I hereby declare that the research included in this thesis is the result of experiments conducted at the Department of Biological Sciences and Bioengineering, Indian Institute of Technology Guwahati, India under the supervision of Dr. cloacal droplets from 2-week-old chicks 96 hours post-infection in different experimental groups propagated in 9-day-old embryonated chicken eggs. Haemagglutination test of virus secreted in oral and cloacal droplets of two-week-old chickens 5 days after infection in different experimental groups, multiplied in 9-day-old embryonated chicken eggs.
INTRODUCTION
- Newcastle disease
- Aetiology
- Epidemiology
- Transmission
- Pathotypes
- Geographical distribution
- Diagnosis
- Prevention and control
- Rationale for the study
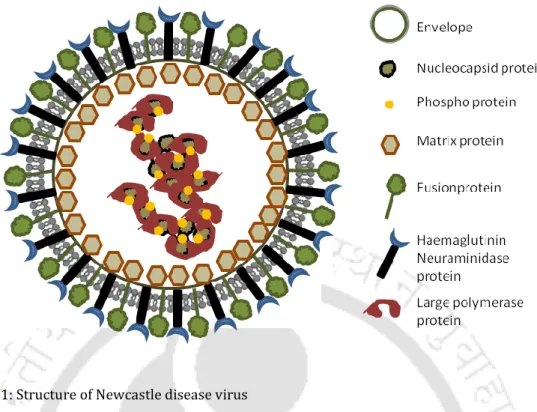
It is a member of the family Paramyxovidae in the genus Avulavirus and species Avian Paramyxovirus serotype 1. Based on the severity of the disease caused in birds and the degree of morbidity and mortality, NDV strains are grouped into five pathotypes (Alexander and Senne 2008). . Such a scenario requires extensive research to understand the molecular pattern of the disease in the country so that appropriate control measures can be taken.
OBJECTIVES
According to the OIE, ND is considered the fourth most devastating disease in poultry after highly pathogenic avian influenza, avian infectious bronchitis and low pathogenic influenza (Anonymous 2011). The development of cost-effective and efficient diagnosis remains another challenge in the prevention and control of NDV. The development of antiviral therapeutics against this disease could overcome deficiencies in vaccination practices, particularly in developing countries with limited basic infrastructure.
REVIEW OF LITERATURE
- Outlook on Newcastle disease: Global scenario
- Outlook on Newcastle disease: Indian scenario
- Newcastle disease virus
- Structure and genome organization of Newcastle disease virus
- Replication of Newcastle disease virus
- Molecular characterization of Newcastle disease virus
- Diagnosis for Newcastle disease
- Prevention and control of Newcastle disease
- Antivirals
The protein has also been shown to be involved in the production kinetics of the virus (Jang, Hong et al. 2010). The NDV F protein has been related to the pathogenicity of the virus (Heiden, Grund et al. 2014). Label-free biosensors are among the emerging diagnostic tools of the century (Sang, Wang et al. 2016).
- Abstract
- Introduction
- Materials and methods
- Eggs and cells
- Virus isolation and plaque purification
- Clinico-pathological assays
- RNA extraction and sequencing
- Phylogenetic and evolutionary analysis
- Results
- Biological characterization
- Genetic characterization of the NDV strain Kamrup
- Phylogenetic and evolutionary characterisation of NDV strain
- Independent evolution of NDV strain Kamrup
- Discussion
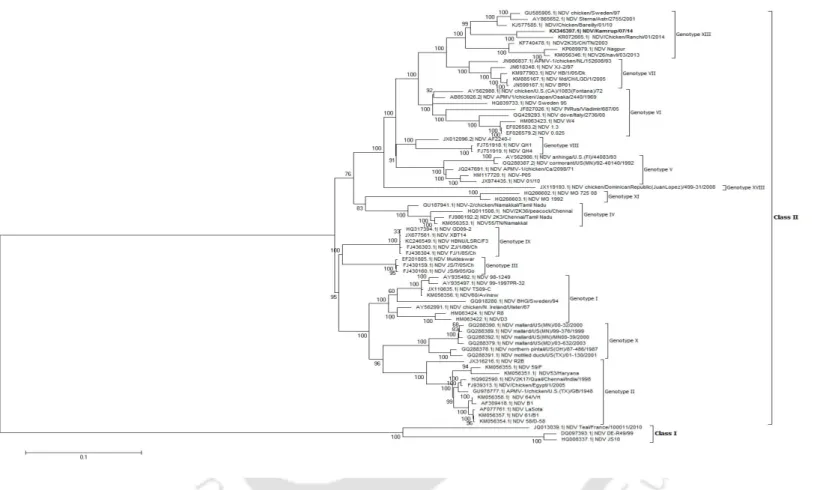
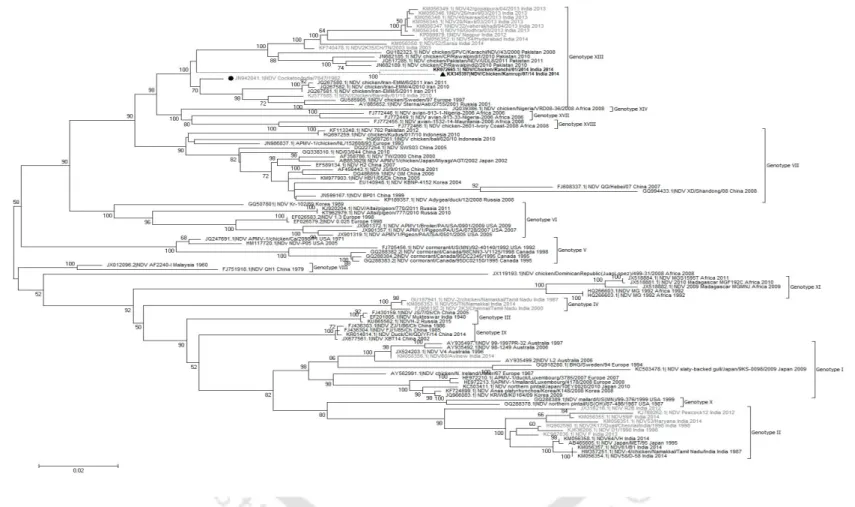
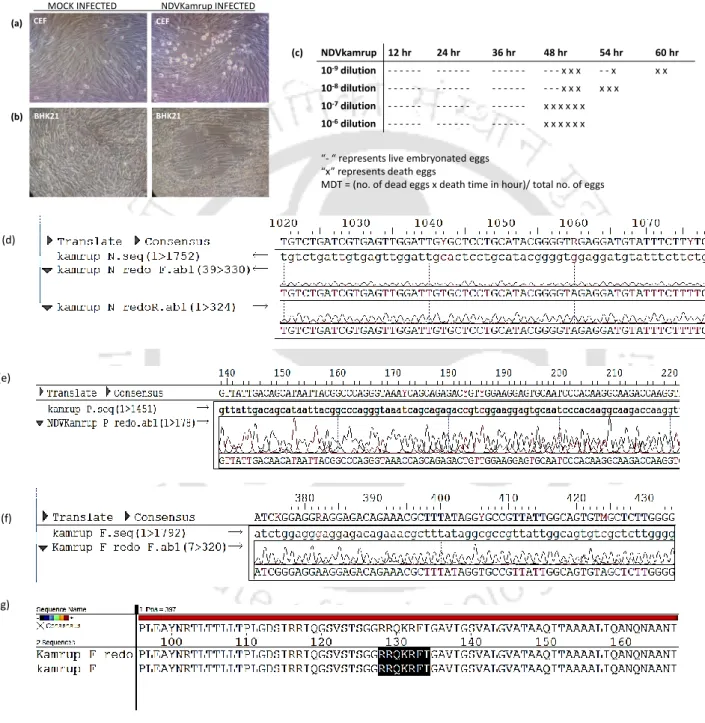
NDV strain Kamrup formed a new subgenotype XIIIc together with two geographically distinct NDV strains. In addition, the sequence of the entire F gene was used to determine the phylogenetic relationships of NDV strain Kamrup with other representative genotype XIII strains (total n = 40). The complete genome of NDV strain Kamrup was submitted to GenBank (accession number KX345397).
The nucleotide sequence and the deduced amino acid sequence of the F and HN proteins of NDV strain Kamrup were compared with available GenBank sequences (Table 2.2b). In addition, the evolutionary distance of complete genome of NDV strain Kamrup showed minimum 8.9% distance with sub-genotype XIIIa strains. The NDV strain Kamrup showed its clustering with the strains of genotype XIII with confident bootstrap value of 100.
Phylogenetic and evolutionary analysis of the NDV strain Kamrup showed encouraging results for its placement in an evolving clade among genotype XIII viruses. Clinicopathological evaluation of the NDV Kamrup strain showed typical viscerotropic virulent signs causing severe hemorrhagic lesions in the kidneys. The determined amino acid sequence of the F and HN proteins of the NDV Kamrup strain showed significant substitution mutations with other representative strains of genotype XIII.
We proposed to conserve the NDV Kamrup strain in a new subgenotype XIIIc with other Indian strains.
- Introduction
- Materials and methods
- Phylogenetic and evolutionary divergences analysis using nucleotide
- Determination of T cell epitopes in NDV phosphoprotein
- Cells and Virus
- Cloning and expression of the NDV P gene into a prokaryotic vector
- Cloning and expression of the NDV P gene into a eukaryotic vector 61
- Transfection
- Real-time analysis of innate immune molecules
- Results
- Phylogenetic and evolutionary analysis of NDV using P gene
- CTL epitopes in NDV phosphoprotein
- Cloning and expression of recombinant phosphoprotein
- Regulation of innate immune molecules on NDV infection and NDV P
- Discussion
In another study, high avidity CTL response against a P protein epitope of Paramyxovirus Simian virus 5 was reported (Gray, Parks et al. 2001). IRF7 has been initially identified to induce the expression of IFNs and IFN-stimulated genes (ISGs) in the hosts (Marie, Durbin et al. The phylogenetic analysis was performed using MEGA6 software (Tamura, Peterson et al. 2011) with standard errors as calculated from 500 bootstrap replicates.
The initial tree(s) for the heuristic search were obtained by applying the Neighbor-Joining method to a matrix of pairwise distances estimated using the Maximum Likelihood Composite method (Tamura, Stecher et al. 2013). The P protein sequence from lentogenic NDV LaSota and velogenic NDV/Kamrup/07/14 was assessed for the presence of cytotoxic T lymphocyte (CTL) epitopes using NetCTL1.2 software (Larsen, Lundegaard et al. 2007). Next, cDNA was prepared using the specific forward primer P gene, designed based on the whole genome sequence of NDV strain LaSota (GenBank accession number: JF950510) (P for: 5´-CTAGCTAGCATGGCCACCTTTACAGATG-3´).
Further, cDNA was prepared using a P gene-specific primer designed on the basis of the whole genome sequence. In this study, we assessed the genetic diversity of NDV (n = 60) based on the full-length sequence of the P gene. Induction was prominent in the later phase of the virus life cycle, when overexpression of the P protein was observed.
However, this may also be due to the expression of the IFN antagonist protein V in the early life stages of the virus.

- Abstract
- Introduction
- Materials and Methods
- Cloning and expression of NDV P protein in prokaryotic system
- Polyclonal antibody production in chicken
- Chicken serum samples
- Formulation of ELISA
- Positive negative threshold and determination of standard curve
- ELISA sensitivity, specificity and accuracy
- Results
- PNT curve analysis and determination of single serum dilution for
- Sensitivity, specificity and accuracy of the recombinant P protein
- Discussion
In this study, a recombinant P-based ELISA was developed to evaluate its effectiveness in detecting NDV-specific antibodies in various serum samples and generate a rapid and effective diagnostic tool for NDV infection in poultry. In addition, 50 serum samples were collected from healthy birds vaccinated with the LaSota strain and showing no signs of the disease. The collected serum samples were screened negative for avian influenza and salmonella infection (IDEXX, USA).
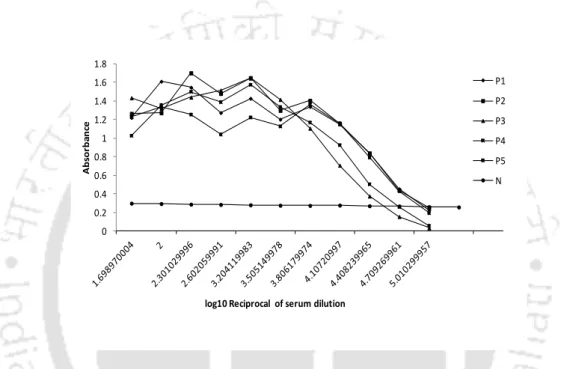
NDV-specific positive and negative serum samples were obtained from Khanapara veterinary college (Assam, India). The negative serum samples were used to construct the positive-negative threshold (PNT) baseline as shown previously (Mohan, Dey et al. 2006). The positive negative threshold (PNT) line was calculated using ten negative serum samples that were screened negative with HI.
The PNT baseline with different positive serum samples was presented diagrammatically and single serum dilution was calculated by linear regression (Figure 4.2a). ELISA using the single serum dilution method was performed using 50 serum samples (randomly selected) from collected lot of 300 samples. Thus, 1:100 dilutions of serum samples were further considered to predict the titer of remaining serum samples.
In the present study, pooled serum samples from different vaccinated herds and farms showed a concordance of 99% by single dilution ELISA and HI (kappa value = 0.992).
- Abstract
- Introduction
- Materials and Methods
- Viruses, cells and compound
- Infectivity assays and virus titration
- In-ovo study
- Chicken experiment
- Analysis of innate immune response to imidazole treatment
- Results
- Cytotoxicity assay of imidazole and albendazole
- Antiviral activity of imidazole and albendazole
- In ovo antiviral activity
- Antiviral activity of imidazole derivative in chickens
- Upregulation of innate immune response on imidazole treatment
- Discussion
Vaccination and biosecurity measures remain the only available preventive choices (Ganar, Das et al. 2014). The mNDV titer was quantified by plaque assay in BHK21 cells as described previously (Krishnamurthy, Huang et al. 2000). Western blot experiment was performed for expression analysis of the viral protein in the different experimental sets (Nayak, Kumar et al. 2010).
In the first experiment, the expression of type I interferon (IFN) was checked on mNDV-infected CEF cells treated with 30 mM imidazole and compared with mock-infected and virus-infected cells. The graphical representation of the reduction in viral titers in the plaque assay and TCID50 experiments is shown in Figure 5.2d. A two-fold reduction in mNDV titer was observed in the egg treated at the post-entry, co-entry and pre-entry steps compared to the mNDV-infected control.
The twofold reduction in the mNDV particles was supported by the HA titer and plaque titer using infected allantoic fluid (Figure 5.3c). The amount of mNDV excreted into the environment by vaccinated birds is considered an important parameter for vaccine efficacy (Dortmans, Peeters et al. 2012). The difference in the antiviral effect in cellular and living models can also be attributed to the variation of viral kinetics in those systems (Jogler, Hoffmann et al. 2006).
In the present study, the efficacy of imidazole on the impedance of NDV replication was shown to be dose dependent.
- Abstract
- Introduction
- Materials and methods
- Cells and Virus
- Cell toxicity assay
- Virus infectivity in time of addition assay
- Intracellular acridine orange assay
- Endosomal trapping and pH rescue experiments
- Glucose uptake assays
- Nutrient starvation and glycolytic inhibitor studies
- Real-time RT-qPCR analyses of factors involved in glucose
- In-ovo experiment
- In-vivo experiment
- Results
- Cell viability and Infection under niclosamide treatment
- Reduction in NDV replication in the presence of niclosamide in vitro
- Niclosamide inhibits NDV entry
- Antiviral efficacy of niclosamide co-tracks with inhibition of cellular
- Reduction in NDV replication in the presence of niclosamide in ovo
- Reduction in NDV replication in the presence of niclosamide in vivo
- Discussion
Antiviral potency of niclosamide is correlated with its protonophore activity in the endosomes and blocking of metabolic adaptation necessary for efficient viral propagation. The virus titer was determined by capturing GFP fluorescence in the cells 48 hours after infection (Das, Baro et al. 2019). For the estimation of glucose flux in the presence of niclosamide, 3x106 DF-1 cells were seeded in 6 well plates (Matrix, Thermo Scientific) and incubated with 2 μM.
The amount of glucose and lactose in the supernatant was then measured using the standard HPLC protocol (Wannet, Hermans et al. 2000). The presence of mNDV in the smears was determined by inoculating it into fertilized chicken eggs and performing an HA test. A fold reduction in mNDV titer was observed in the egg treated at the post-entry, co-entry and pre-entry steps compared to the mNDV-infected control.
The fold reduction in the mNDV particles was supported by plaktites using infected allantoic fluid (Figure 6.5b). RT-PCR of mNDV-infected DF-1 cells treated with 2 μM niclosamide at pre-, co-entry and post-entry steps (b). Fold change in mNDV genome copies in the presence of glycolytic inhibitor sodium oxamate in DF-1 cells (g).
Modifications in the chemical structure of niclosamide to generate a prodrug may prove useful in promoting its physicochemical interference for the treatment of respiratory viral infections.
34;Biological activities of monoclonal antibodies against the haemagglutinin-neuraminidase (HN) protein of Newcastle disease virus." 34;A dot immunoblotting assay (dot blot ELISA) for early detection of antibodies against the Newcastle disease in chickens." 34; Newcastle Disease Virus in little owls (Athene noctua) and African penguins (Spheniscus demersus) in an Israeli zoo.
34;Overexpression of Newcastle disease virus (NDV) V protein increases NDV production kinetics in chicken embryo fibroblasts.". 34;Complete genome sequence of a Newcastle disease virus isolated from wild peacock (Pavo cristatus) in India." Genome Announc 2(3). 34; Genetic characterization and pathogenicity assessment of Newcastle disease virus isolated from wild peacock." Virus Genes.
34;Detection and Molecular Characterization of Newcastle Disease Virus in Peacocks (Pavo cristatus) in Haryana State, India.". Biological. 34; Antigenic Characterization of Newcastle Disease Virus Intrinsic Proteins by Monoclonal Antibodies." Virus Res.
34;Loss of N-linked glycosylation of the hemagglutinin-neuraminidase protein alters virulence of Newcastle disease virus." J Virol. 34;[Full-length genome analysis of two genotype III velogenic Newcastle disease virus strains reveals their close relationship to vaccine Mukteswar] ." Wei Sheng Wu Xue Bao. 34; Identification of the P proteins and other disulfide-linked and phosphorylated proteins of Newcastle disease virus." J Virol.