We applied new time-implicit dynamic models to re-evaluate the classic Luria-Delbrück Fluctuation Test (LDFT), which has established the spontaneous and random nature of genetic mutations. Despite the study's profound impact, full quantitative features of the apparently simple mutational population dynamics are still under active investigation. In particular, with a recent increase in interest in Bayesian inference, there has been renewed interest in the Luria-Delbrück experiment in terms of potential support for the controversial concept of "adaptive mutation", whereby cells increase mutation rates corresponding to environmental stresses.
Although the majority of the detailed analyzes have shown that the mutations are not intended in any adaptive way but occur without any consideration of their potential benefits, there is a possibility that random spontaneous mutation works together with Lamarckian mechanisms as exemplified by the CRISPR -Cas immunity in bacteria. Here, we used an alternative mathematical analysis, where the size of the wild-type population takes the place of time. First, by eliminating time, the common but unjustified assumption of exponential growth could easily be relaxed.
Furthermore, our analysis re-discovers all stochastic versions of the original Luria-Delbrück model and the resulting statistics can be re-examined on a more quantitative basis, shedding light on recent calls for expanding the Modern Synthesis.
Random versus adaptive mutations
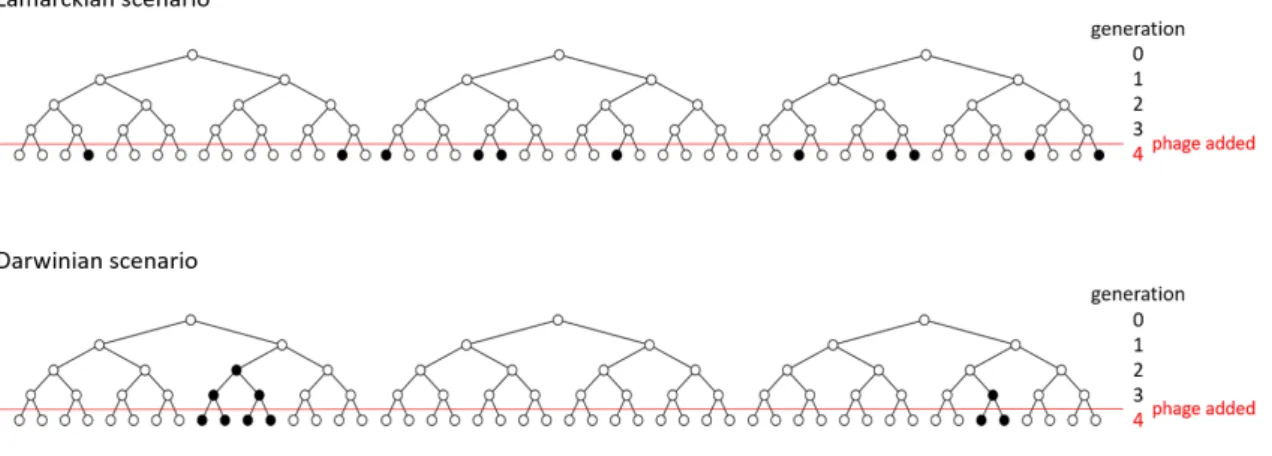
In a famous experiment in 1943, a small number of Escherichia coli (less than 500) were inoculated into individual culture tubes. By repeating the experiment several times, the mean number of mutants and the variance can be estimated. In contrast, according to the Darwinian scenario (random mutation hypothesis), a mutation can occur before the introduction of a bacteriophage and spread their resistance to their descendants. C) The variance of the number of resistant colonies is shown as a function of the mean number of resistant colonies.
The results show that the observed fluctuations are larger than those predicted by the Lamarckian scenario (adaptive mutation model), which predicts that the variance should be equal to the mean. In the Lamarck scenario, where resistance to the virus is caused by induced activation in bacteria, mutations will only occur after phage infection, and each plate should contain approximately the same number of phage-resistant bacteria. In other words, the distribution of the resistant (mutant) cells in each culture should be Poisson, and therefore the variance in the number of resistant bacteria is more or less the same as the mean.
On the other hand, in the Darwinian scenario, where resistance is due to heritable genetic components in bacteria, mutations will occur randomly before phage exposure and there should be a large variation in the number of phage-resistant bacteria. From the results, they discovered that the distribution of phage-resistant bacteria was rather heavy-tailed, thus not conforming to the Poisson distribution, leading to the conclusion that bacteria evolve by the Darwinian mechanism. The black dots illustrate phage-resistant bacteria and the white dots are bacteria susceptible to the T1 phage.
A single bacterium grows for a few generations and the T1 phage is then introduced into the plate culture (the fourth generation of each tree in the picture). This will result in a standard deviation of the number of resistant bacteria, which is measured by the square root of their mean. In other words, there will be little variation in the number of resistant bacteria between trees.
Therefore, a number of experiments will reveal a broad distribution of the number of resistant bacteria, meaning that the variance will be large compared to the Lamarckian case.
![Fig. 1.1 Schematic of the Luria-Delbrück experiment excerpt from [6]. (A) Experiment starts with small number of bacteria (less than 500) grown for approximately 15 generations](https://thumb-ap.123doks.com/thumbv2/123dokinfo/10523025.0/12.892.142.777.445.815/schematic-delbrück-experiment-excerpt-experiment-bacteria-approximately-generations.webp)
Review of the previous studies
Luria-Delbrück distribution and beyond
The Luria-Delbrück model
We begin by listing the fundamental assumptions of the Luria-Delbrück formulation for future reference, as the subsequent formulations presented below are essentially variants of the Luria-Delbrück model. At a constant rate β1, normal bacteria grow deterministically and the number of normal bacteria at time is. Thus, if a mutant is initiated by normal bacteria at time s > 0, the clone of that mutant will be of size eβ2(t−s)fort≥s.
Denoting X(t) as the number of mutant bacteria that exist at a given time, and from Assumption 1–4, X(t) can be expressed mathematically as .
The Holmes model
Mutation occurs only during synchronous divisions, where each of the offspring independently turns into a mutant (Darwinian), or only after introduction of bacteriophage (Lamarckian). If we denote N0 as the initial number of wild-type bacteria sensitive to phage and gas, then the total number of epochs before the introduction of bacteriophages is the total number of bacteria N after the final level of division. Now that we have θL for the probability of a Lamarckian mutation after phage introduction and reduce the probability of an additional mutation occurring in the post-resistant progeny, the average number of Lamarckian mutations in epoch g is the same.
For the Darwinian model, there may be a number of ways to estimate a certain number of mutant bacteria k in the population of size N before bacteriophage introduction. As an example, to obtain five resistant bacteria (k=5): (i) One mutation can occur two generations before the bacteriophage introduction (with the total living population N/4 at that generation) or (ii) Two mutations can occur one generation before introduction of T1 subjects, and a further mutation in the last generation.
Implicit-time formulation
Instead, a mathematical formulation of population dynamics of asexual populations came about, where the population size of the wild-type cells is the independent variable, playing the role of time [16]. If we use the size of the wild-type population (WTP) as the proxy of time, we do not need to assume any specific growth dynamics such as Monod, Gompertz, Richards or Stannard [21]. When a wild-type cell divides, regardless of the length of the cell cycle, there is a small probabilityµ that a mutant daughter cell will appear.
Assuming "no death" in a culture of wild-type bacteria growing from the initial size N0 to size N, we call the random variable describing the number of newly born mutant cells while WTP remains at n. In the case of the deterministic growth of a cell, probability only comes into play for the point in time at which a cell division gives rise to a mutant cell. Thus, letΓNn be the contribution of a mutant cell to the final number of the mutant cellsMN when WTP size reaches N,.
Once PGF is calculated with respect to tos, the inverse transformation gives the probability distribution for the number of mutant cells via . The contribution of the emerging mutant to the magnitude of WTP in the final population N was denoted as ΓNnXwhere the deterministic propagator ΓNn was assumed to be N/n for the deterministic case and the MGF eΓNnXn was expressed as⟨esΓNnXn⟩=1−µ+µesN/n. The first term on the right-hand side of 2.30 is what we have already derived for the case of deterministic growth.
The following expression is the contribution of the stochasticity ofΓNn on the variance of the number of mutants. According to the combined model, the probability distribution for the number of mutant cells is given by the convolution of the two independent contributions, i.e. where λ is the mean to be determined by regression. The probability of the number of mutant bacteria is estimated by the convolution of the two independent contributions, i.e. The Poisson distribution from the Lamarck mechanism and that of the Darwin mechanism given by Eq. it is sufficient to calculate the PGF with different requirements.
Recently, Kessler and Levine [22, 23] found an explicit expression of the mutant population distribution in terms of the Landau distribution in both deterministic and stochastic growth models.
Moment generating functions
A single mutant that appeared when the WTP size is larger expands to the size KnN when the wild-type population reaches N. The main chemical equation for the probabilities of the propagator Pr(KnN =m) is then given by the main equation. Now we change the notation to denote N as the wild-type population size of the mutant occurrence, and N as the final size of the wild-type population we obtain.
If the relative growth rate of the mutant is given by c̸=1, the transition probability for the mutant once it appears is. This equation can be rewritten as Eq. 3.8) with a simple scaling N→Nc for the ratio of growth rates of wild-type and mutant cells. FGM is therefore given by. 3.12) from which the CGF of the number of mutants can be derived.
The Luria-Delbrück experiment confirmed the validity of the conceptual framework of the modern synthesis. While LDFT provides clear evidence for random mutations before exposure, it does not rule out the possibility that mutation occurs in response to exposure to a selective environment. Moreover, the ability of the conceptual scheme of the modern synthesis to keep pace with rapid changes in developmental biology, genomics, and ecology has been constantly questioned.
Developmental processes, operating through niche construction, pervasive inheritance, and developmental biases, share responsibility for the rate and direction of evolution and the origin of character variation. Stress-induced mutations are an adaptive response to stress, but there is still no convincing evidence that the mutations are specifically targeted to a set of genes that are responsible for growth in a negative selective media. To qualify as "Lamarckian," mutations must be specifically directed to genes that can relieve stress.
In this thesis we investigated the possibility of including induced mutagenesis as a new component of the evolutionary synthesis. A population birth-and-mutation process i: Explicit distributions for the number of mutants in an ancient culture of bacteria. Scaling solution in the large population limit of the generalized asymmetric stochastic luria–delbrück evolution process.