The enzyme activity assays of the recombinant protein were performed with the substrates cytosine and the prodrug 5-FC. Furthermore, the efficient purification and activity assays of the recombinant CD-UPRT fusion protein were demonstrated.
CSF Granulocyte-colony stimulating factor HSV Herpes Simplex Virus
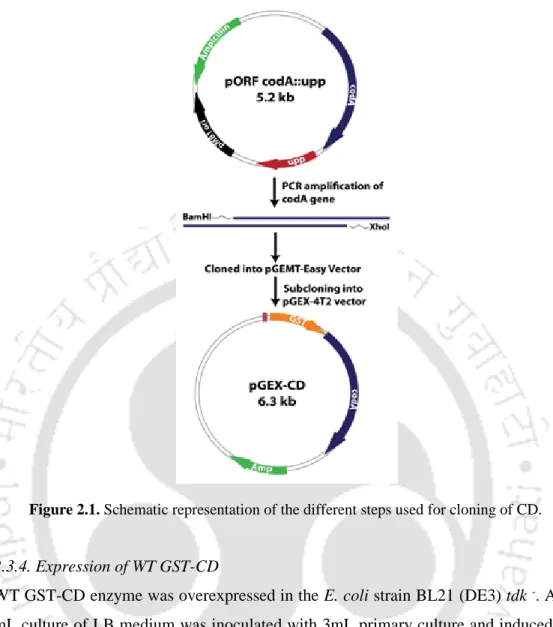
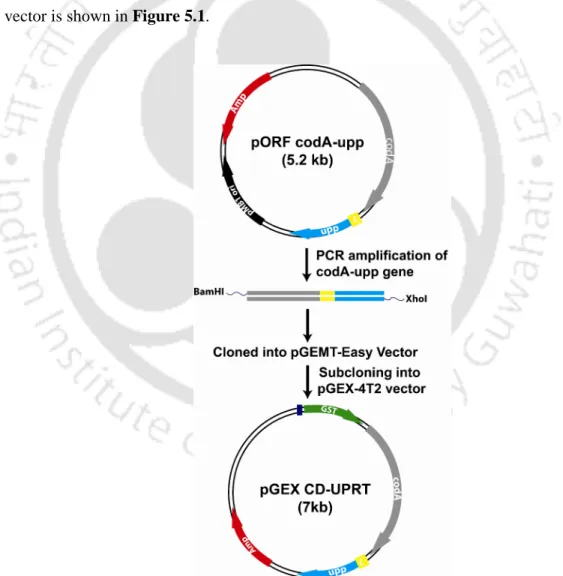
Schematic representation of the cloning steps involved in the construction of the pGEX CD-UPRT vector. SDS-PAGE showing the solubilization of recombinant GST-tagged CD-UPRT protein using different concentrations of sarkosyl.

INTRODUCTION AND LITERATURE REVIEW
- Introduction
- Recombinant DNA technology approaches for therapeutic proteins
- Genetic engineering approach to redesign gene(s) to improve functions of recombinant enzymes
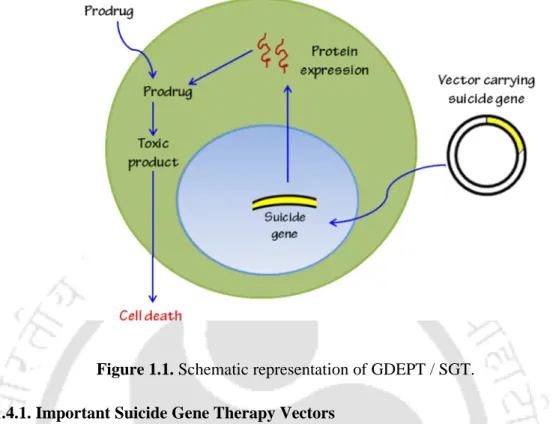
- Fundamentals of Suicide Gene Therapy
- Important Suicide Gene Therapy Vectors
- Prospects of Non-Viral Vectors
- Major Suicide Gene Therapy Approaches
- Expression of Suicide Genes
- Applications of Bioinformatics tools in the genetic engineering of enzymes Computational approaches such as homology modeling and molecular docking are
- Key Areas and Scopes
- The Present Work 1. Objectives
- Significance and Salient Features of the Present Thesis Work The significance and salient features of the present study are given in below
Salman et al., (2010) used site-directed mutagenesis to investigate the effects of Ser 348 phosphorylation on the activity and cellular localization of human SULT2B1b, a major isoform of the human cytosolic sulfotransferase2 (SULT2) family. Takahashi et al., (2012) identified and characterized novel homologues of RPE65 (retinal pigment epithelium isomerohydrolase), RPE65c and 13-cisisomerohydrolase (13cIMH).
CLONING, EXPRESSION, PURIFICATION AND FUNCTIONAL ACTIVITY ASSAYS OF E.COLI CYTOSINE
- Introduction
- Outline of the Research Work
- Experimental Section 1. Materials
- Results and Discussion
- Conclusion
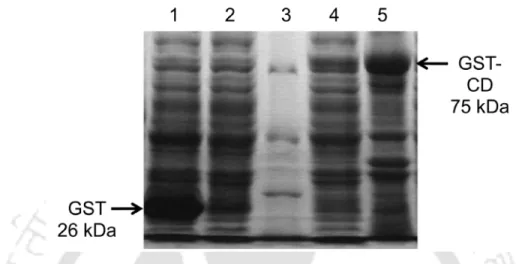
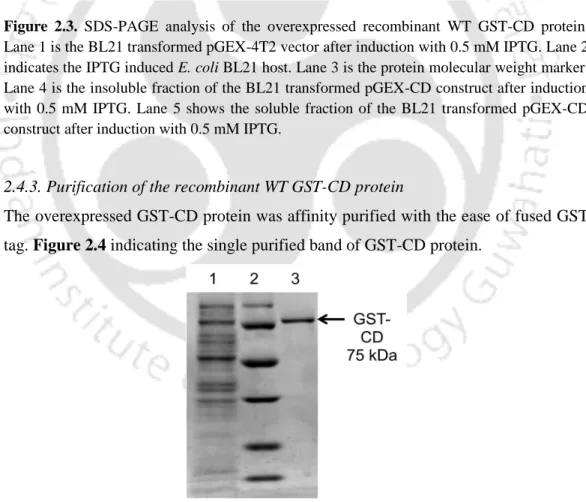
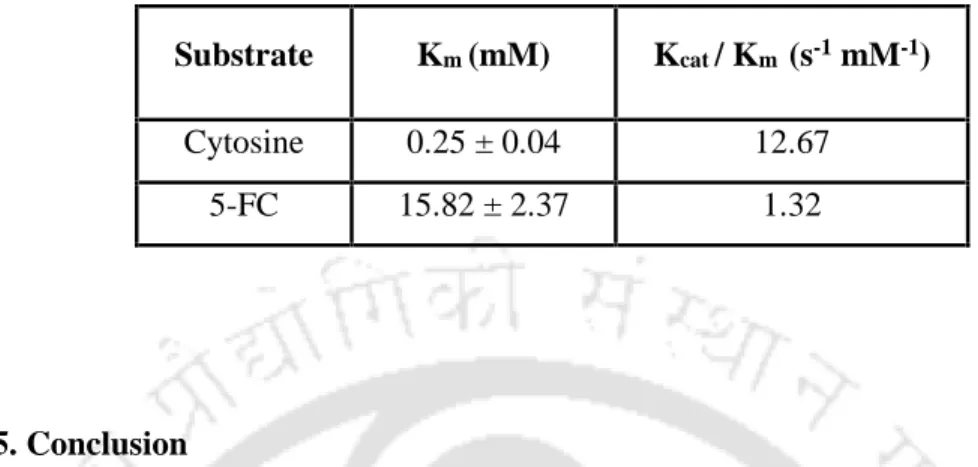
The WT GST-CD enzyme was purified to near homogeneity by affinity column chromatography using immobilized glutathione-agarose beads. The protein concentration of the WT GST-CD enzyme was determined using the Bradford method using BSA as a standard. Kinetic values for WT GST-CD were obtained with 5-FC substituted with cytosine as substrate.
Overexpressed GST-CD protein was affinity purified with an easily fused GST tag. The kinetic data obtained for the WT GST-CD enzyme with the substrates cytosine and 5-FC are shown in Table 2.1. Kinetic parameters obtained using double reciprocal plots for recombinant WT GST-CD enzyme when assayed with cytosine and 5-FC.
SDM) Approaches to create Novel E. coli Cytosine Deaminase Mutants
IN SILICO AND IN VITRO SITE DIRECTED MUTAGENESIS (SDM) APPROACHES TO CREATE
Introduction
Therefore, the in silico-based approaches could be used to generate novel CD mutants with increased specificity for the prodrug, which could serve as better candidates for potential SGT applications.
Outline of the Research Work
Experimental Section
The structure of the cytosine and 5-FC ligands were drawn using Java Molecular Editor (JME) (Peter, 2006) and transferred to PRODRG (Schüttelkopf et al., 2004), a small molecule topology generator to convert ligands into format required for docking studies. The results were analyzed based on the known interactions between the macromolecule and the ligands, as well as the binding free energy of the ligands. The double reciprocal plot was used to determine Km values for each mutant and WT CD enzyme.
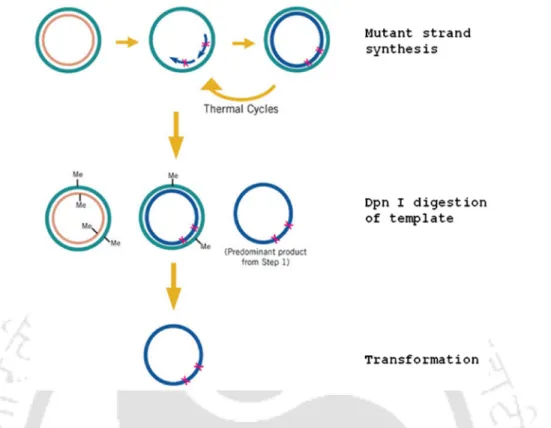
The coding region of both WT CD and its S126R variant were PCR amplified from their pGEX-4T2 vectors using the gene-specific primers 5'CGTCCATGGGAATGGTGTCGAATAACGC3' and. After sequence analysis, purified recombinant plasmids containing WT CD and mutant genes encoding S126R were transfected into human cervical cancer cells (HeLa) using electroporation according to the manufacturer's protocol. For MTT cell viability assays, HeLa cells expressing WT CD and S126R genes were seeded in 0.1 mL of DMEM medium (1X104 cells/well) in a 96-well microtiter plate in triplicate and treated with the prodrug 5-FC (0- 25 mM) for 48 hours.
Results and Discussion
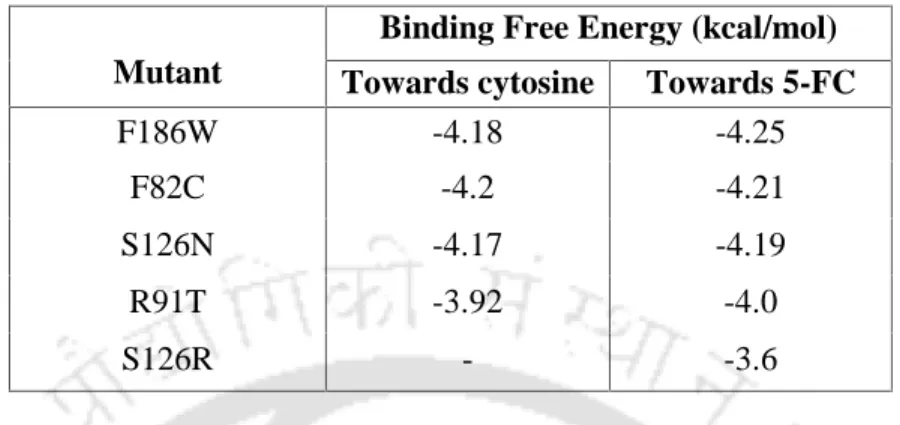
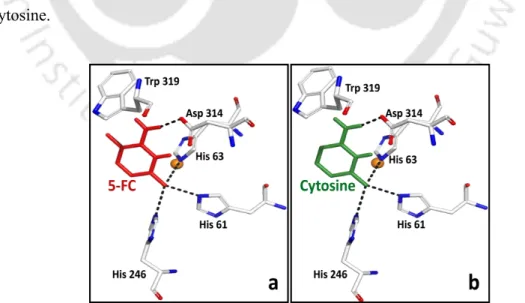
Fifty-two of the 68 mutants showed reduced docking scores against cytosine than WT (Appendix Table 3.A3). The absence of any significant changes in the active site residues in the superimposed images of the mutant and WT further supported this observation (Figures 3.5a, 3.5b). These essential hydrogen bonding interactions can facilitate the binding of ligands in the active site of the enzyme.
The catalytic efficiency (Kcat/Km); where Kcat = Vmax[E] ([E] is the total enzyme concentration) was also calculated for each of the WT and the mutant CD and is shown in Table 3.3. It is noteworthy that the docking studies also revealed the inability of cytosine to bind to the active site of the enzyme. The relative specificities of the WT and the mutants with both substrates are given in Table 3.5.

Conclusion
To assess the effect of WT CD and the S126R mutant, HeLa cells transfected with pVITRO2:CD and pVITRO2:S126R were subjected to in vitro 5-FC sensitivity assays as described above. The results of the prodrug sensitivity assay showed that the S126R variant increased sensitivity to 5-FC compared to WT CD. Compared with WT CD, the S126R mutant showed a remarkable response to 5-FC with reduced cell viability.
MTT cell viability assays for 5-FC sensitivity were performed for HeLa cells transfected with WT CD and S126R. The docking results were further confirmed experimentally by protein expression and enzyme assays and revealed the fact that the new S126R, F186W and F82C mutants may act as better candidates in SGT applications; with greater relative specificity for the 5-FC prodrug than WT CD. To further understand the effects of the developed mutants in a mammalian expression system, WT CD and its S126R variant were cloned and transfected into human cervical cancer cell lines.
Appendix
Binding energy scores and number of clusters obtained from AutoDock for WT and the mutants docked with cytosine and 5-FC. The electrostatics and van der waals energy scores obtained from AutoDock for the mutants docked with cytosine and 5-FC. With cytosine With 5-FC With cytosine With 5-FC. kcal/mol) Van der Waals energy (kcal/mol).
With cytosine With 5-FC With cytosine With 5-FC. kcal/mol) Van der Waals energy (kcal/mol). The chapter also describes the in silico site-directed mutagenesis studies performed with the UPRT protein.
MOLECULAR CLONING, FUNCTIONAL ASSAYS AND BIOINFORMATICS STUDIES ON E.COLI URACIL
Introduction
Outline of the Research Work
Experimental Section 1. Materials
Then, the sample was centrifuged at 15,000 rpm for 30 minutes at 4C, and the supernatant, which contains the soluble fraction of the UPRT protein, was collected. The concentration of the purified protein was determined using the Bradford method with BSA as a standard. A purified fraction of the protein of interest was run in SDS-PAGE; then blotting was performed in a PVDF membrane.
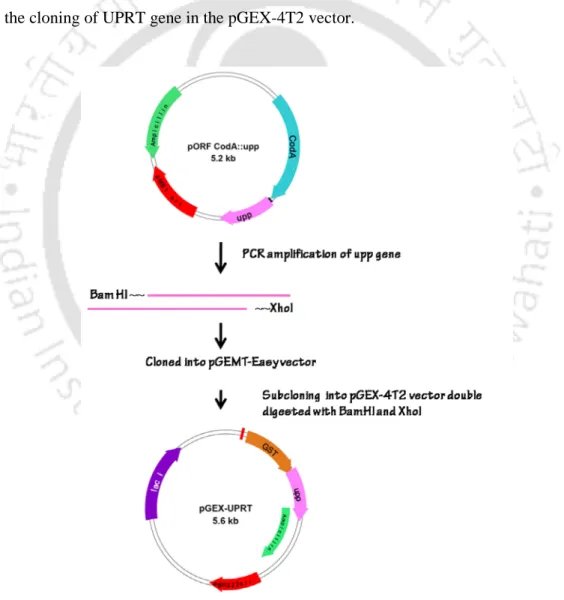
Uracil degradation and UMP formation were monitored at 265 nm and 280 nm, respectively. Mutagenesis hotspots present over the single unit of UPRT (208 amino acid residues) were identified by the HotSpot wizard. The web server takes the protein's PDB file (code: 2EHJ) as input and assigns variability scores from 1-9 to each and every residue present on the protein. The Lamarckian Genetic Algorithm (LGA) was applied to achieve different binding modes of the ligands which are energetically favorable.
Results and Discussion
Maximum root mean square tolerance is set as 2 Å. Figure 4.3. Purified fractions of the N-terminal GST-tagged UPRT and GST enzymes. Molecular docking of the mutants with uracil revealed that there were 19 mutants that did not allow the ligand to bind to their active site. As observed in the case of the uracil, lack of any significant changes in the active site residues after superposition of WT and the mutant proteins (Figures 4.6c-4.6e) could attribute the mutants G191K, I194D and K208V to have similar activity versus 5-FU as with the WT protein.
The hydrogen bond interactions achieved while mutant I194L was coupled to uracil and 5-FU are shown in Figures 4.7a and 4.7b. Figure 4.7a shows that the oxygen atom of the Met 132 is bonded to the N3 atom of the substrate uracil. The nitrogen atom of Val 139 forms a hydrogen bond with the O2 atom of uracil and 5-FU; Furthermore, the oxygen atom of the Met 132 forms a hydrogen bond with the N3 atom of both ligands; Figure 4.9a shows that coupling mutant M101V to 5-FU showed highly significant hydrogen bonding interactions.

Conclusion
Appendix
Mutant binding Free energy (kcal/mol) Binding affinity with Uracil with 5-FU With Uracil with 5-FU.
No Mutant Binding Free Energy (kcal/mol) Binding Affinity With Uracil With 5-FU With Uracil With 5-FU
TASSER predicted three dimensional structure of E. coli CD-UPRT fusion protein
CLONING, EXPRESSION AND PURIFICATION OF THE BIFUNCTIONAL CYTOSINE DEAMINASE-URACIL
- Introduction
- Outline of the Research Work
- Experimental Section 1. Materials
- Results and discussion
- Conclusion
The three-dimensional structure of the CD-UPRT recombinant fusion protein was modeled using the I-TASSER server (Zhang, 2008). The release of a 2 kb fragment of the CD-UPRT gene is clearly visible in the agarose gel in Figure 5.2. Bands corresponding to the overexpressed CD-UPRT fusion protein began to appear in the supernatant (S) with the addition of the sarkosyl detergent.
SDS PAGE showing the increase in the binding affinity of the recombinant CD-UPRT protein with the glutathione-agarose beads using different ratios of sarkosyl and Triton. Finally, three-dimensional structure of the CD-UPRT fusion protein was modeled and is shown in Figure 5.6. More than 95% of the recombinant protein was solubilized in a single step using the alkyl anionic detergent sarkosyl.
CONCLUDING REMARKS
Summary of the Present Work
The in silico results obtained may lead to the development of UPRT mutants to increase the sensitivity of the cancer cells more towards the drug 5-FU. Finally, the recombinant fusion protein CD-UPRT was cloned and overexpressed with N-terminal GST tag. The intact recombinant protein was effectively recovered from the inclusion bodies using sarkosyl as a solvent.
The optimization of the ratio between the detergents sarkosyl and Triton ×100 was performed to improve the binding affinity of the recombinant protein to the glutathione beads. Furthermore, enzyme assays of the recombinant protein against its substrates cytosine and 5-FC have been described, the kinetic parameters of both substrates. The efficient purification and kinetic values determined for the CD-UPRT enzyme may open new avenues to redesign hybrid CD-UPRT with high therapeutic efficacy in cancer gene therapy.
Scope for Future Work
Experimental gene therapy for brain tumors using adenovirus-mediated transfer of the cytosine deaminase gene and the uracil phosphoribosyltransferase gene with 5-fluorocytosine. In vivo gene therapy for colon cancer using adenovirus-mediated, cytosine deaminase and uracil phosphoribosyltransferase fusion gene transfer. Apoptotic induction with bifunctional E.coli Cytosine Deaminase-Uracil Phosphoribosyltransferase-mediated suicide gene therapy synergism by curcumin treatment in vitro.
Understanding apoptotic signaling pathways in cytosine deaminase-uracil phosphoribosyl transferase-mediated suicide gene therapy in vitro. Combined radiation and gene therapy for brain tumors with adenovirus-mediated transfer of cytosine deaminase and uracil phosphoribosyltransferase genes. Combined suicide gene therapy for human colon cancer cells using adenovirus-mediated transfer of Escherichia coli cytosine deaminase gene and Escherichia coli uracil phosphoribosyltransferase gene with 5-fluorocytosine.