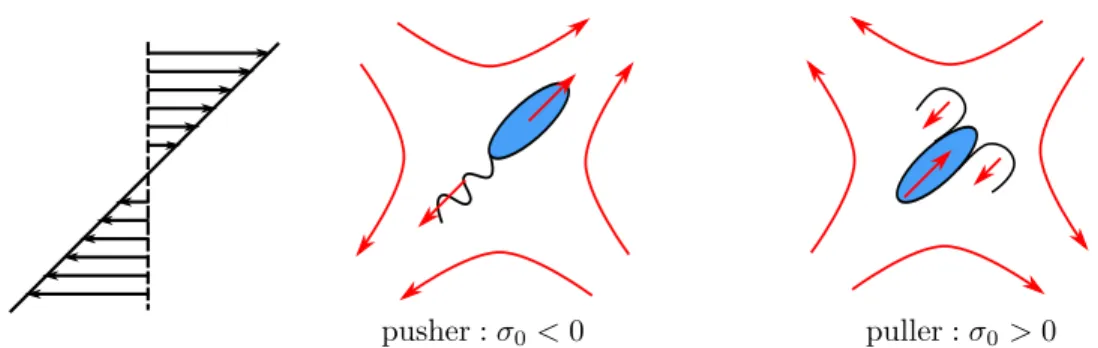
Problem formulation
- The Smoluchowski equation in 2D
Due to the fluctuating nature of the ABP suspension, the external force averaged over Brownian fluctuations is taken into account. Because the bath particles are active, the phase space of the microstructure includes both the relative position r and the orientation q.
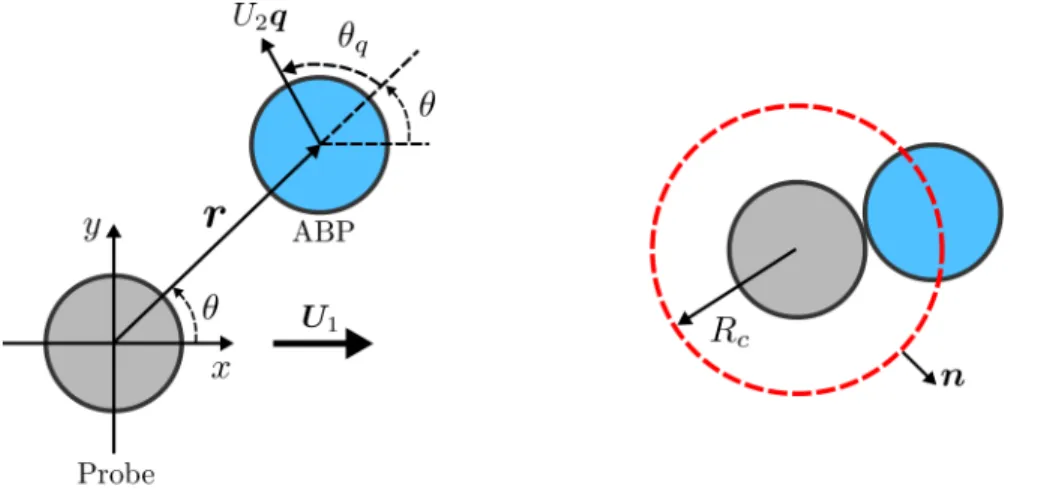
A slow probe
- Perturbation expansion of the microstructure
- Fast-swimming ABPs
- Zero-forcing microviscosity
We now consider the suspension microstructure and the zero-force microviscosity in the fast swimming limit characterized by
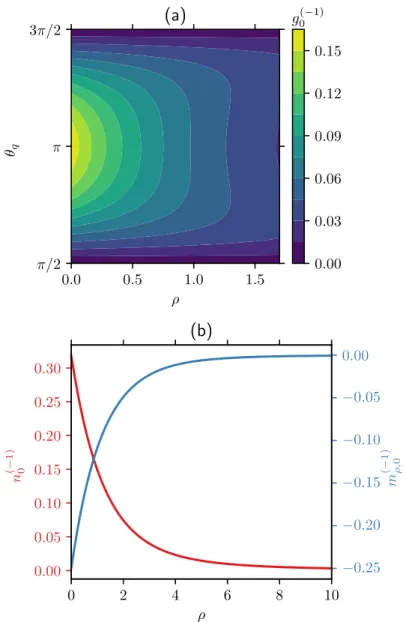
Numerical solution of the Smoluchowski equation
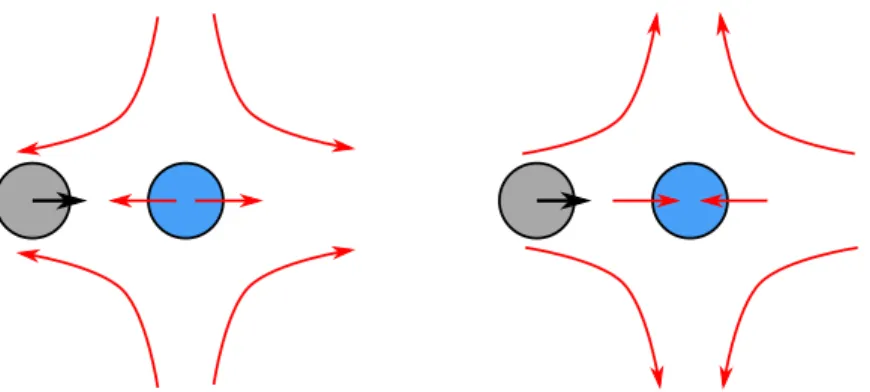
As 𝑃𝑒 increases, a prominently non-uniform density distribution develops at contact with an accumulation at the front and a depletion in the rear of the probe as can be seen in figure 5.8(a). We especially note that the contact density is increased on all sides of the probe.
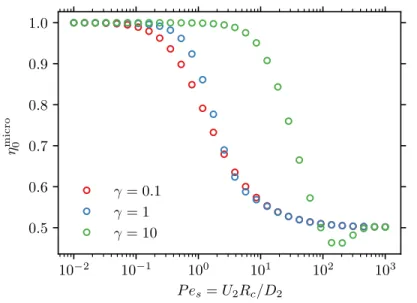
Brownian dynamics simulation
To understand the development of offspring, consider an ABP that is behind the probe (on the left). When the speed of the probe is much larger than that of the ABPs, 𝑈1 ≫ 𝑈2, the trailing wake is quite extended in the horizontal direction.
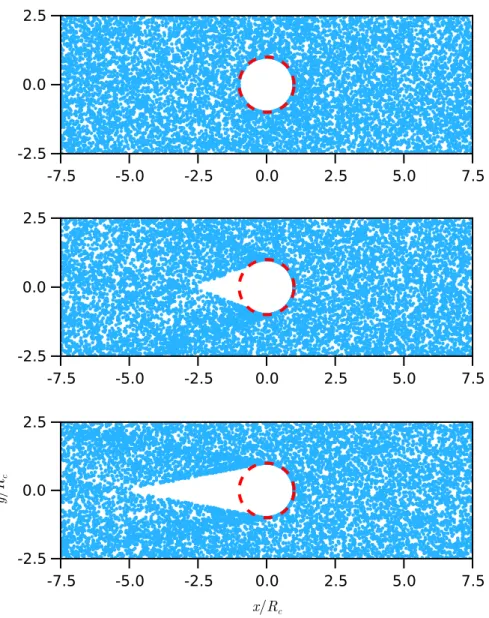
Microviscosity
Effects of hydrodynamics
- Fluid disturbance due to the active force dipole
- Fluid disturbance due to the probe motion
ABPs in Stokes probe flow experience spatially varying fluid advection and rotate with eddy. For comparison, previously obtained results in the absence of probe disturbance flow are also plotted (see Figure 5.11).
Concluding remarks
The behavior is reversed in the absence of probe interference, with more ABP pushing anteriorly than posteriorly. In conclusion, we have shown that in the absence of hydrodynamic interactions, the microviscosity of active suspensions is always positive.
Appendix: Orientational moments for a slow probe
It is also important to note that the negative shear viscosity depends on the non-spherical shape of the active particle. This is a manifestation of the so-called linear response where the disturbance fields are proportional to the vector U1 – the weak driving force.
Appendix: The slow-swimming limit
The average density of ABPs inside the vesicle is 𝑛 = 𝑁/𝑉𝑖, where 𝑉𝑖 = 4𝜋(𝑅− ℓ𝑚)3/3 is the volume of the interior. At the inner surface of the vesicle (𝑦 = 0), the leading-order density is large and given by 𝛾2.
Microviscoelasticity
Introduction
In the previous chapter, we studied the microviscous response of active colloidal suspensions. From a theoretical perspective, Khair and Brady (2005) and Swan et al. (2014) discussed the oscillatory microrheology of passive colloidal suspensions.
Theoretical framework
In the absence of hydrodynamic interactions between the probe and the ABP, the translational and rotational fluxes are in the Smoluchowski equation (6.2), respectively. To characterize the microviscoelastic response of the suspension in the dilute limit, a microviscosity can be defined.

Small amplitude oscillations
- The governing equations in 2D
- The high-frequency limit
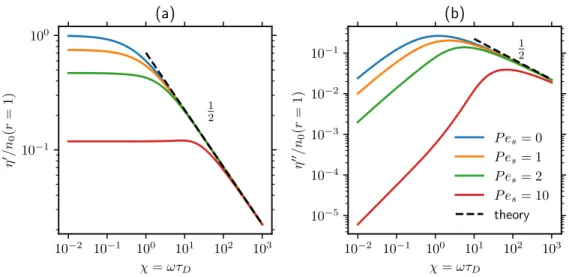
Because the swimming motion is eclipsed by a fast probe, swimming thinning is most important in the slow probe regime and disappears in the fast probe limit. For an oscillating probe, as we consider in this chapter, the swimming motion is likewise obscured in the fast-probe limit and the microviscoelastic response becomes the same as that of passive suspensions.
The zero-forcing microviscoelasticity
Here, as seen in (6.24), ℎ is driven by the radial advective flux of the 𝑔0 field at contact. In the large𝜒 limit, both the viscous and the elastic part of the microviscosity decay as 𝜒−1/2 [see equation (6.27)].
Appendix: Numerical solution to 𝑓 1 and 𝑓 2
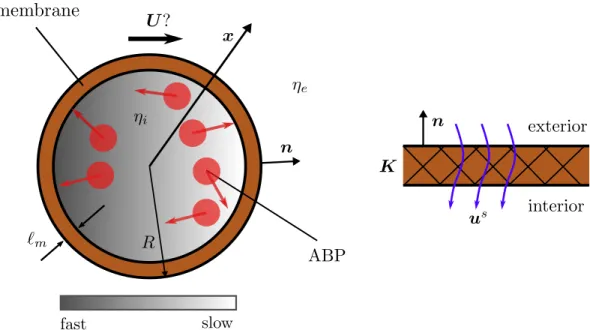
In this limit, the translational velocity of the vesicle is linearly proportional to the driving force - the osmotic pressure. Particles and solvent inside the bubble are treated as a continuum and manipulated.
Activity-induced propulsion of a vesicle
Introduction
Of more interest is whether the bubble moves in the same or opposite direction of the concentration gradient. At this high activity limit, ABPs have a thin accumulation boundary layer on the inner surface of the vesicle.
Problem formulation
- The exterior flow
- The interior suspension
- Dynamics of ABPs
- Transport in the membrane
- Dynamics of the vesicle
- Non-dimensional equations for a spherical vesicle
At the inner surface of the vesicle, the flux must vanish relative to the rigid-body motion. Here ℓ𝑚 is the thickness of the membrane and the thin membrane state is ℓ𝑚 ≪ 𝑅 with 𝑅 being the radius of the outer surface.
Vesicle motion in the limit of weak interior flow
- Governing equations
- High activity
- A large vesicle
- Vesicle motion due to an external orienting field
This boundary layer structure allows us to relate the osmotic pressure at the inner surface of the vesicle to the swimming pressure outside the boundary layer. The variation of the swimming speed leads to a gradient in the number density in the bulk of the interior.
Slow variation in activity
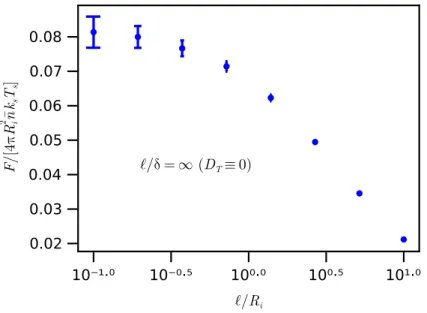
7.4(a), we show the dimensionless net force exerted on the inner wall by the ABPs, F𝑤/(4𝜋 𝑅2 . 𝑖𝑛 𝑘𝑠𝑇𝑠), as a function of the field strength for different values of ℓ/𝑅. For finite thermal diffusion, the net force is reduced and so is the speed of the vesicle.
Concluding remarks
Appendix:Brownian dynamics simulations
Conclusions and outlook
Conclusions
Outlook
The rotational operator
The orientational moments
The spatial moments