79
The taxon pulse: a general pattern of lineage radiation and extinction among carabid beetles
TERRY L. ERWIN
Abstract. Evolution of carabid beetles seems not be a random process whereby natural selection has infinite limits. Selection agents are repetitious through time and they have worked on a basic, though diverse, genetic and phenetic groundplan. This process leads to convergence and parallelism throughout the family in structural features and geographic patterns. Taxa arise in equatorial latitudes and are driven through stages along pathways to extinction in three directions: toward the poles, up mountains, and into the forest canopy. Driving forces are many and varied, including development of more potent adversaries, climatic cycles, tectonic events, and microhabitat changes based on floral evolution, but evolutionary results are of a repetitious nature through time as evidenced by disparate groups of carabids following the same paths over and over. This is certainly not a totally new concept, but rather a new formulation or synthesis of many concepts. The realization that changes occur along ordered pathways and are repeated lineage after lineage in the history of major groups leads to the conclusion that evolution has boundaries in which it operates, and that perhaps there are some rules.
Introduction
Patterns are the grist of the natural historian, phylogenist, and biogeographer;
patterns are found in the synthesized data points of the ecologist and ethologist;
patterns allow reasoning hominids to understand their surroundings, their past, and for the more clever among them, their future. Patterns are all about us, in our daily lives, in world events, and in the life forces of the biosphere. As biologists, we are lucky in that we can spend our lives observing these biospheric life forces, and through them catch glimpses of past events which shaped the world in which we live today.
It was with distributional patterns of beetles and cold-blooded vertebrates that Philip Darlington (1943,1948 respectively) set the stage for a modern understand- ing of animal distribution and their past movements over the face of Earth. When Darlington wrote his 1943 paper, the relationships among carabid beetles were not as well understood as they are today. Nevertheless, the patterns Darlington detected, general as they were, gave him the opportunity to discuss the following:
(1) area limitation and its influence on dispersers (or 'founders' as Mayr (1942) called them); (2) why certain beetle features (such as flightlessness) are found
G.E. Ball (ed.). Taxonomy, Phytogeny and Zoogeography of Beetles and Ants.
(E) 1985, Dr W. Junk Publishers, Dordrecht. ISBN 90-6193-511-3.
more commonly on islands and mountains than in lowland continental habitats;
and (3) evolution of species in temperate and tropical climates. Almost contem- poranously with Darlington, Simpson (1949) elaborated points made by Darwin (1859) in a scenario describing the history of a phylogenetic group (mammals and reptiles). Brown (1957), deeply influenced by Darlington, wrote about the mech- anisms of how animal populations underwent centrifugal speciation emphasizing centrist populations as the principal source of evolutionary change and how population density fluctuations aid in spreading characteristics and making and breaking contacts between populations. Wilson (1959, 1961) brought many of Darlington's notions to focus with his description of taxon cycles in ant taxa of Melanesia, and Greenslade (1968, 1969) and Ricklefs and Cox (1972) amplified Wilson's ideas. Numerous other authors, including me, have analysed extant carabid distribution patterns and variously proposed that these resulted from such events as continental drift, dispersal across watergaps or landbridges, vicariance, and so forth. Recently, having been stimulated early in my career by Darlington and after working out carabid phylogenies according to Hennigian principles and observing distribution and natural history patterns of carabid beetles, I (1979b, 1981a) proposed a unified (but briefly described) theory on the evolutionary process determining where these beetles are today geographically and phylogenetically, after a history of more than 250 million years. In developing this theory, I used the building blocks laid by many past and recent workers, particularly Philip Darlington. Thus, it is the purpose of this paper, and appropri- ately so as part of this book honoring and remembering him, to elucidate and exemplify the nature of carabid 'taxon pulses'. Here, I discuss these in relation to geographic and structural (phylogenetic) patterns. In a separate paper, I will examine the relationship of behavioral (feeding, running/wedge-pushing, disper- sion, etc.) and physiological (wet-dry adaptations, hot-cold adaptations, repro- duction rhythms, etc.) traits to the pulse theory and in another article provide a synopsis and historical review (Erwin, in prep.) of the ideas leading to the present synthesis.
Methods
In past studies on carabid beetles, there has been a tendency to work only on southern groups without consideration of their sister relationships with the north or visa versa, or to believe that polar (as opposed to equatorial) groups have been always been cold-adapted and that they descended from cold-adapted lineages.
These constraints have not allowed workers to judge the patterns before them with a broad perspective in time and space. The other problem is that studies are often made on such narrow groups that general patterns are not even detected;
then, from these narrow studies, deductions are generalized too broadly.
Analyses such as the present, are accomplished through extensive studies of
taxa of entire families, or at least, more inclusive taxa and this is best accom- plished by exposure to vast collections and to more senior workers with vast experience. I have been fortunate in that I had the unique opportunity to study under G.E. Ball, P.J. Darlington, and C.H. Lindroth during my formative years as a carabidologist. Subsequently, I became based at the Smithsonian Insti- tution's National Museum of Natural History (more than 1/2 million carabids) and have curated entire collections at the Museum of Comparative Zoology, the American Museum of Natural History, the National Museum of Natural History, and worked extensively on collections at the British Museum (Natural History), Museum National d'Histoire Naturelle in Paris, The California Academy of Sciences, and the Strickland Museum at the University of Alberta. In addition, I have been able to do great amounts of field work, which allowed me to know most groups in life, rather than just as dead carcasses. These opportunities permitted me to gather observations, the sum of which is presented below as patterns. In order to recognize and discuss these patterns, it is imperative to have a sound classification. In making comparative statements, one must use truly comparable subjects, if one is to construct testable hypotheses and arrive at reasonable answers. Although carabid classification is reasonably accurate now, many in- cluded taxa of similar rank perhaps are not equivalent in terms of lineage age.
Hence, global events affecting the taxa under discussion may not be the same, thus the taxa are not directly comparable. For example, past classification have ranked the genus Agra as a tribe comparable with Lebiini. This ranking implies, in a Hennigian system, that they are of the same age and the same global events have affected present distributions of taxa of the two groups. Recent studies show Agra to be a highly derived lebiine genus related to Callidiola (Erwin 1982), and therefore the species, of Agra must be discussed within the lebiines and not compared with the whole set. Other examples are the Eucheilini, now considered a group of the pericalina lebiines (Ball & Shpeley 1983) and the Amblystomini, now considered part of the Harpalini (Noonan 1976). Here I use the traditional names, but write of lineages that I believe to be comparable in terms of age of origin. Appendix A provides a current classification of carabids based on the cladogram in Figure 3 which included analysis of structural features, chemical features, habitat, and behavior of both adults and larvae; further development of this will be found in current studies by Erwin and Stork (in press). I present this modified classification here and it is based on all the latest information at my disposal including Bils (1976), Moore (1979), Burmeister (1976), Evans (1977, 1982), Goulet (1983), Jorge-Silva and Costa (1983), and on reanalyses of numer- ous earlier studies by Bell, Ball, Jeannel, Lindroth, and others (see Ball 1979;
Ball & Erwin 1983). In addition, many new ideas and subsequent rearrangements of groups come from discussions with Ball, Kavanaugh, Stork, and others. I made liberal use of these discussions and I thank these colleagues for numerous hours of their time. However, the final arrangement is mine, and as can readily be seen, I do not necessarily share interpretations of character significance with some of the
above authors. However, I do take full responsibility for my own errors and faulty reasoning.
In the present study, I identified spatial patterns of taxa and analysed the relationships of these patterns among the taxa. Evolutionary trends in structural development also provided patterns and I examined these in terms of geographic and habitat distribution. Finally, I used reciprocal illumination to form the final presentation of patterns by subjecting phylogenetic patterns based on structure to geographic pattern analysis in order to determine, at least, hypothetical con- gruence. With these scrutinized patterns in mind, I attempted to recognize a general pattern of lineage radiation and extinction among carabid beetles and provide an explanation in evolutionary terms. These results are capsulized in a theory of taxon pulses.
Taxon pulse: a definition
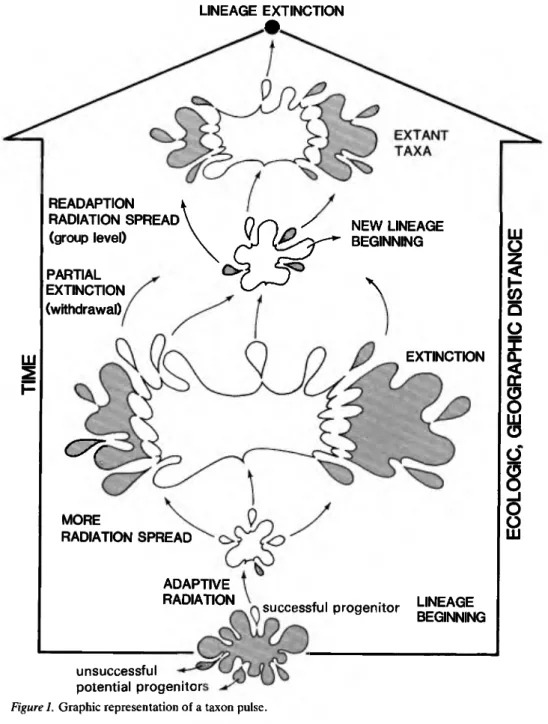
Taxon Pulse is the term I adopted (Erwin 1979b) for the combined adaptive shifts through geographic space experienced by a lineage of organisms from its incep- tion to its cessation. Embodied in the theory of pulses is the concept that generalist groups occupy equatorial wetland centers and these groups undergo special adaptations and geographic movements through time (Fig. 2). The groups are replaced centrally by younger groups, which in turn undergo radiation, movements, and special adaptations. Replacement can be by derived members of the replaced taxa or by unrelated taxa. Most surviving lineages are located peripheral to their centrist ancestors (ecologically, altitudinally, geographically, and structurally). Elements of older lineages that survive centrally or peripherally are those which become specialists, and often their descents become super- specialists; it is common that the latter are located furthest from the common ancestral center of the group ecologically, altitudinally, geographically, and/or structurally. Ultimately, after a lineage has been replaced ecologically at its center by an unrelated group, and its specialists and superspecialists taxa become extinct peripherally, the lineage becomes extinct and the pulse is over (Fig. 1, top).
In other words, a taxon pulse is the complete adaptive and geographic history of a lineage from its monophyletic beginning to its end (Fig. 1), that is, extinction of all its progeny. Although we can observe only the patterns left by pulses of lineages, their history includes the processes and driving forces which brought about the shifts, the pathways along which the processes occurred, and other events of lineage succession occurring through time and space. Of course, such pulses have a beginning and an end in time and space only in terms of our study of them because, in reality, evolution occurs in a continuum in time with numerous epacme (Haeckel 1866) or punctuated equilibria (Eldredge & Gould 1972).
Flowering of taxa is not a new idea (see Erwin 1979b, for a list of other synonyms of this phenomenon), but none of the past explanations were based on strict
recognition of monophyly, repetitive adaptive shifts by successive lineages along the same pathways, and analysis of patterns of extant taxa (for the most part) for the data set upon which hypotheses are erected. In addition, most attempts at explaining taxon cycles dealt with island faunas and did not consider that 'islands' also occur in the middle of continents and are of an ecologic nature (for example,
LINEAGE EXTINCTION
LU
READAPTION RADIATION SPREAD (group level)
PARTIAL EXTINCTION (withdrawal)
NEW LINEAGE BEGINNING
EXTINCTION
C7r
MORE
RADIATION SPREAD
ADAPTIVE RADIATION
unsuccessful potential progenitor
Figure 1. Graphic representation of a taxon pulse.
successful progenitor LINEAGE BEGINNING
o
LUCO Q
g
i Q.
<
(5
rr
O LU
CD O O O _i
o o
LU
see various papers in Prance 1982) and that these ecologic islands have shifted with rifting continents, changing world climates, polar shifts, and geologic time.
In terms of evolving lineages, sequential movement through space, or successio- nal habitats, and concomitant sequential adaptive adjustments in structure, behavior, and physiology, and the genomes which temporarily fix these in taxa, are leaving their mark in extant patterns. It is up to us to discover and recognize those patterns and synthesize them into a meaningful and testable evolutionary scenario with predictive value. For carabid beetles, the nature of taxon pulses is at a hypothesis stage as given below.
Taxon pulse operatives
The so-called Taxon Cycle (Wilson 1961) is a beginning operative of the Taxon Pulse, as are, what I shall call here, 'Darlingtonian Waves' which deal with movement of animals through geographic space. The former, according to Wil- son (1961:187), refers to species with continental (or very large island) 'headquar- ters' and stable populations which repeatedly experience the loss of dispersers to smaller islands, saturating the marginal habitats there. In time, speciation occurs by various mechanisms (Brown 1957; Mayr 1963; Endler 1977; Erwin 1981a;
Erwin & Adis 1982; Bush 1975) with 'marginal' species moving into more cen- trally (on islands) located habitats, for example upland forests, and being re- placed in the marginal habitats, for example sea beaches and lowland scrub, by sequential waves of dispersers from outside sources. The pattern is cyclical, but not the species, of course. Wilson did not extend his ideas beyond the ant fauna of Melanesia and limited his discussion to movements of animals from mainland to islands (that is, land surrounded by water), but he was on the track of the larger picture that I deal with in this paper when he described evolution of 'potent new genera and higher taxa' going from larger source areas to smaller recipient areas rather than the reverse (see his figure 9).
'Darlingtonian Waves' are sequential movements of animals from evolutionary centers to peripheral areas, through time, as described by Darlington (1957) and further developed by Brown (1957). Again, the pattern is cyclical, but not the taxa in the waves. Both these authors were dealing with geographic movements of animals, implicitly assuming that adaptations to new habitats were concomitant with movement.
There are additional observable operatives in carabid Taxon Pulses, including acquisition of dominance (see below), structural and physiological adaptations (Thiele 1977), behavioral adaptations (Den Boer 1977; Den Boer et al. 1979), altitudinal and latitudinal shifts (Kavanaugh, Ph.D thesis; Noonan 1981a, 1981b, 1982), habitat sequencing (Erwin 1981a, 1981b, and Adis 1982; Allen & Ball 1980), vicariance (Erwin 1970, 1975; Erwin & Kavanaugh 1981; Erwin & Stork 1984 (in press); Noonan 1973; Ball 1978; Spence 1983), and dispersal (Darlington 1971;
Erwin 1979a; Ball 1970; Erwin & Kavanaugh 1981; Freitag 1969; Whitehead 1972).
Generalists, specialists, superspecialists
The gauze through which extant patterns must be viewed is the notion that sequential evolution occurs from a generalist role toward a superspecialist role in the ecosystem in which the lineages terminates. The words, 'generalist' and 'specialist', have specific meanings in the fields of ecology and phylogeny. Thus, it is important to define them in terms of this paper. These two words should always be used in relation to parts of the same organism, and should not be used in context of whole organisms except in an abstract way when relating overall generalness between groups! The latter, though often seen in the literature, is not very useful and often obscures the author's intended focus. For example, carabid beetles have a rather uniform basic structural ground plan throughout the family's 86 tribes and 40,000+ described species. This groundplan is that of a generalist beetle (using the abstract meaning above), relative to the groundplan of bruchid beetles or seed weevils; that is, there is less deviation from the megalopteroid ancestral structural assembly in carabids than in extant seed weevils.
Variations on the carabid generalist theme occur however, and these may be extensive and highly specialized, such as the entire feeding mechanism of cicin- delines and hiletines, or minor, such as the pattern of marginal elytral setae in tachyinas. Further, a generalist groundplan such as in hiletine body structure can be coupled with the most highly specialized feeding mechanism found in the entire family (Erwin & Stork in press). Thus, a species may be a 'generalist' runner/weak wedge-pusher and a 'specialist' feeder at the same time (for exam- ple, Loricera). Similarly, a species may be a generalist structurally, but be a specialist physiologically and live under ice margins of montane glaciers (for example, Nebria). A parallel and related problem occurs in the jargon of cladists, that is those who use the Hennigian system of systematics (classification). Prop- erly, character states or even character systems are described as 'plesiotypic' or 'apotypic', that is, less derived or more derived relative to each other. Again, these two terms can only be used in relative context. Often however, many authors have identified entire taxa as plesiotypic or apotypic. These authors mean of course, that the species (or higher taxon) members have more primitive (less derived) character states than derived ones (within those studied and analysed) or visa versa, than in other in-group species or higher taxa. The problem with such loose use of these words is that meanings are lost and confusion reigns. In the future, it would be more appropriate to use modifiers with the words generalist and specialist, for example, feeding-specialist, running-specialist, and in the case of plesiotypic and apotypic, restrict these to character states.
Specialisation of a character system, whether it proceeds independent of, or concurrent with, other systems, occurs in space and time in organismic lineages.
Occasionally, such as in the development of the single-toed horse, genus Equus, a nearly complete lineage with failed side branches is preserved in the fossil record.
In carabid beetles, many such lineages are preserved (as a developmental pat-
tern) in persistence of side branches in the extant fauna. Thus, various stages in the development of specialisation can be inferred by examination of enough taxa, connection of homologous features, and determination of correct polarity of development. Placement of such development in time and space is difficult however, because formative geologic and climatic events which influenced the patterns have drastically altered and left little traces at fine enough resolution to be helpful.
Patterns among carabid beetles
Numerous patterns are exhibited by carabid beetles in: geographic distribution, habitat preference and other behavioral traits, character-state distribution, and climatic adaptations and other physiological traits. These patterns, together with the phylogentic relationships among the lineages which exhibit them, led to the development of Figure 2. In this section, I describe in detail some of these patterns to exemplify their repetitious nature among carabid taxa and show their distribution along the pathways to extinction seen in Figure 2. Figure 3 illustrates a hypothetical phylogeny of early caraboids and ages of lineages; in the text below, reference is made to these lineages and ages. Numbers on Figure 3 refer to characters listed in Table 1. First, I discuss spatial patterns of taxa in terms of hypothetical phylogenetic progression, that is assumed age of taxa in relation to each other. Secondly, I discuss structural patterns among taxa in relation to geographic and topographic distribution, ordering the discussion in terms, again, of hypothesized phylogenetic progression.
Spatial patterns
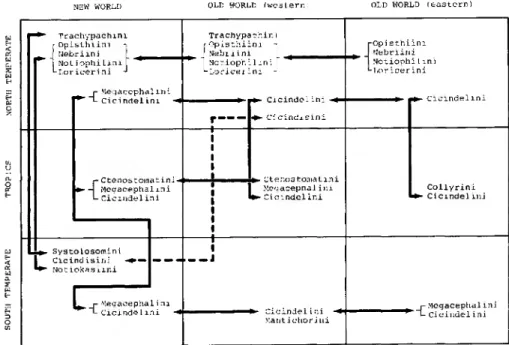
The north temperate pattern. - At the taxon level of tribe (see below for Tra- chypachidae), very few groups have a northern temperate distribution without having a sister taxon in the southern hemisphere. Only the old carabid lineage Loricerini and the monobasic Gehringiini are found solely in the north temperate zone with no known southern sisters (Figs. 4, 8). These groups are Holarctic (Loricerini) or Nearctic (Gehringiini) in distribution. The Gehringiini are still an enigma as far as relationships go; they may be ancient (Lindroth 1961; Evans 1984) or related to the Scrobifera (Erwin 1979b) and thus middle-aged. Other lineages, previously thought to have solely north temperate distribution do not and are treated below. All other groups, either at the tribal, subtribal, or generic level have tropical and/or south temperate relatives and are also discussed below.
The south temperate pattern. - Only one higher taxon, thought to be in the middle- aged lineages and thus not so old as those discussed above, is found solely in the south temperate zone, namely, Zolini(Fig. 8). The relationship of this group with
X o
NUMBER OF SPECIES ALTITUDE
G O
•o
_3
"3 >
3
D Xi
W H
ca < C
W H
o z, <D J u
o
T3
s so
Table 1. Cladistic classification of character states for selected higher taxa of Caraboidea
Character Plesiotypic state Apotypic state
Head
1. Antennal flagellum Glabrous Pubescent
2. Antennal base Exposed Partially covered (a)1
Fully covered (b)2
3. Antennal scape Short, compact Elongate, narrow
4. Subgena Normal Grooved
5. Mandible Crushing/chewing Piercing
6. Scrobe Unisetiferous Glabrous
7. Mouth appendages Normal Mycelivorous
8. Attitude Prognathus Partially hypognathus
Thorax
9. Pro/meso-connection Broad Pedunculate
10. Mesepimeron Broad Narrow
11. Mesocoxa Separate Conjunct
12. Metepimeron Absent Distinct
Elytron
13. Margin Smooth Coanda
14. Texture Smooth Beaded
15. Shape Normal Depressed
Legs
16. Anterior tibiae Normal Fossorial
17. Unguitractor plate Undeveloped apically Tab-form (a)1
Seta-form (b)2
Absent (c)3
18. Dorsal guard setae 2 1
19. Claw base setae Absent Present
Vestiture i
20. Tactile setae Normal distribution
and size Increased number, size
Male genitalia
21. Median lobe, base Lobes symmetrical Lobes asymmetrical (a)1
Lobes fused, open (b)1' Lobes fused, bulb-form (c)2'
22. Median lobe, dorsum Sclerotized Membraneous
23. Parameres Stylate, setiferous Glabrous (a)1
Non-styliform, brushy (b)1' Non-styliform, setiferous (c)2' Conchiferous, glabrous (d)3' Left glabrous, styliform (e)1"
Abdomen
24. 7th segment Smooth Microserrate
25. Defence mechanism Bilateral, open tubes Bilateral, turret tubes (a)1
Medial, exposed (b)1' 26. Defence chemistry Saturated acids, etc. Quinones
Note: Characters with more than one state are coded as follows: lower case letters designate different states; numbers designate transformation series, 1, 2, 3, n, or branched pattern if primed. This classification is, for this paper, necessarily terse, however it is meant to demonstrate what happens to the system if some traditional characters are set aside or considered to be convergent. It is offered as an alternative hypothesis for the purpose of testing.
NEW WORLD
Trachypachin r Opisthtim.
J Nebriinj I NoL i ophi Lini
•- Lor iceri ni
• Heqaccphalini - Cici ndeL ini
OLD WORLD (western)
Trachypachin i Op Lsthiini Nebr i ini Nonioph il m i.
•Loricerini -1
*- Cicindolini -^
*- Cicindxsini
OLD WORLD (eastern)
•Op i sthiim Webriini Notiophi I in.i
•Lor icer ini
• Cicindelini
•{
Ctenostomat inj Mcgacephalxni Cicindelini- Ctenostomatmi MeqacepnaJ ini - Cicindclini
Collyrini
• Cicindelini
Systolosomini Clcindisini NotiokasLini
Z
_r Megacephali ni"L Cicinde1ini Cicindeli ni Manti chorini
i- Moqacephalini
~L Cicindelini
Figure 4. Graphic depiction of spatial distribution of primitive carabid tribes centered around the nebrioids.
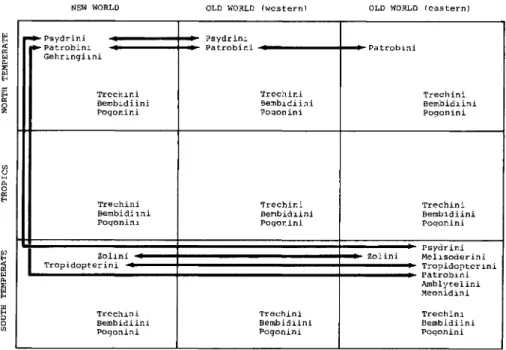
other carabids is not clear now. If they are part of the Trechitae, they are undoubtedly a very early pulse of that group that successfully adapted to harsh south temperate conditions, just as later trechines adapted to mountain tops and arctic conditions. I include them here rather than with the trechines because the relationship is not clear.
The amphitropical pattern. - Numerous groups exhibit a pattern of distribution which bridges the tropics, either by total absence of equatorial taxa or by a paucity of taxa and dominance of a derived sister group in the tropics. These two kinds of patterns seem to be related to age of the group; one pattern, that of absence from equatorial areas, is confined to ancient lineages, while the second pattern, that of replacement by derived sister taxa, is found in middle-aged or younger lineages. Often, in the case of the latter, sister taxa are found mostly or only on tropical mountains or sea beaches (for example, Bembidion, trechines, agonines).
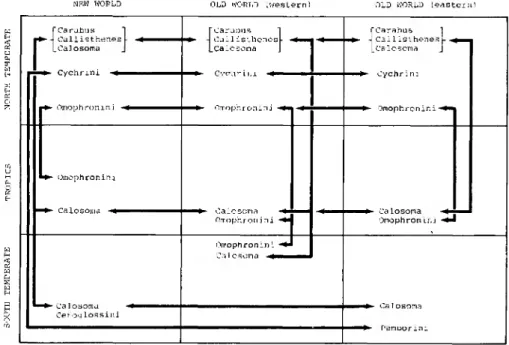
Amphitropical pattern I. - Old amphitropical relationships are many (Figs. 4,5,7, 9); most are not striking because of low numbers of taxa in the south, for example, nebrioids/Notiokasiini, Cychrini/Pamborini, CarabuslCeroglossus, Elaphritae/
Migadopitae, Metriini/Mystropomini, taxa within Promecognathini and Cicin- disini, and the Trachypachidae, TrachypachuslSystolosomus. These groups, for
the most part, are more diverse in the north and less diverse in the south at the species level; in addition, none have tropical representatives. An exception to the north/south diversity pattern is the southern supertribe Migadopitae, which is more diverse than its northern sister. In several instances among the taxa men- tioned, there are only one or two species in the south.
Amphitropical pattern 11. - Amphitropical patterns exhibited by middle-age or younger groups are more impressive in terms of numbers of taxa, especially since they have a large tropical sister component. For example, the lineage Bembidion, with more than 1000 species, provides an example of a nearly perfect amphitropi- cal pattern (Fig. 8); the only species occurring adjacent to the equator are those on mountains (temperate climate), in the arboreal ecosystem, or along sea coasts.
The pattern of the more derived sister lineage of Bembidion, Tachyina (Fig. 8), is a mirror image. This group is concentrated in the tropics and gradually disappears in north and south latitudes and at higher elevations. This general pattern, found within tribes and not between them, occurs also in Trechini, Pterostichini, Scaritini, Carabini, and among the tribes of Cicindelitae.
The amphiantarctic pattern. - Many groups, both older and younger, have a transantarctic distribution (Figs. 4-8). Younger, mere highly derived groups judging from their structural features and classified usually below the tribal level,
NEW WORLD OLD IVORT.D (western) OLD WORLD (eastern)
r
Ca rabus Callisthenes Calosoma CychriniOmophroninj -^
Carabus Call is Lhcncs Calosoma
Omophronini
Carabus Cal1 is thenes Calosoma
^- Omophronini-
Omophronini
Calosoma Omophron i ni
•*- CaLosona Omophron in
•Ca1osoma Ceroqlossini
Omophronin i Ca1osoma -^
•*- Calosoma
"^- PamD© IT in:.
Figure 5. Graphic depiction of spatial distribution of primitive carabid tribes centered around the caraboids.
NEW WORLD OLD WORLD (western)
•*- Broscini -*•
Apotomini
OLD WORLD (eastern)
^*~ Broscini
Apotomini-^ Apotomini -
Melaeniini •
Apotomini - Melaeniini
Broscini -* N- Broscini
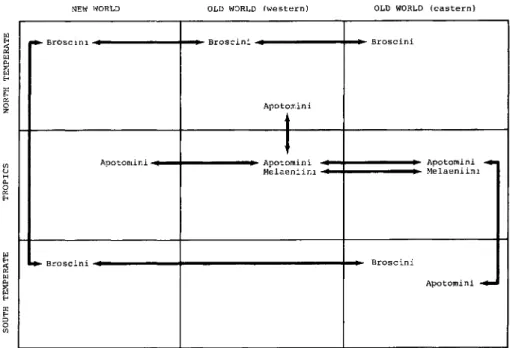
Figure 6. Graphic depiction of spatial distribution of primitive carabid tribes centered around the melaenoids.
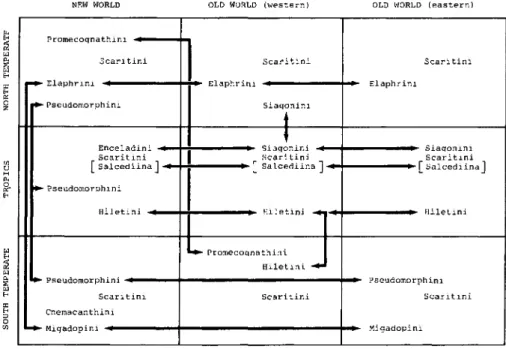
NEW WORLD OLD WORLD (western) OLD WORLD (eastern)
Promecoqnathini -*^
Scari tini
• Elaphrini -^^- -Pseudomorphini
Scaritini
*- Elaphrini -^N Siaqonini
Scari tini
•*- Elaphrini
Enceladini -*•
Scaritini [ Salcediina1 -^
-Pseudomorphini Hiletini «
Siaqonini -*•
Scari tini -*- f Salcediina 1-^
i^^- Siaqonini Scaritini
*- fSalcediina1
-*~ Hiletinj -+1-* •*- Hiletini
• Promecoqnathini Hiletini - Pseudomorphini -^^^•
Scaritini Cnemacanthini - MiqadopLni -^^^^^^•
Scaritini
Pseudomorphini Scaritini
Miqadopini
Figure 7. Graphic depiction of spatial distribution of primitive carabid tribes centered around the scaritines.
NEW WORLD OLD WORLD (western) OLD WORLD (eastern)
-Psydrini - Patrobim
Gehringiini
Trechini Bembidiini Poqonini
Psydrini Patrobini
Trechini Bembidiini Poqonini
Trechini Bembidiini Pogonini
Trechini Bembidiini Poqonini
Trechini Bembidiin Pogonini
Trechini Bembidiini Poqonini
Zolini -^
Tropidopterini »• '
Trechini Bembidiini Pogonini
Trechini Bembidiini Pogonini
Psydrini Melisoderini Tropidopterini Patrobini Amblytelini Meonidini Trechini Bembidiini Poqonini
Figure 8. Graphic depiction of spatial distribution of primitive carabid tribes centered around the Psydrinae.
for example, Loxandrus, Pericompsus, and notiobioids, exhibit a typical subtrac- tion pattern from the Neotropics, through southern South America, across western Antarctica to southeastern Australia, and from the Neotropics north into the southern United States. The amphiantarctic part of the pattern is the mirror image of the subtraction pattern across Middle America into North America. A variation of this pattern, that is absence from one or more portions of the pattern, in some taxa probably resulted from extinction in the South American tropics, for example, Pasimachus-Carenumy etai, Tachyss. str., and Psydritae. The Amphi- antarctic Pattern is easily explained when it is realized that western Antarctica was ice free until 9 million years ago (Kerr 1981, 1982). Thus species groups distributed across this area were probably subjected to vicariance by ice develop- ment rather recently in terms of their evolutionary history. Until 16 million years ago, Antarctica was likely extensively ice free (Kerr, ibid), thus distributions of other carabid groups undoubtedly extended across it on both the eastern and western flanks, including Migadopini, Pseudomorphini, and some Scaritini, Zolini, Broscini, the psydrite and panagaeite complexes, and Pterostichini.
Transatlantic pattern. - A large number of taxa are clearly transatlantic in terms of their sister group relationships. For the most part, these are at the generic or subtribal level. The Transatlantic Pattern (south) includes such taxa as found within Ozaenini (part), Brachinus, Pheropsophina, Callidina, parts of Lebiini,
Galeritini, Helluomorphina. Other groups are transatlantic (north), for example, Tachyta, Anisodactyloids, some pterostichines.
The pantropical pattern. - Numerous genera and subtribes of higher carabids are pantropical in distribution, extending beyond the limits of the transatlantic distribution described above, particularly among the Agonina, Selenophorina, Calleidina, Callistini, Odacanthini, Ctenodactylini, and others. Some parts of these groups also form a subtraction pattern into northern (for example, Chlaenlus, Calleida) or southern (for example, ctenodactylines) latitudes. Two old groups, the highly specialized Hiletini and Siagonitae, also share this type of distribution (Erwin & Stork in press).
The transberingian pattern. - Numerous Holarctic genera or species groups owe their distribution to the presence of a Bering Land Bridge which appeared at various times during the Tertiary and Pleistocene (Lindroth 1979; Kavanaugh 1979), for example, species groups of Nebria, Pterostichus, My as, Bembidion, etc. Virtually all these holarctic taxa are 'young' in terms of evolutionary age of extant species, although at higher rank they are very old lineages.
The cosmopolitan pattern. - The few taxa truly cosmopolitan in distribution are species carried by human transport, for example, Perigona nigriceps, and are capable of breeding in decaying vegetation of nearly any kind, a microhabitat created by the transporters.
The Malayan Archipelago filter pattern. - Darlington's (1979) excellent analysis of carabid taxa distributed across the southeast Asian islands from the mainland to Australia shows that it is only recent species or species group stocks that have spread in this region. Most of the dispersal traffic has been from the Oriental mainland south to New Guinea and Australia. A similar series of events probably occurred when the Indian plate reached Laurasia; whatever was on the Indian subcontinent when it collided with the northern shores of the Tethys seaway was swamped by Oriental species (Erwin 1984). Old stocks found in south India and Africa are few (for example, the two crepidogastrines of the genus Tyronia) and probably were more extensively distributed in the past (Erwin 1971). This filter has acted on certain recent groups for the most part because there are few ancient or old groups in the Oriental region (south). Of the few older groups available, such as the Pheropsophina and Siagonini, only a few species have been able to penetrate southward. From the south, no older groups have been able to pene- trate northward. Those recent groups which have dispersed are ones which live in the lowlands and easily cross water barriers (for example, harpalines, scaritines, tachyines).
The Central American filter pattern. - Like the Malaysian Archipelago filter,
volcanic activity and sea level risings in Middle America through time has prevented the majority of groups from spreading north or south. However, several groups that have managed to do so occur in typical subtraction patterns in one direction or another, for example Brachinus, Agra, Loxandrus, Elaphropus, etc. Unlike the Malaysian filter, old and middle age groups (for example, Pasi- machus, Loricera, Notiophilus, Calosoma, and paussines and brachinines) and those of mountains have been able to use the more continuously connected Middle American landmass together with the younger lineages (see Ball 1970).
The Pacific waif ting pattern. - The islands of the Pacific have numerous carabid species, but of only a few tribes. Two patterns exist, (1) old lineages, with flightless species occupying the uplands of most islands with appropriate to- pographic relief, viz., psydrites, agonines, and Bembidion; (2) recent species which have been transported by human movement of goods and plants or dispersed by natural means such as rafting or through the air (Darlington 1970).
Larger islands of the southwest Pacific, such as New Zealand and New Cale- donia, have special histories and the faunas are related to Australia, more continental in nature, and are, in general, old assembleges of early Australian stocks.
The Atlantic waifting pattern. - The islands of the middle Atlantic have carabid faunas that, in part, undoubtedly date to rifting of the major continents or shortly thereafter (for example, Aplothorax). For the most part however, taxa are most closely related to the fauna of the closest continent, either Europe or Africa, (for example, Calathus, Loricera, Bembidion, Carabus) and these stocks arrived over water by rafting or through the air; those which arrived early have formed endemic species groups, especially in the Canaries. Some larger island groups, such as the Canaries and Azores, have also received some species probably introduced by human transport (Lindroth 1960a).
The Indian Ocean waifting pattern. - The pattern in the Indian Ocean is the same as in the Pacific except, since there are no suitable upland habitats on remote islands, there are no old flightless groups represented. However, there are endemic groups classified at the generic level on Mauritius, and endemic species on the Mascarenes (for example, Halocoryza maindroni), Sri Lanka (Erwin 1984), and probably other islands. Many species on islands adjacent to Africa are of African or Madagascan origin and most still exist on the mainland (for example, see Basilewsky 1970); those adjacent to India are of Indian origin and, except for Sri Lanka, there is little or no endemism. Although Sri Lanka is an island on the continental shelf and very close to the mainland, it has 7.9% of its carabid fauna endemic (Erwin 1984). Much of this is due to the availability of a high central massif.
Structural patterns
Flightlessness. - Flightlessness in carabid beetles is well documented and Dar- lington devoted much time and words to the subject in several papers (for example, 1943, 1979). However, a synopsis is not available for the family as a whole. In general, only certain lineages have taken the path to flightlessness and thus forever losing the power to disperse readily. It is entirely possible that some groups with flight wings also have lost powers of flight by a number of internal mechanisms, such as atrophy of muscles or wing muscles innervation, without change in sclerite shapes, but these seem to me to be physiological responses to special situations and possibly may be reversible (this must be investigated by physiologists and geneticists). Structural changes in sclerites concomitant with wing loss however, certainly must be permanent and not reversible. The evolu- tionary reasons for wing loss, muscle reduction or its equivalant, and the resulting loss of flight are unknown. Since the time of Darwin, students have speculated on the subject, arriving at such conclusions as (1) avoid being blown away in a storm from mountain tops and islands; (2) saving of developmental energies for with- standing rigors of harsh environments; (3) shifting of developmental energies into ambulatory adaptations for certain kinds of environments; (4) no longer needing flight in certain habitats such as caves. The answer to 'why flightlessness?' could be all of these and study is needed across the family in all the habitats in which this phenomenon occurs.
Lineages with stuctural changes in sclerites resulting in flightlessness may be categorized in two ways: (1) by group age or: (2) by group adaptation with respect to altitude or latitude. Old groups, at either the center or periphery of their ranges tend to be flightless; younger groups tend to be flightless mainly or only on montain tops or in the boreal and arctic zones, thus at the limits of their overall ranges. Ancient, old, middle-age, and young lineages (Fig. 3) are, for purposes of discussion, convenient categories of age while altitudinal and latitudinal adapta- tion trends are convenient categories of distribution. Some lineages are discussed in more than one of these categories.
Numerous ancient and old lineages are represented today by flightless relicts in various parts of the world. These are mostly in the far south and some are amphipolar (for example, Ceroglossini, Pamborini, Mystropomini, Migapodini, Cnemacanthini) or in predominantly nonglaciated regions of the north (for example, Carabus, Cychrini, Metriini, Promecognathini). Very few ancient and old lineages are represented by flightless taxa occurring in the lowland tropics (some Cicindelites, Enceladini, some Hiletini and Scaritini). Most ancient and old north and south temperate groups are represented by all flightless species in entire genera or even tribes. None of the ancient lineages are strictly montane, although species may occur at higher elevations; nor are any of them arboreal except some members of a group of cicindelites which occur on tree trunks and mimic ants (Ctenostoma, Pogonostoma, and Tricondyla). Taxa which live under
bark of trees, such as Enceladus, are not considered arboreal. The pattern is almost exclusively latitudinal, in the north and south temperate zones.
Some totally flightless lineages presently classified in the middle of Carabidae, and presumably of Cretaceous origin show remarkable geographic patterns. For the most part, these Middle-age lineages are amphipolar in distribution with regard to Antarctica (Peliciini/Agonicini/Disphaericini, Zolini, some Psydrinae).
Because these patterns are very similar to some of those discussed under ancient and old lineages (above), it is possible that these groups represent very old lines within their respective supertribes and probably gained their present distribu- tional pattern at the end of the period of ancient pattern development, that is, early Cretaceous. The majority of middle-aged lineages have taxa with macrop- teran adults or those in which wing development is dimorphic or polymorphic. In almost all species, brachyptery occurs in higher latitudes or altitudes, for example many scaritines occurring in cloud forests above 2000 m. I know of one Ar- distomis species at sea level in a tropical forest habitat, Barro Colorado Island, Panama, which is wing-dimorphic; others have been reported, but are very rare.
No arboreal species derived from middle-age lineages (for example, Pseudo- morphini) are brachypterous and 'i know of none that are wing-dimorphic. Thus, again the pattern is almost exclusively latitudinal, in the north and south temper- ate zones, or altitudinal in the tropics as well as in temperate zones.
Recent lineages, classified as Harpalinae here, were derived from basal stocks of the middle-age lineages, probably as a radiation of the psydrites. This early radiation resulted in worldwide complex patterns that are difficult to interpret, for example Pterostichitae. Considering this latter group in the strict sense as an example, the pattern consists of a markedly diverse agonine complex which is centered on the equator, and another markedly diverse pterostichine complex which is centered in north and south temperate zones. This pattern is exactly the same as in Bembidiini, and the two are probably contemporary. There is little doubt that the pterostichine complex is a grade and that many groups have evolved from the tropically centered agonines. In so doing, many 'pterostichine' taxa have become flightless relicts in high latitudes and altitudes, while few agonines (classical sense) have become latitudinal relicts. In tropical mountains however, agonines dominate and most of these are wingless or wing-dimorphic.
Many other groups of recently derived carabids are found just about anywhere in habitats that are somewhat stable over long periods (even though annual stability may be low, for example, river banks) and consist of those lineages in which brachyptery appears in only some of a population's individuals. This has been termed 'wing dimorphism' by numerous carabid workers. When peripheral populations are isolated and undergo speciation, it is possible that brachyptery becomes fixed as dominant and eventually all adults are flightless. This is proba- bly one mechanism whereby permanent flightlessness occurs in species. There is a greater tendency for certain groups to become brachypterous wherever they occur in the world; these same groups are found also as altitudinal and latitudinal
Fossorial adaptations. - Only a few groups of carabid beetles have managed to adapt to, and radiate in, subsurface conditions in self-made tunnels. The scari- tines, cnemacanthines, and a few desert and steppe harpalines and zabrines have acquired fossorial front legs and/or fossorial middle and hind tibiae which do not excavate like the front tibiae, but do move loose soils in relatively great quan- tities. Nearly all lineages whose members use their front legs for excavation live in finely textured clays or lateritic soils, or in sand, and most of them are tropical.
Groups whose members burrow, other than scaritines, all live in sandy soils.
Omophronines use their heads as earth movers and it is likely other carabids do so as well; while numerous groups, on the other hand, use soil cracks and crevices, worm and rodent holes, etc. to go underground, but I do not regard any of these as fossorial. Evolution of fossorial forms seems logical for ground beetles, but so far there has been no explanation for this adaptation in regard to its driving forces. The entire Scaritini are equiped with fossorial front tibiae and are the only carabids so endowed. Rhysodines have legs adapted to burrowing in wood and in part resemble those of scaritines (Bell & Bell 1962), but I do not regard these as fossorial.
Among ancient lineages, no group has developed fossorial habits of any kind.
Among old lineages, cnemacanthines are loose soil movers but not truly fossorial and these are found only in the south temperate zone. Only a few species of few genera of scaritines have radiated into the North Temperate zone; Dyschirius, for example, it is predominently North Temperate, but many species also occur in the tropics. Among younger lineages, most soil-mover forms occur in grasslands or steppe habitats where soil is sandy and these for the most part occur in the north temperate zone. Fossorial forms are not common at higher altitudes anywhere, but do reach middle altitudes especially in the tropics. Thus, the over all pattern of fossorial adaptation is similar to that of arboreal adaptation, common in the tropical belt and decreasing latitudinally (Fig. 2) and altitudinally.
Troglobitic adaptations. - Very few lineages of hypopigean or troglobitic super- specialists have evolved. These groups are characterized by reduction or loss of melanic pigments, reduced or absence of eyes, elongate marginal setae, elongate appendages, and predatory habits. Among these, most are groups of the Psy- drinae and Harpalinae: trechines, agonines, harpalines, masoreines, and lebiines in caves; and tachyines, anillines, trechines, and agonines in hypopigean habitats.
No ancient group occurs in these habitats, and the only old lineages whose members have evolved troglobitic habits are certain Clivinini; in the middle-aged groups, only zolines, Idacarabus. Some other lineages have adapted to facultative cave living, such as tachyines, pterostichines, and lebiines. These groups are mostly cosmopolitan in distribution; radiation into the cave or hypopigean en- vironment by these groups has happen throughout the non-glaciated parts of the world, especially by the trechines. Among the groups mentioned, only Ida- carabus is restricted to the far south temperate zone. Caves at middle altitude are
also inhabited, but no high monticolous cave fauna is known. The general pattern is one of a few special groups with a propensity toward cave life taking up that life anywhere a cave exists for a long enough period to allow evolution to take place in it. Hypopigean groups are, for the most part, the same general groups which radiate in caves and these occur anywhere in non-glaciated parts of the world.
A synthesis of the patterns
In the beginning
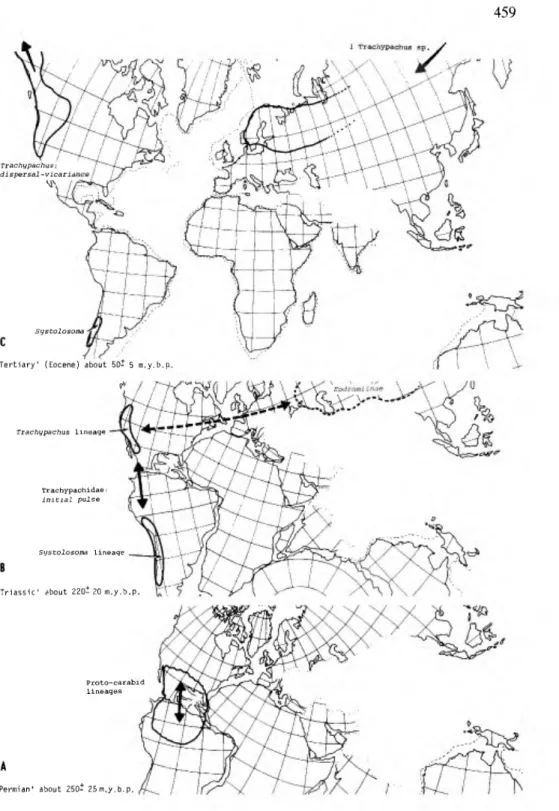
The family Trachypachidae, until recently considered part of the Carabidae, provides the oldest spatial pattern in the 'geadephaga'. Old, good fossil material is available from Madygen series of Central Asia (Middle or Upper Triassic) (Ponomarenko 1969, 1971, 1977). This area was, and has been, in the North Temperate zone since the Permian (Fig. 9), when modern Coleoptera had their beginnings (Lawrence & Newton 1982). These fossils indicate that evolution of terrestrial 'ground beetles' had, already, a long history. Associated water beetles, for example Coptoclavidae, in these early deposits, indicate that some radiation had also already taken place in aquatic habitats. Trachypachids are no longer in the area where the fossils were found, but do occur further to the northwest in Europe, and in the Pacific Northwest of America and Canada, ranging as far south as southern California. In Chile and Argentina, a land area that has always been in the South Temperate zone, another trachypachid genus occurs (Sys- tolosoma), and is the sister group of the present North Temperate genus (Tra- chypachus); thus the spatial pattern of the earliest known terrestrial caraboid group is today partially Holarctic and amphitropical in the New World. The current pattern suggests, giving time for evolution of the group, that cosmopolita- nism of trachypachids was attained by late Permian or early Triassic. The evi- dence for this is fossil material occurring outside the present range and the amphitropical presence of sister groups.
Trachypachids share numerous character states with 'hydradephaga1 and sev- eral more with geadephaga (Evans 1984; Roughley 1981; Hammond 1979). The larvae (Lindroth 1960b) are typically caraboid. Although their position is not completely resolved, the weight of evidence points to the scenario illustrated in Figure 3. These ancient relicts probably represent an off-shoot of the structural groundplan of the earliest caraboids, both in the adult and larval stages. Although today these beetles live in dry dusty habitats in conifer forests, the earliest of their kind radiated in habitats unlike any found today (very early conifers, cycads, tree ferns). These habitats persisted on a megacontinent with climates remote in nature from those today. If this is an example of how the earliest terrestrial caraboid began and we see today how it has ended up, it would be instructive to compare geographic patterns of progressively derived caraboid lineages in a
Trachypachus:
dispersal-vicariance
Systolosoma
c
'Tertiary' (Eocene) about 50- 5 m.y.b.p
«1
Trachypachus Lineage
Trachypachidae initial pulse
Systolosoma lineaqe
'Triassic' about 220- 20 m.y.b.p
Proto-carabid Lineages
'Permian' about 250- 25m..y.b.p
Figure 9. Hypothetical time sequence distribution maps of Trachypachidae from Permian based on extant forms. Fossils found in middle Russia indicate Triassic species were Holarctic. (4a) events of the Permian; (4b) events of the Triassic; (4c) events of the Tertiary.
search for a concordant pattern. However, since no good fossil records exist for these other caraboids, the analysis must be made from knowledge of extant fauna distribution patterns.
The road to dominance
According to Darlington (1957), dominance 'reflects underlying qualities or characteristics that make for initial success and lead to evolution of numerous, varied, successful types' ... 'often (eventually) widely and continuously dis- tributed in unfavorable as well as favorable places'. Even though Darlington was discussing vertebrates, he could just as well have been writing about carabid beetles. They are dominant insects now in a variety of habitats, and have been for considerable time. Dominance has been attained concurrent with the attainment of cosmopolitanism among higher ranks within the family.
Extant carabids (s. str.) presently thought to represent the most ancient lin- eages are the nebrioids (Kavanaugh, per comm.) of the Nebriiformes (Fig. 3 and Appendix A) and these are spatially distributed in two patterns; (1) strictly North OR South Temperate; (2) Amphitropical in the New World with sister tribes in the North and South Temperate zones. Figure 4, in part, graphically depicts these distribution patterns. For nearly all the ancient taxa, a southern sister is found in South America. Although fossil evidence is lacking, it is probable that these lineages lived contemporaneously with early equatorial trachypachids and like them began their existence in wet biotopes of the Glossopteris forests and marshes of that period. They are now members of more or less gymnosperm plant communities, although some recent species may be secondarily adapted to cool- temperate angiosperm plant communities. In many of these habitats, carabid beetles of this group occur in great numbers and are the dominant predators (along with spiders) of cold streamsides and the forest floor.
Another group of carabid lineages of the Nebriiformes, classified and thought to be centered around Carabus, may not be as old as the nebrioid lineages, but are probably derived from a common ancestor, and have four patterns of distribu- tion; (1) Amphitropical; (2) North Temperate/Tropics sister pairs; (3) Pantropi- cal; (4) Cosmopolitan. Figure 5 graphically depicts these distribution patterns.
These groups are broadly adapted to a variety of habitats, including gymnosperm forests, and among them are found the only 'ancient' carabid lineages adapted to modern tropical habitats {Calosoma s. lat., tiger beetles, omophronines). How- ever, only the tiger beetles, for example, Odontochila, are dominant in lowland tropical forests; the remaining lineages are, in part, marginal tropical groups, living in habitats which could be described as subtropical or temperate extensions into the tropical zone, for example, mountain systems (Loricera, Notiophilus), agricultural grasslands (Calosoma).
The numerous lineages classified by Jeannel (1949) as Scrobifera + Stylifera,
plus Siagonitae (which he inexplicably placed with the tiger beetles as Simplicia) and Isochaeta, have been studied by various workers since Jeannel. They are reclassified here on the basis of newly discovered character states (see Appendix A, B, and Fig. 3) and a reanalysis of some previously used states. They are considered as old and middle-aged lineages (Fig. 3) with the names Loximeri- formes and Melaeniformes, respectively. The older lineages among these, proba- bly also contemporary with the later development of Nebriiformes and tra- chypachids, have simple patterns like the latter groups, while the younger lineages have a complex overall distribution pattern as depicted in Figure 6.
Among these, certain subpatterns are well developed and are classed in three general types: (1) North Temperate; (2) Pantropical; and (3) Amphitropical (New World)- Amphiantarctic. In addition to the Loximeriformes and Melaeni- formes, many early Psydriformes, which bridge the gap between the middle-aged and younger carabid lineages, express these subpatterns in their distributions.
Within these three major groupings, is found the first spatial pattern connecting the Oriental and Australian regions (Darlington 1979). The taxa involved, like those ancient lineages discussed above, extend into new zones by virtue of a few widely dispersed species entering habitats or land masses only recently available.
Because of the relationships among taxa and the fact that only recent ubiquitous species (that is, species of the pantropical Brachinini, Apotomini, and Morionini) are involved, the pattern appears to be one of dispersal.
Like the ancient groups discussed earlier, these major old and middle-aged groups (Loximeriformes, Melaeniformes, and Psydrinae) are dominant in the habitats in which they occur, particuliarly waterside and moorland habitats, but also subcortical habitats, gymnosperm forests, ant nests, caves, and mountain tops.
The rest of the Psydriformes, that is Harpalinae (Fig. 3), are distributed mostly pantropically, or as large tropical genera with a few species extended into the North or South Temperate in one hemisphere or the other, or thirdly, confined to one or the other hemispheres in tropical climes. For the most part, these groups are adapted to subdesert, steppe, or broadleaf forest habitats and often are dominant organisms in these biotopes, either as predators, scavengers, or her- bivorous seed-eaters. Most of these taxa, at the tribal level, are so widespread that no patterns are discernable (Fig. 8), except the Pterostichini (see below).
Studies must be made at the subtribal or generic level to detect whatever subpat- terns might exist, but unfortunately, on a world scale, this has rarely been done with currently available methods and knowledge of geological events (but see recent papers by Ball, Bell and Bell, Darlington, Jeannel, Erwin, Noonan, Spence, and Whitehead). The few studies available indicate that these subpat- terns are the result of continental rifting in the early Tertiary and subsequent dispersal of vicariants. The pattern of distribution of the Pterostichini is nearly the same as found in Bembidiini - that is, a tremendous radiation of north and south temperate zone species and accumulated genera with a large sister component
still in the tropics. The tropical forms, although more recent in time in terms of the extant fauna, maintain the primitive and diverse groundplan of the tribe.
Very minor patterns, simple in detail, appear to be the result of recent dispersal across water gaps or into newly available land (that is, ice free), as post-tertiary species attain broader ranges within today's continental configuration. These patterns, which may be complex locally, for example, Nebria (Kavanaugh, Ph.D.
thesis) and Agra (Erwin 1982, 1983), are the result of local events governed by global climatic changes. These changes include advance and retreat patterns of glacial fronts, shrinking and expansion of tropical forests, and broadening of river systems into Ria lakes (Erwin & Adis 1982). These important events in the evolution of species groups served to isolate populations long enough for genetic changes to become fixed resulting in a proliferation of species in specialist and superspecialist groups of boreal and arboreal distribution.
In total, these spatial patterns suggest that the oldest relationships among carabid taxa are centered at the equator with a north/south polarity and that younger, or more recent, relationships have varied patterns, but are centered between landmasses paired in certain ways with an east/west polarity.
The patterns also suggest that within Carabidae, groups classified at more or less the same rank share similar patterns, further suggesting that analyses of structure which led to the present carabid classification reflects age and relation- ships and the proposed classification seems to be in balance. In all age groups, there has been an attainment of dominance in most habitats. Carabid taxa, through millennia, have met all criteria set out by Darlington (see above). It is notable that only one major habitat has not been invaded and that is the desert, where Tenebrionidae are so dominant. Even so, carabid beetles have invaded desert streams and oases where water is readily available suggesting that water (or humidity) is the general limiting factor in evolution and attainment of domi- nance in this remarkable family of beetles.
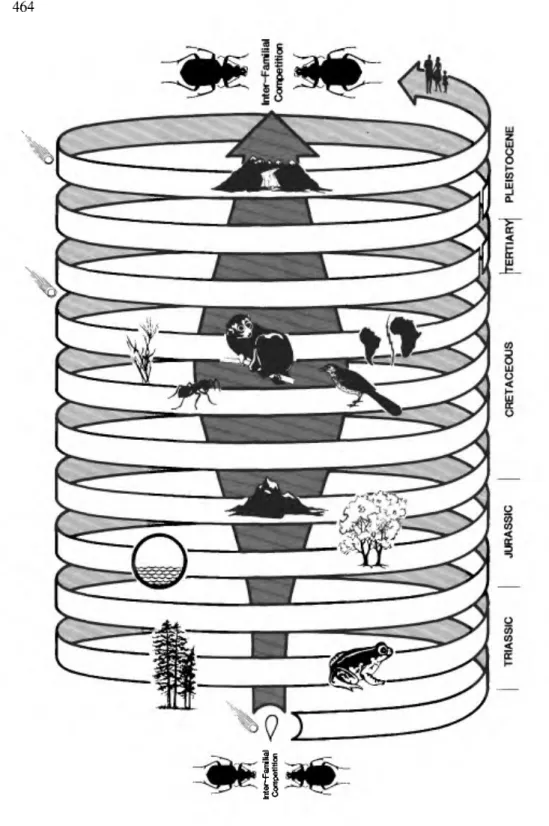
In attaining dominance, carabid lineages have been pushed by driving forces from their ancestral center in the equatorial wetlands in 'Darlingtonian waves' through the latitudes, altitudes, and into the canopy. Yet, at the same time they have maintained dominance in the tropical wetlands. The nature of the initial pushes on lineages was suggested already (Erwin 1981a) and will not be repeated here. The consequences of initial pushes leave lineages represented in the wet- lands at the same time it provides evolving forms which move from the central populations. Factors diagrammed in Figure 10 have been the driving forces pushing taxa in evolution. Adapting to these factors keeps dominance in wetland populations and drives elements to the periphery geographically and ecologically.
A measure of success is the fact that this wetland dominance must have occurred eons ago and no other insect group has replaced the carabids usurping their dominant position there.
The history of structural adaptation
Structural adaptation is the window through which systematists view phylogene- tic progression of organisms. Other windows are available now (for example, behavior, genetics, and structure of internal organs) but seldom used because data bases are not pervasive or large enough at present to provide coherent patterns. However, several structural patterns have been positively correlated with habitat and behavior in carabid beetles (Erwin 1979b) and do provide patterns. It can be assumed that this correlation also existed in primitive carabids long ago. Therefore, comparison between patterns of ancient taxa which have survived until today with younger lineages may be instructive.
For example,among all the carabid lineages regarded as ancient or old, varied color patterns or bright colors are restricted to a few groups or species whose adults live in the open on sand, salt flats, or sea beaches, for example, some Nebria, Notiokasis, Cicindis, Omophron, and some Megacephala. The remainder are somber in color; only deep metallic purple or dark metallic green are found.
However, in younger groups, bright colors and patterns are the rule, for example, many agonines, panagaeines, callistines, ctenodactylines, Galerita, Odacantha, Lebia, Calleida, and others. There is little doubt that this is correlated with adversaries which evolved color vision as a hunting ability, for example birds and lizards. Careful consideration of patterns like these and realization that each time a major event occurs, such as the rise and diversification of birds, a group of beetles such as carabids undergoes, concurrently in many of its lineages, major changes in adaptive strategy.
These kinds of major changes in the history of lineages result in new groups expanding at the expense of older groups. Among carabid beetles, there are several character systems which show progression along the same lines as color and color patterns; this progression potentially can be correlated with the events illustrated in Figure 10 and patterns detected. Aside from color evolution men- tioned above, the following list indicates some of the structural systems which must be studied in detail for the family.
1) Sclerite modifications of the ventral thorax leading progressively through time from loose to tight body rigidity for increased efficiency in wedge-pushing, running, internal moisture control, and/or flight muscle attachments. Hypotheti- cal event: adaptation to or change in (a) ground cover (for example, hard wood logs after angiosperms developed); (b) predator ability; (c) climates and soils.
2) Anterior, middle and hind tibia modifications for increased efficiency in removing soil from appendages and body parts. Hypothetical event: adaptation to or change in soils.
3) Male paramere modifications leading from long brushy margins to short glabrous margins for changes in mating speed, and male aedeagal base modifica- tions leading from open symmetrical lobes through asymmetry to basal bulb, presumably for changes in muscle attachments leading to increased efficiency in
f s
4-1 kg
Figure 10. Driving forces in the evolution of carabids with approximate time of initial impact.