INTRODUCION
Epigenetics
These changes can persist through cell divisions for the rest of the cell's life and can also last for several generations.
Terms used
DNA Methylation……………………………………………..................... 03-04
The mechanisms underlying these changes remain unknown; however, these data suggest that early changes in the DNA methylation machinery accompany and may directly participate in the transformation process. Function: Required for genome-wide de novo methylation and essential for establishing DNA methylation patterns during development. Function: Required for genome-wide de novo methylation and essential for establishing DNA methylation patterns during development.
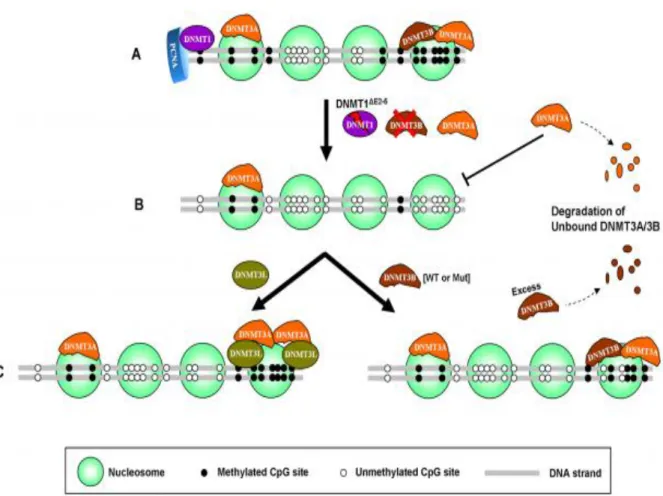
May act as a transcriptional co-repressor by associating with CBX4 and independently of DNA methylation. When DNA methylation is lowered by genetic disruption of DNMT1 and DNMT3B in DKO cells, DNMT3A loses its ability to bind to nucleosomes, resulting in destabilization and subsequent degradation of the protein. Restoration of DNA methylation in such hypomethylated cells through expression of exogenous DNMT3B (WT or mut) or DNMT3L increases DNMT3A protein levels by enabling it to bind to nucleosomes again, resulting in stabilization of DNMT3A protein.
5-Aza-2'-deoxycytidine (5-Aza-CdR) inhibits DNA methylation and is used in vitro to induce the re-expression of genes thought to be silenced by promoter methylation. DNMT3B proteins affect DNA methylation in cancer cells and DNA methylation to control gene expression. To study the effect of DNMT3B7 expression on the DNA methylation patterns and phenotypes of cancer cells.
DNA methylation is involved in many normal cellular processes and is abnormally distributed in cancer cells, leading to some of the phenotypes of cancer cells. The cancer cells express shortened forms of DNMT3B, one of the enzymes that carry out the DNA methylation reaction, and we hypothesize that truncated DNMT3B proteins contribute to abnormal DNA methylation patterns in cancer cells. The effect of DNA methylation inhibitor 5-aza-2-deoxycytidine on the cell cycle and apoptosis.
It is important in the regulation of promoter methylation status and the expression of tumor suppressor genes. Tumor development is epigenetic changes; that is, a change in DNA methylation patterns that induces tumor suppressor gene silencing. According to one scenario, DNA methyltransferase (DNMT), which is responsible for DNA methylation, represents the main epigenetic maintenance and turnover.
DNA methylation alternations also occur in the precancerous stage before the establishment of cancer and determine the clinicopathological characteristics of developing malignancies. Lucy A Godley: University of Chicago Location: USA LABOME.ORG Role of DNMT3B in DNA Methylation of Cancer Cells.

Hepatocellular Carcinoma can be prevented by Inhibition of denovo
DNA methylation in breast cancer
DNMT3B polymorphisms and risk of primary lung cancer
Polymorphisms and haplotypes of the DNMT3B gene may influence DNMT3B activity on DNA methylation, thereby modulating lung cancer susceptibility. The DNMT3B gene has two transcriptional start sites located in different exons (exon 1A and 1B) and its expression is regulated by different promoters. One promoter is embedded in a CpG-rich region, whereas the other promoter is found in CpG-poor.
In this study, carriers with the -283T and -579G alleles were at reduced risk of lung cancer compared to individuals with the -283C and -579T alleles. In order to determine whether the association between the DNMT3B −283T > C and −579G > T polymorphisms and the risk of lung cancer is due to a difference in the transcriptional activity of the DNMT promoter, we compared the promoter activity of the wild-type allele or polymorphic allele of these two polymorphisms by luciferase assay. Since the −283T > C and −579G > T polymorphisms were in LD, the functional effects of the two polymorphisms may be haplotype dependent.
We found that the polymorphisms −283T>C and −579G>T in the DNMT3B promoter, and their haplotypes, were significantly associated with the risk of lung cancer, especially AC. DNMT3B sequence variants and their biological function are also necessary to understand the role of DNMT3B polymorphisms in determining lung cancer risk. Genetic polymorphisms often vary between ethnic groups; Further studies are needed to clarify the association of the DNMT3B polymorphism with lung cancer in diverse ethnic populations (Lee, et al., vol 26).
The Role of DNMT3B in the DNA Methylation of Cancer Cells
The DNA within a cell can be modified by methylation to change its structure and affect gene expression. The cellular pathways found to mediate effects of truncated DNMT3B proteins reveal paradigms common to other processes involving DNA methylation, such as mammalian embryonic development, X-chromosome inactivation, genomic imprinting, and aging.
DNMT3b gene expression in Human biliary tract carcinoma cell due to
Expression of m RNA and protein of the target gene can be reduced by antisense technology. Construction of the antisense DNMT3B gene eukaryotic expression plasmid using liposome and transfected into the human bile duct carcinoma cell line QBC-939. Semi-quantitative RT-PCR was used to observe the effect of transfection with pcDNA. DNMT3B decreased the expression level of DNMT3B the expression level of DNMT3B gene m RNA IN qbc-939.
Found that transfection with pcDNA-DNMT3B decreased the expression level of DNMT3B protein in QBD-939 (Zuo, et al., 2006).
DNMT potential and its Epigenetic Regulation for Lung Cancer..................... 15-16
The hypermethylation of TSG is a common thing in lung cancer, and it is widely recognized that DNMT1 is correlated with hypermethylation in the TSG promoters, especially among smoking SCC patients (Suzuki et al., 2004). The proposals of epigenetic biomarker, epigenetic silencing, methylation profile, histone coding, reflect the investigation of epigenetic regulation in lung tumorigenesis (Tang et al., 2009). DNMTs for the hypermethylation of the promoter of the target genes we need to measure the expression level of the specific gene of the enzyme.
Thus, our aim was to compare the expression level of DNMT3A and DNMT3B in cancer tissue and normal tissue.
MATERIALS AND METHODS……………………………………………. 18-20
Then Nano drop 1000 was used to measure the concentration by setting the sample type setting to Second sample and the constant to 33 and stored at -80°C. After isolation of the total RNA from normal blood and cancer tissue, we checked their concentration by taking its OD in spectrophotometer, in case of gall bladder the concentration was found to be very low ie. because it took a long time to process after sample collection and also cDNA was not synthesized from the total RNA by the rt-PCR method due to an unsuitable condition.
286.69 and 301.58 respectively in normal blood cells, but in cancer tissue it was found that the concentration increases, i.e. After gene-specific amplification, the concentration of DNMT3A and DNMT 3B was observed to be expressed in lymph node cancer tissue. In many research articles, we have found that DNMT3B is more expressed in cancer tissue, but in lymph node cancer, DNMT3A is shown to be overexpressed compared to DNMT3B.
So from this we can assume that these overexpression of DNMT3A leads to hypermethylation of TSG (for example p53), which in turn decreases the expression of p53 and causes the cancer. We also know that the expression of EZH2 shows the methylation of the histone proteins. When the expression of DNMT 3A and 3B is more, the methylation occurs and due to this methylation in DNA and also Histone proteins, the overall cell methylation increases, leading to the suppressive function of the Tumor Suppressor gene.
As a result, the number of tumor-forming cells will increase, which ultimately leads to the development of cancer. This is the reason why the concentration of the p53 gene (TSG) decreased in the lymph node cancer cell that we used for our work. Therefore, we can assume that due to the increased expression of Denovo methyltransferases, the methylation of the promoter of tumor suppressor genes increased and thus they can be suppressed.
Therefore, the cell undergoes rapid cell divisions that can ultimately lead to tumor formation and the development of cancer. Interference with protein binding at AP2 sites by sequence-specific methylation in the late E2A promoter of adenovirus type 2 DNA.