In twentieth-century biology, the physiology of cells, tissues, organs, and organisms in all kingdoms was the main focus of biological research. These include sensitive self/non-self recognition between slightly differently adapted populations of the same species in the same ecological habitat.
Signaling Molecules Serve in More than One Communication Process
Interpretation and Response Behavior to Sensory Data of Inanimated Nature
Such defense activities are also deployed as antimicrobial substances against the microflora in the root zone. Intercellular communication in the root zone (in the soil) differs from that in the stem region above ground.
Intracellular Communication
The cellular organization of the roots is determined during the plant's embryonic development and is controlled by intercellular communication. Telomeric and subtelomeric sequences consist of tandem repeats at the physical end of the chromosomes.
Viral Symbiotic Interactions
If, for example, the message transmitted was 'the sky is blue', little information is being conveyed as the information is little surprising. If, on the other hand, the conveyed message is "the sky is green and black-striped", this is surely surprising, containing unexpected new information, thus increasing its novelty and in turn its information content. .
Noise Is Likely Inevitable in Living Systems
One of the main concerns of Shannon and Weaver (1949) was to try and assess the accuracy of message transmission over telephone lines.
Evidence for Noise in Genetic Circuitry
Noise Can Spontaneously Induce Polarity and Ensure Each Cell Is Effectively a Unique Phenotype
Transcription and Translation in Single Cells Takes Place in Brief Bursts
The learning mechanism involves the variable synthesis of specific proteins in this case, and such learning and memory capabilities are equally present in plant cells (Trewavas1999). A further study that labeled 2500 proteins in yeast under different growth conditions found that there were dramatic differences of specific proteins in the noise that were related to function (Newman et al.2006).
Is Noise Useful or If Not, Can It Be Reduced?
However, these authors also reported that there was much more noise in proteins that responded to environmental signals, while those involved in protein synthesis were much quieter. There are physical limits to the sensitivity with which external signals can be sensed, and low-impact signals will only be perceived with greater noise than larger ones (Bialek et al. 2005).
Relevance of Noise in Genetic and Transduction Circuitry for Plant Development
If the transcription factor is short-lived, the noise can be amplified by inducing extrinsic noise on each of the genes to which the transcription factor binds; including, if so arranged, the original transcription factor itself. Depending on the number of transcription sites and the potential variable strengths of promoters, different target genes can be tuned to switch to high output at different concentrations of the transcription factor.
Synchronising Effects of Signals in Plant Cells
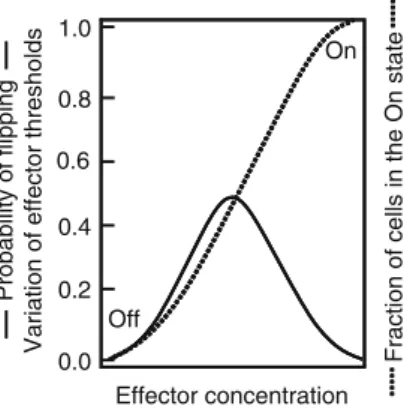
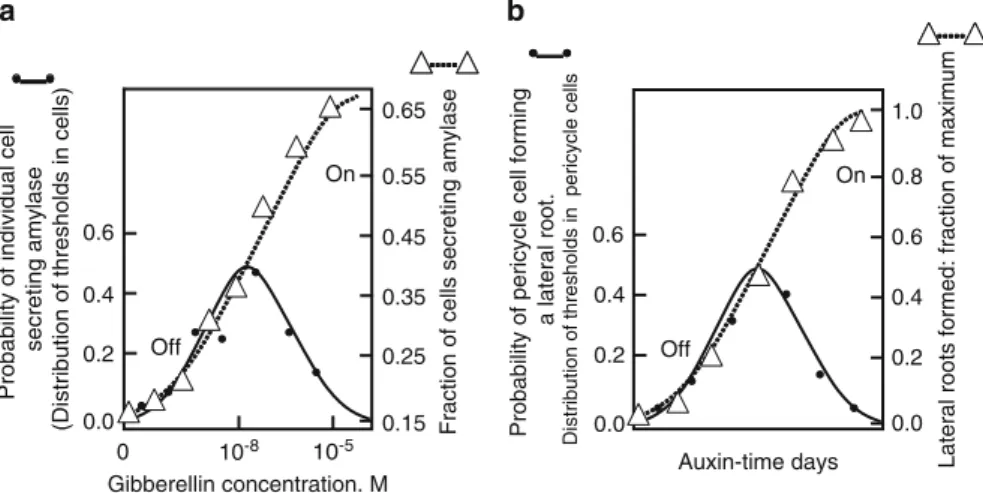
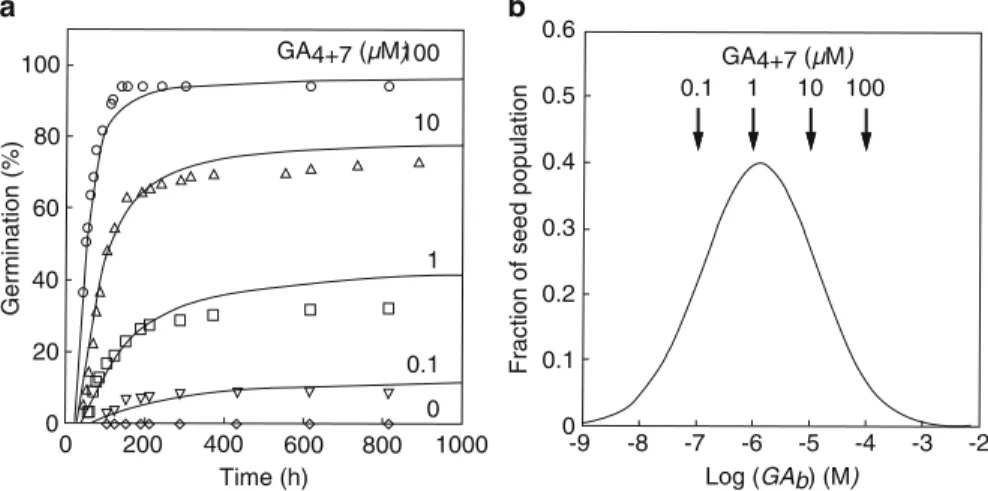
The distribution of thresholds and thus the probability of cells to synthesize amylase is similar to that of Figure 2 and . The symbols are plotted on the template of Fig.1 and indicate the strong similarity in behavior to Fig.1.
Stochastic Responses Are Observed in Individual Plant Cells In Situ
The distribution of thresholds is stochastic and likely results from stochastic variation in noise coupled with a positive feedback mechanism during aleurone cell development. As more cells cross their thresholds as auxin exposure increases, more lateral roots form.
Two Signals in Plant Development Change?
However, the speed with which patches change guard cell openings in response to closure signals suggests potential patch interaction arising from another faster source. Excess short-term water loss from the leaf causes short-term stomatal closure through vapor phase signals.
Cytoplasm Is an Organised, Highly Structured Network
Of note is the peripheral cytoskeleton complex located beneath it and attached to the plasma membrane, which is about 100 times thicker than the plasma membrane (Alberts et al. 1983). It is this structure, rather than the inner cortical matrix, that is known to be responsible for regulating specific aspects of morphological development in Acetabularia (Briere and Goodwin 1988; . Goodwin and Pateromichelakis 1979; Goodwin et al. 1983; Mandoli 1998).
The Design of Specific Synthetic Gels Is Intensely Researched
All these data suggest that the cytoplasm is a complex integrated network with perhaps microdomains specific for specific functions. The description of the cytoplasm as a gel capable of transitioning to a sol is long-standing and owes much to observations of organisms such as Amoeba whose pseudopodial behavior is constructed by rapid changes between gel and sol.
Synthetic Gels Indicate the Presence of Structured Water
4 Potential connection of structured water to gel formation in cells. a) Water dipole, slight negative charge on oxygen, slight positive charge on hydrogen. Cytoplasmic structured water (referred to as restricting the freedom of movement of water molecules) has been detected by various physical approaches such as NMR, frequency dielectric dispersion and quasielastic neutron scattering (Pollack 2001).
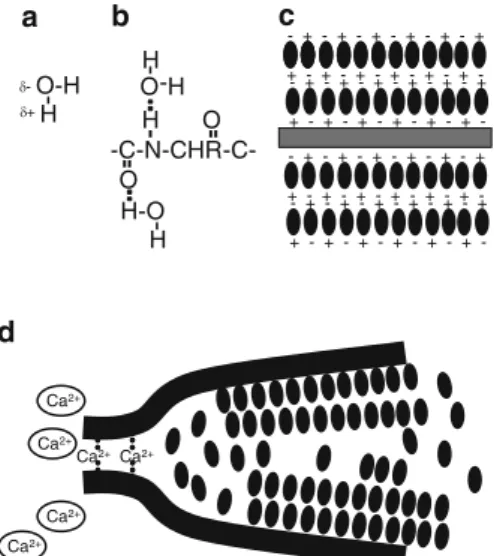
Structured Water in the Cytoplasm May Affect Ion Fluxes
The third, more weakly bound water originates from hydrogen bonding to the water molecules already attached to the open polypeptide regions and can form layers of structured water bonding, four to even ten layers deep as indicated in Fig.4c (Pollack2001; Ling2006). This structured water is in a form somewhere between the structure of ice and liquid, that is, in the structure expected of a gel.
The Impact of Structured Water for Cytoplasmic Functioning
Watterson (1987) hypothesized that other proteins (at least 60 are known) bind to actin by mimicking the topological structure of water around actin and removing structured water as a complete entity. Finally, Pollack (2001) suggested that structured water may play an important role in allowing water to rise easily to the tops of tall trees.
Gels and Phase Transition Cooperativity: conformational spread
The ordered or structured water that apparently permeates the entire gel prevents the formation of the normal ice structure, which itself depends on a tight arrangement of hydrogen bonding between water molecules. The binding of a single gelsolin molecule to one end propagates a conformational change throughout the entire actin molecule, and this may be sufficient to disrupt the structured water between adjacent molecules, thereby breaking the actin gel structure.
If the conformational change allows critical proteins to now contact each other, this may not be sufficient to initiate new developmental changes. Once initiated, the entire structured water complex is cooperatively destabilized using the energy available from the low-entropy structure of the structured water.
Is There a Role for K + in Phase Transition?
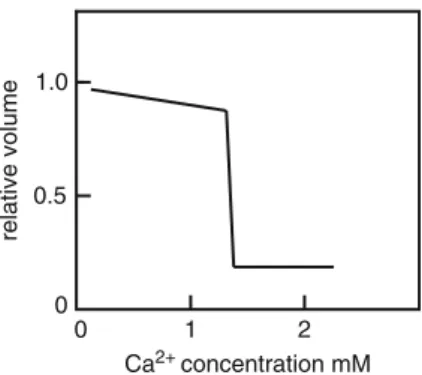
Urry (1971) has shown that Ca2+ binding in two proteins and doubtless many others occurs in regions rich in glycine residues increasing the potential binding sites available and cites sulfhydryl groups as potential binding sites as well. The gel structure must be in some kind of dynamic state that allows some Ca2+ ions to penetrate the gel structure to initiate cross-linking.
Could Phase Transitions Be Communicated Through The Plasmodesmata?
Goodwin BC, Patromichelakis S (1979) The role of electric fields, ions and cortex in the morphogenesis of Acetabularia. Despite differences in amounts and mixtures between plants, tissues and specific invaders, they participate in the regulation and translation of induced defense signaling (Pieterse et al. 2009).
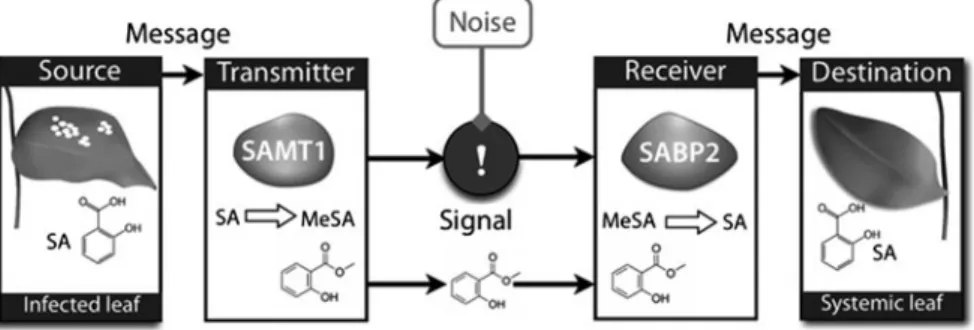
Salicylic Acid
Recent advances in the understanding of the role of phytohormones have revealed an extensive interplay between various hormones (Pieterse et al.2009). As a monomer, NPR1 translocates to the nucleus where it interacts with transcription factors such as TGAs and WRKYs to enhance defense gene expression (Vlot et al.2009).
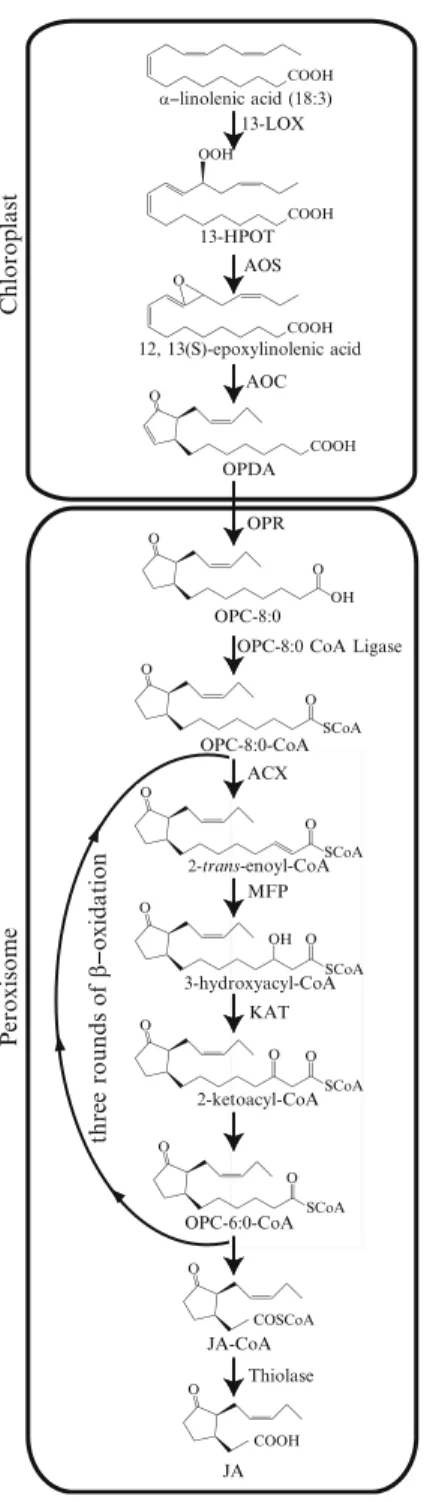
Jasmonic Acid
However, grafting experiments showed that SA is not required in the tissue that transmits the SAR signal, while it is necessary in the systemic tissue that receives the SAR signal (Vernooij et al.1994). In addition to modifying NPR1 by shifting the redox state, SA also induces the expression of thioredoxins (TRX) that catalyze the monomerization of NPR1 (Tada et al.2008).
Ethylene
JA can be further metabolized into various products, including volatile MeJA, and it can be conjugated with amino acids and sugars (Wasternack2007). Further studies must be undertaken to reveal in detail how the products of JA-responsive genes contribute to the fight against disease.
Auxin
Abscisic Acid
During the initial phase of invasion, ABA positively regulates resistance by mediating stomatal closure. In the subsequent early phase of invasion, ABA increases resistance to fungi and oomycetes by triggering callose deposition, but also decreases resistance by inhibiting reactive oxygen species (ROS) generation and callose accumulation in bacterial infection.
Brassinosteroids, Cytokinins, and Gibberellin
Consequently, GA appears to be involved in promoting resistance to biotrophs and susceptibility to necrotrophs.
Systemin
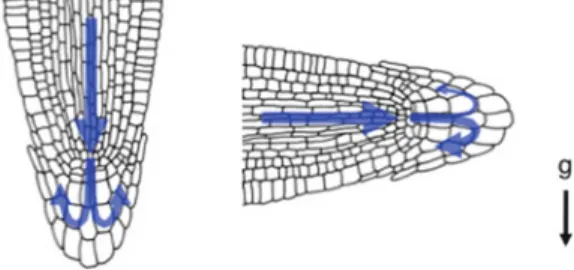
The expression of the JA-responsive genePDF1.2 requires the simultaneous activity of JA and ET signaling cascades (Penninckx et al. 1998). Second, amyloplast sedimentation is observed in the direction of gravity in the tissue of the reacting organ (Takahashi 1997).
Gravitropism
Methyl jasmonate (MeJA) was the first oxylipin identified, originally extracted from Jasminum grandiflorumoil (Demole et al.1962). Mutants defective in one of the two MFP genes (target 1) show impairment in wound-induced JA production (Delker et al. 2007).
Immunity Against Insect Herbivores
Depending on the type of invading insect, distinct, yet overlapping, gene expression patterns can be observed (De Moraes et al. 2001; Heidel and Baldwin 2004). Recent studies suggest that oral insect secretions, such as fatty acid–amino acid conjugates and peptides, can be perceived (Kessler and Baldwin2002; Schmelz et al.2006).
Pathogenic Microbial Defense
COMATOSE ATP-binding cassette transporters are required for full fertility in Arabidopsis.Plant Physiol. The SCF(COI1) ubiquitin ligase complexes are required for jasmonate response in Arabidopsis.Plant Cell.
Anatomy and Biochemistry of the Phloem
These methods can be useful for the identification of phloem sap components, although they potentially cause artefacts (Dinant et al. 2010). As for the isolation of phloem cells, laser microdissection has been used successfully (Nelson et al. 2006).
Long-Distance Signalling to Wounding and Herbivory
Hydrogen peroxide has also been suggested to form a secondary messenger in sink organs (Orozco-Cardenas et al.2001). This established that ROS accumulation along a systemic signaling front is essential for long-range signaling in plants (Miller et al.2009).
Systemic Response of Plants to Pathogen Attack
MeSA is then transported systemically and constitutes a critical signal for establishing the systemic response (Seskar et al.1998). Jasmonates have also been described as essential signals for establishing systemic immunity in response to Pseudomonas syringae (Truman et al. 2007).
Phloem Conductivity in Response to Injury and Aphid Feeding
In addition, airborne cues, including MeSA, MeJA and green leaf volatiles, contribute directly to these defense mechanisms (Farmer2001; Frost et al.2007; Shah2009). These changes were specifically regulated by aphid attack, as viral or bacterial infections led to a different response (Divol et al. 2005).
Response to Phosphate Starvation
Moreover, this miRNA was detected in the phloem sap of two plant species (Pant et al. 2008). The existence of additional, miR399-independent systemic signals triggered by vacuolar Ca2+/H+ transporters has recently been pointed out (Liu et al. 2011b).
Nitrate Homeostasis
In roots, this transduction pathway also interacts with auxin signaling that also affects nitrate nutrition (Krouk et al. 2011). Interestingly, two of these miRNAs, miR167 and miR169, were detected in the phloem sap of pumpkin or rapeseed, suggesting a role in phloem long-distance signaling (Yoo et al.2004; Buhtz et al.2008,2010).
Plant Reproduction: The Identification of a Florigen Component
Furthermore, the results of Lifschitz et al. 2006) suggested that SFT-stimulated signals are conserved in different plants. In this work, two FT-like proteins were detected in the phloem sap of Cucurbita maxima (Lin et al. 2007).
Other Components Acting on Flower and Tuber Induction
Furthermore, when labeled GA5 is exogenously applied, it is transported to the shoot apex (King et al. 2001; King and Evans 2003). These increases are likely due to the transport of GA4 and sucrose produced outside the shoot apex (Eriksson et al. 2006).
Vegetative Development and Morphogenesis
Another major exciting issue is the identification of the factors necessary for the transport of macromolecules in the course of translocation. The recent discovery of the formation of large ribonucleoprotein complexes in the phloem sap indeed suggests that this process is highly regulated and may be involved in the specific transport of selected molecules (Ham et al. 2009; Ma et al. 2010; Li et al. al. 2011).
Intercellular Protein Trafficking
Antirrhinum DEFICIENS (DEF) and GLOBOSA (GLO) B factors move from L2 to L1 in a developmentally regulated manner (Perbal et al. 1996). It was proposed that this transport likely works in both directions (Sieburth et al.1998; Urbanus et al.2010).
Movement of Small RNAs in Floral Development
Genetic suppression of L-rhamnose biosynthesis results in transcriptional upregulation of cell wall remodeling genes (Diet et al. 2006). This process is facilitated by cell wall enzymes produced in the subventral glands and secreted by the nematode (Davis et al. 2008).
Does the Plant Sense the Nematode While It Migrates Within the Root Tissue?
Unfortunately, there is no information on the involvement of any plant receptors that recognize the extracellular cell wall degradation products in the nematode perception of the nematode during the migration phase. Possibly the L-type lectin RKL and CrRLK-L1 may be involved in the recognition of the cell wall damage caused by endoglucanases secreted by nematodes (cellodextrin).
Nematode Effectors Interfere with Cell Wall Performance in the Nematode Feeding Site
However, during later stages of feeding site development, most of these receptor-like kinases do not show any change in their expression or are down-regulated (Szakasits et al. 2009). However, so far, no experimental data regarding the early stages of the nematode-plant interaction support that, but in older syncytia, most WAKs are down-regulated or their expression remains unchanged ( Szakasits et al. 2009).
Expression of Cell Wall Enzymes in the Nematode Feeding Site Is Specifically Affected
During nematode infection, these different triggers affect a large number of different sets of cell wall-related genes, including genes involved in cell wall biosynthesis, degradation, and remodeling. In tomato, it has been shown that EXPA5 may be involved in cell wall relaxation that supports feeding site hypertrophy.
Overproduction of Roots in Response to Another Plant’s Roots
This response has been referred to as a 'Tragedy of the Commons' scenario (Maina et al. 2002;. The source of the applied activated carbon may also be important in determining the response achieved (Kabouw et al. 2010).
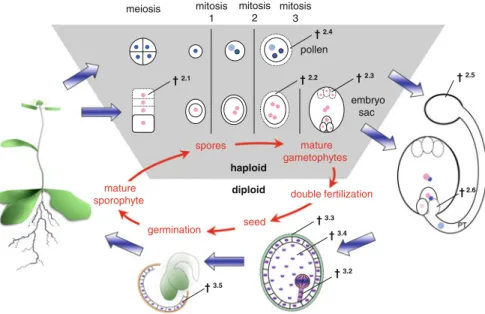
Nucellar Cell Death
After meiosis, the callose deposits surround all four megaspores but are removed asymmetrically in the cell wall of the FM facing the parent sporophyte tissue. It has been speculated that the callose blocks PCD inhibitory signals thus only reaching the surviving functional megaspore (Papini et al. 2011).
Antipodal Cell Death During Embryo Sac Maturation
This is thought to sustain pollen growth through vesicular transport of degraded material that is taken up by pollen by endocytosis (Hiratsuka and Terasaka 2011).
Cell Death in the Tapetum Layer
Pollen Cell Death During Incompatible Pollen-Pistil Interactions
Double fertilization results in two individual fertilization products: the diploid zygote that will develop into the embryo and the triploid nutritive tissue of the endosperm. The fate of the two fertilization products cannot be more different—while the embryo lives on to form the next sporophytic plant generation, the endosperm's function and life is limited to seed development and ends with germination (Berger 2003; Berger et al. 2006; Sabelli and Larkins 2009).
Synergid Cell Death During Fertilization
In Arabidopsisgfa2 mutants, the micropyle correctly attracts pollen, but pollen penetration and synergistic cell death do not occur (Christensen et al. 2002). Live-cell imaging of the fertilization process in Arabidopsis has shown that cell death of synergid cells is initiated only after pollen tube–synergid cell contact, but before pollen tube penetration and release.
Embryonic Suspensor Cell Death
These data suggest that, at least in Arabidopsis, short-range communication rather than mere physical disruption induces synergid cell death (Sandaklie-Nikolova et al.2007). A Tudor staphylococcal nuclease (TSN) was found to be a natural substrate of type II metacaspase in spruce (Sundstrom et al.2009).
Cell Death During Seed Coat Formation
In addition to their role in HR cell death, one of four Arabidopsis VPEs, dVPE, has been shown to control developmental cell death in the two inner integument layers during seed development. dVPE is specifically expressed in these two layers before the initiation of cell death, and mutant plants deficient in dVPE show a delayed cell death and collapse of these inner integument cells (Nakaune et al.2005).
Central Endosperm Cell Death
Ingram (2010) speculates that the cell death regulation in ESR and starchy endosperm must be quite different as they serve different purposes and produce different outcomes, autolysis in the ESR versus maintenance of the cellular corpses in the starchy endosperm. However, concrete evidence exists for the participation of phytohormone signaling in cereal endosperm cell death.
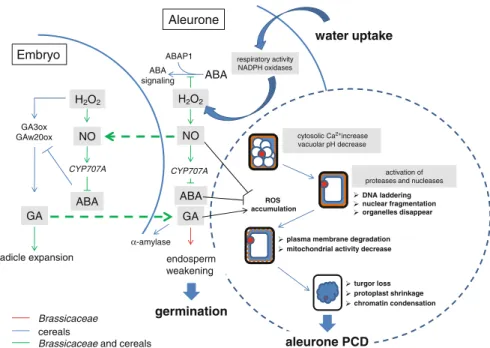
Survival and Cell Death Decisions During Aleurone Development
Bethke PC, Lonsdale JE, Fath A, Jones RL (1999) Hormonally regulated programmed cell death in barley aleurone cells. Young TE, Gallie DR (2000b) Regulation of programmed cell death in maize endosperm by abscisic acid.