Matt Mazalouskas, whose help with both technical matters during my graduate school career and whose help in editing the paper presented here were key to the successful completion of this phase. Katherine Betke whose friendship and encouragement during many long years of graduate school were essential to my survival. She always lent a helping hand and insight into the scientific aspects of my projects, as well as the occasional adult drink.
To my friends and family, your unwavering support over the years gave me the strength and determination to see it through to completion. And finally, to my loving and ever-supportive husband, Carter, there aren't really any words to convey all that you've done.
Background
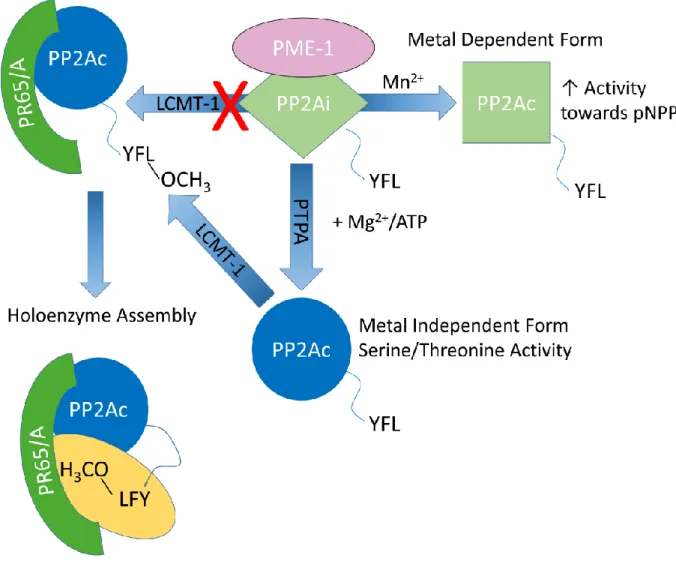
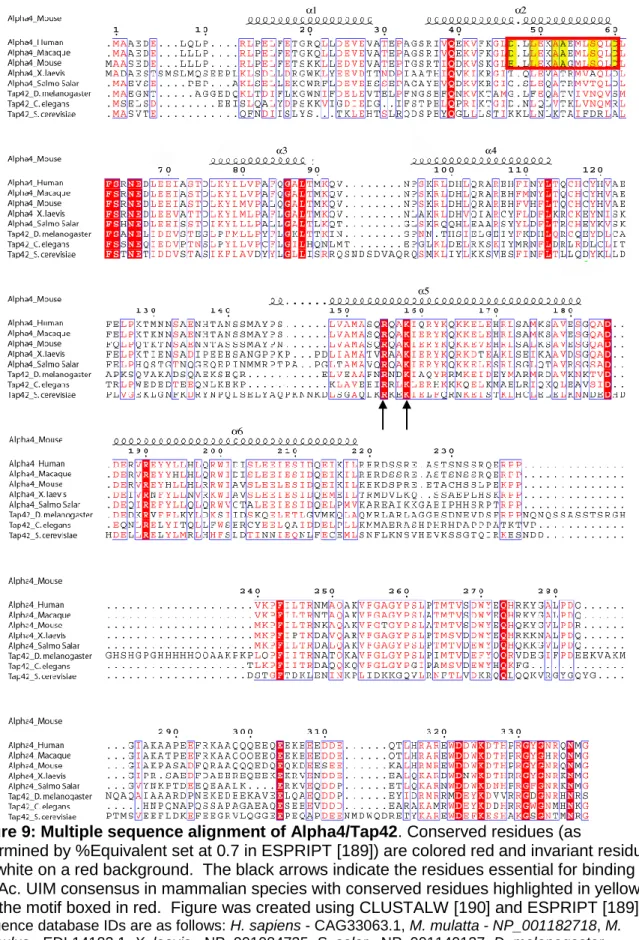
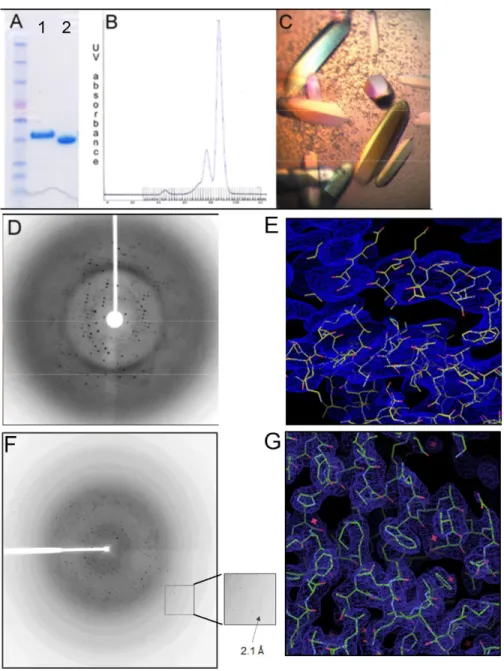
Post-translational modifications of the catalytic subunit: phosphorylation and methylation The C-terminal conserved tail of the PP2A family of phosphatases undergoes two forms of post-translational modifications, tyrosine phosphorylation and carboxymethylation [4] (Figure 3). While the importance of tyrosine phosphorylation of the PP2A family members is not clear, carboxymethylation of the C-terminal residue, L309, plays a critical role in regulating holoenzyme assembly. However, the requirements of various B-type regulatory subunits for methylation of the catalytic subunit to promote binding are controversial.
Some studies indicate that the association of B subunits (PR55) with PP2AD requires methylation [51]; while others note that the formation of this trimer is independent of methylation [8,64,65]. Two non-canonical subunits have been identified that directly interact with the catalytic subunit of all PP2A family members in the absence of canonical regulatory subunits. Alpha4/Tap42 has been shown to bind directly to all PP2A family catalysts.
These effects on apoptosis and DNA damage repair are consistent with the idea of Alpha4 being a putative oncogene and mirror many of the phenotypes seen with knockdown or overexpression of catalytic or regulatory subunits of the PP2A family.

Structural Analysis of the N-terminal PP2Ac Binding Domain of Alpha4
To determine the relative orientation of helix 5 in the absence of the crystal lattice, DEER spectroscopy was performed on Alpha4AC labeled with MTSSL at The UIM is located within helix 2 of the structure and on the opposite side of Alpha4 to the PP2Ac binding site (Figure 14A). We double labeled the protein at residues M56C, located in helix 2, and K98C, located in the main body of the protein (Figure 17A).
When Alpha4ΔC was incubated with untagged ubiquitin using any of the constructed point mutants (K98, S44, or M56) (Figure 17C), we could not detect any significant changes in the line spectra. Furthermore, the overall topology of Alpha4ΔC differs from the canonical TPR by three repeats in the arrangement of the last three helices, and these differences allow for greater flexibility (Figure 19A). Many TPR proteins also contain a terminal closing helix that widens the concave surface of the molecule [268].
Alpha4C differs from a canonical TPR repeat in the topology of the final TPR motif and the capping helix with the addition of a large loop and an inversion in orientation. This altered topology in Alpha4AC allows the opening and closing of the helix containing the PP2Ac binding residues, creating a potential PP2Ac binding site (Figure 14C). Comparison of the structure of Alpha4AC with that of its yeast homolog, Tap42, shows the extended helix containing the PP2Ac binding determinants that exist in multiple conformations, indicating that the PP2Ac binding region is flexible.
The structure of Tap42 was determined in two conformations and Alpha4 adopts a third conformation, with most of the variation among the three structures occurring in the relative position of the extended helix containing the PP2Ac binding residues (Figure 14B). The Alpha4 structure also has the critical PP2Ac binding residues oriented to face the concave side of the molecule (Figure 14). This region of the protein has been reported to be important for Alpha4 regulation of PP2Ac polyubiquitination.
Role of the PP2Ac and E3-Ubiquitin Ligase Binding Domains
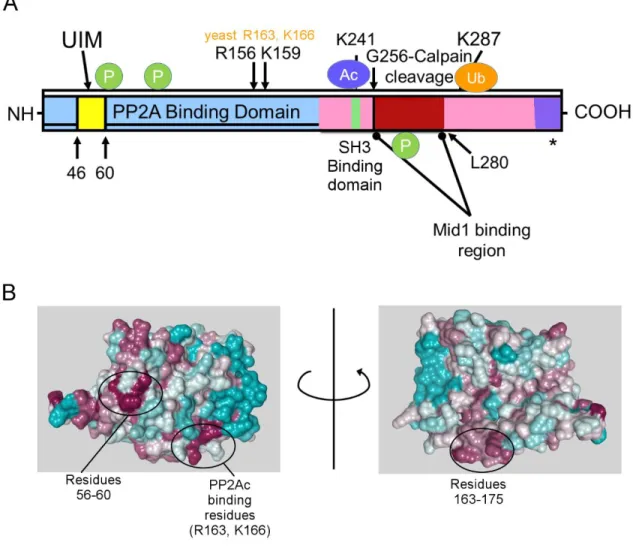
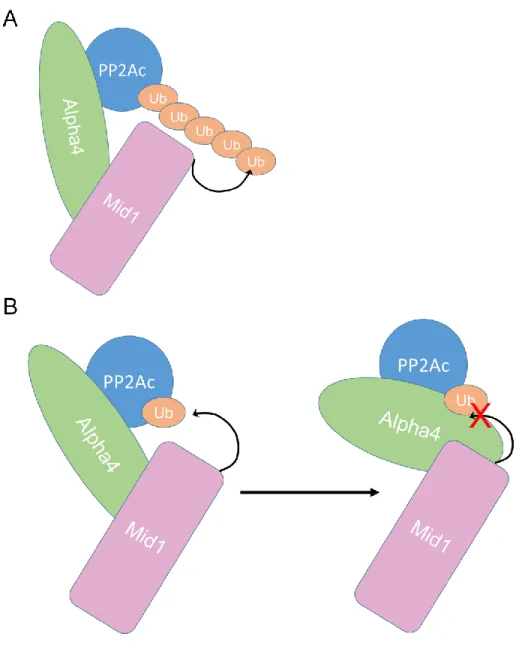
Alpha4 co-localizes with Mid1 in cells, with the C-terminus of Alpha4 and the B-Box1 domain of the Mid1 protein mediating the association. In this chapter, we investigate the role of the E3 ubiquitin ligase and PP2Ac binding domains of Alpha4 in the regulation of PP2Ac ubiquitination and degradation. Both the Mid1 binding domain and the PP2Ac binding residues of Alpha4 are essential for regulation of PP2Ac polyubiquitination.
To investigate the role of the Mid1-binding domain and PP2Ac-binding residues of Alpha4 in protecting PP2Ac from polyubiquitination, we performed TUBE isolation experiments on cell lysates of cells transfected with HA3-PP2Ac and with empty vector, full-length FLAG- FLAG-Alpha4, Alpha4ΔC, or FLAG-Alpha4_ED. These findings indicate that both the Mid1-binding domain and the PP2Ac-binding residues of Alpha4 are essential for Alpha4-mediated protection of PP2Ac from polyubiquitination. Both the Mid1 binding domain and the PP2Ac binding residues of Alpha4 are required for Alpha4 to protect PP2Ac from degradation.
These results indicate that both the Mid1 binding domain and PP2Ac binding residues are essential for the stabilizing effect of PP2Ac Alpha4. Inhibition of deubiquitination by PR-619 leads to increased polyubiquitination in the presence and absence of Alpha4. Differential effects of alpha4 knockdown and expression on the expression of protein phosphatase 2A family members.
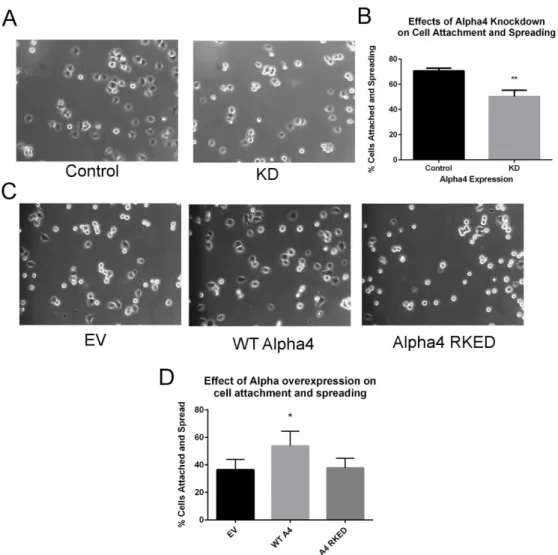
Studies of the effects of Alpha4 overexpression or knockdown on endogenous levels of PP2Ac have been inconclusive. Based on these results, we sought to determine the effects of Alpha4 knockdown and overexpression on PP2Ac, PP4c, and PP6c expression levels. To test the effects of both transient and stable knockdown and expression, we developed a lentiviral-based panel of Alpha4 knockdown cell lines.
Also examine the ability of Alpha4 and Alpha4 mutants to rescue the knockdown. Flag-Alpha4 cDNA and scrambled shRNA plus human Flag-Alpha4 cDNA were created in HEK293T cells and expression levels of Alpha4 were compared by Western analysis of cell lysates (Figure 26B).

Alpha4 Effects on Cell Adhesion, Growth, and Viability
The effects of Alpha4 on other members of the PP2 family have not been extensively studied. Previous studies presented in this work (Chapter 4) investigate the effects of knockdown and expression of Alpha4 on PP2Ac, PP4c and PP6c and we found that Alpha4 has different effects on PP2A family members. Stable knockdown of Alpha4 results in a significant reduction in PP4c and PP6c expression levels, while having minimal effect on PP2Ac expression levels.
The ability of Alpha4 to interact with PP2A family phosphatases is required for Alpha4 effects on cell adhesion. Expression of the PP2Ac binding-deficient mutant of Alpha4 leads to increased cell death in HEK293T cells. The known relationship between cell growth, apoptosis, and Alfa4 led us to explore the possibility that differences in cell growth among viable HEK293T cells were due to underlying differences in rates of cell death or cell proliferation.
Characterization of this cell line revealed that, unlike the stable HEK293T cell lines, knockdown of Alpha4 had a significant negative effect on cell growth (Figure 33B,C). The effects of Alpha4 overexpression on the response of the lung cancer cell line A549 to two different cell stressors, hydrogen peroxide (H2O2) and cycloheximide (CHX), were investigated. Contrary to our hypothesis, overexpression of Alpha4 led to a dramatic increase in cell death of A549 cells exposed to H2O2 (Figure 35).
Overexpression of alpha4 led to reduced cell death compared to WT A549 cells when cells were treated with CHX, consistent with the idea that alpha4 plays a role in protecting cells from apoptosis (Figure 36). Accordingly, we investigated whether expression of the RKED mutant had different effects on the expression of PP2Ac, PP4c, and PP6c to elucidate which of the PP2A family members was involved in the proliferative effects of Alpha4 cells. This is in contrast to a number of studies performed in various cell lines showing that increased expression of Alpha4 increases the rate of cell proliferation and decreases cell apoptosis [223,238].
Summary and Future Directions
Carboxymethylation of the catalytic subunit of Protein Phosphatase 2A in insulin-secreting cells: Evidence for functional consequences on enzyme activity and insulin secretion. The structure of the protein phosphatase 2A PR65/A subunit reveals the conformation of its 15 tandemly repeated HEAT motifs. A functional role for the B56 alpha subunit of protein phosphatase 2A in ceramide-mediated regulation of Bcl2.
Methylation of the protein phosphatase 2A catalytic subunit is essential for association of Bα regulatory subunit but not SG2NA, striatin, or polyomavirus center tumor antigen. Identification and Functional Analysis of Two Ca2+-Binding EF-Hand Motifs in the B"/PR72 Subunit of Protein Phosphatase 2A. Purification of Pig Brain Protein Phosphatase 2A Leucine Carboxyl Methyltransferase and Cloning of the Human Homolog.
The Saccharomyces cerevisiae phosphotyrosyl phosphatase activator proteins are required for a subset of the functions disrupted by protein phosphatase 2A mutations. Monoubiquitination promotes calpain cleavage of protein phosphatase 2A (PP2A) regulatory subunit α4, altering PP2A stability and microtubule-associated protein phosphorylation. Overexpression of mTOR alpha4 phosphoprotein activates protein phosphatase 2A and increases Stat1α binding to PIAS1.
B cell receptor-associated protein alpha4 exhibits rapamycin-sensitive binding directly to the catalytic subunit of protein phosphatase 2A. The structure of the tetratricopeptide repeats of protein phosphatase 5: Implications for TPR-mediated protein-protein interactions. Phosphorylation and microtubule association of the Opitz syndrome protein mid-1 is regulated by protein phosphatase 2A via binding to the regulatory subunit alpha 4.
The E3 ubiquitin ligase and protein phosphatase 2A (PP2A) binding domains of the Alpha4 protein are both required for Alpha4 to inhibit PP2A degradation. Essential roles of the tap42-regulated protein phosphatase 2a (PP2A) family in Drosophila melanogaster imaginal disc development.