These results indicate a critical role of the additional methyl group in determining the attractive potential between dsDNA. Our findings suggest plausible explanations for the physical origins of chromosome arrangement and its epigenetic modulation. In addition to protein-mediated programmed chromatin structuring, the direct electrostatic interaction between dsDNA can be a key factor when it changes with DNA sequence or chemical modifications such as cytosine methylation.
From MD simulations, we found that this is due to the steric repulsion of the polyamine chains out of the DNA grooves by the hydrophobic methyl groups that place them at the bridging sites between the dsDNAs.
Chromosome organization
Histone modification
The comparison of different histone functions and observation of their modifications would provide useful information about epigenetic regulation on chromosome [12, 25], Because histone forms the beads and strand formation with DNA, DNA flexibility and condensation on histone is considered to be an important role in the organization. chromosome.[26] Based on these reasons, this research used hexa-lysine (6lysine) as analog of histone and aimed to uncover the physical mechanism behind epigenetic regulation from interaction between positively charged lysine residues and DNA. By using modified lysine, such as tri-methylated lysine, we intended to detect the model histone modification effect on the DNA condensation compared to unmodified model histone 6lysine.
DNA methylation
FRET
FRET efficiency from donor emission
If the absorbance of the donor at the excitation wavelength is the same between two samples in properties such as excitation wavelength and concentration, the two emission intensities by wavelength are used to calculate the FRET intensity; E can be calculated as (equation 1.2). Where IDA is the total fluorescence intensity of the donor in the presence and ID is the total fluorescence of the donor in the absence of the acceptor. This calculation provides an absolute value relative to the quantum yield difference between donor and acceptor.
Where AD and ADA are the donor absorbance at the excitation wavelength in the donor and donor including acceptor samples, respectively.
FRET efficiency from donor/acceptor intensity ratio
Single molecule FRET
Multi-Color FRET
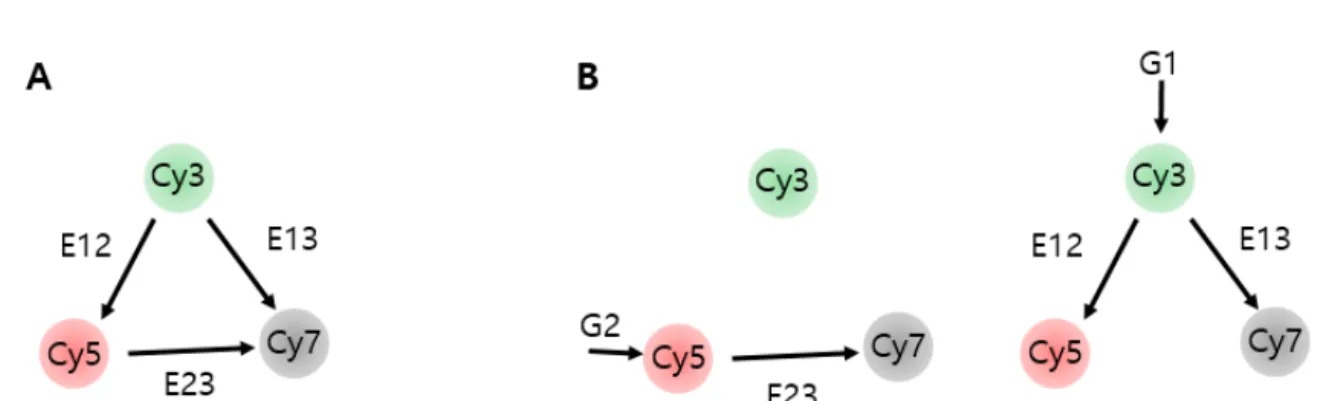
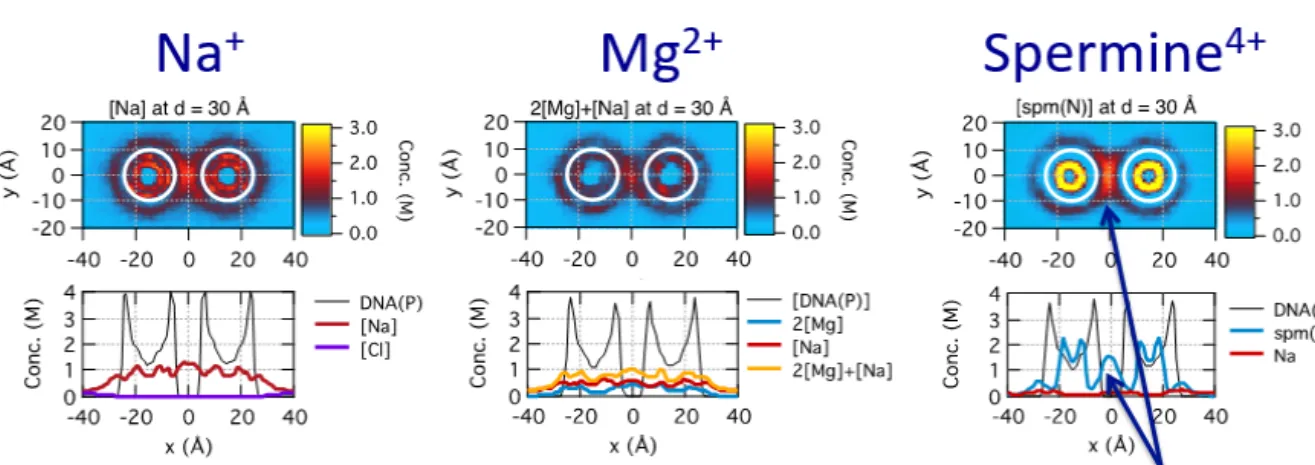
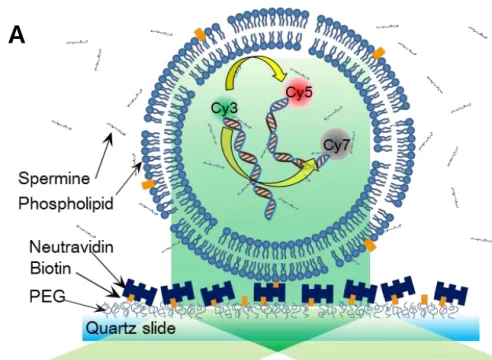
Property of Cy3-Cy5-Cy7 trio (A) Solid and dashed lines representatively represent the emission and absorption spectra of Cy3 (green), Cy5 (red), and Cy7 (gray). A schematic diagram for determining 3-color FRET efficiency using the single-molecule ALEX technique. a) FRET interactions between Cy3-Cy5 or Cy3-Cy7 or Cy5-Cy7 of each energy transfer (b) The ALEX technique allows three FRET efficiencies to be determined simultaneously[38].
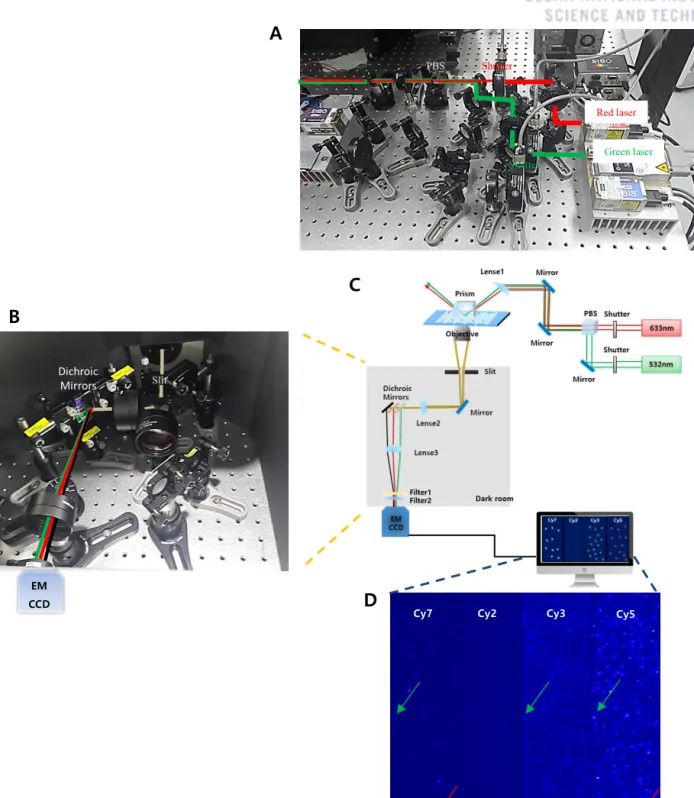
Experimental Setup of TIRF microscopy
Materials and Sample Design
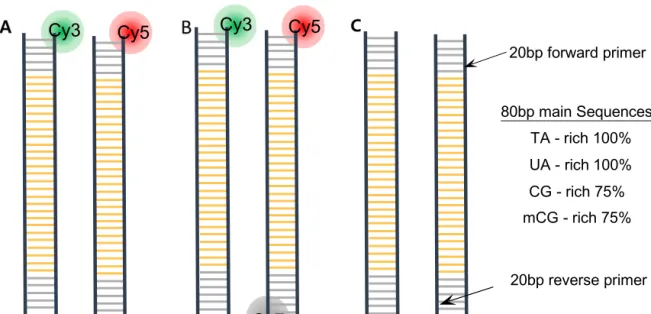
DNA Design
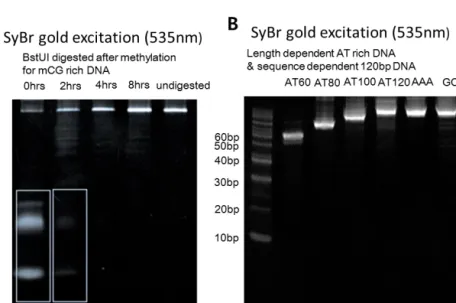
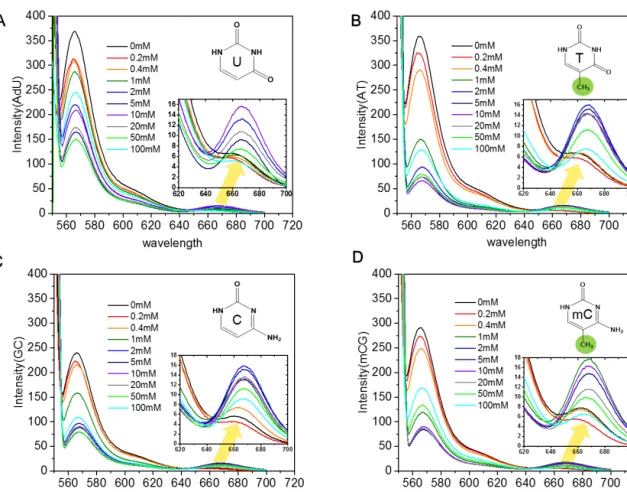
TA-rich and mCG-rich DNA have C5 methyl group on Thyimine and methylated Cytosine, respectively, while CG-rich and UA-rich DNA have no methyl group on C5 of Cytosine and Uracil. 120 bp methyl group-dependent DNA oligos were synthesized by Integrated DNA Technologies (US) with HPLC purification. Methyl group-dependent dsDNA constructs are produced by PCR using 120 bp designed templates and forward and reverse primers.
Unlike 120bp of TA, CG, mCG-rich DNA was made by PCR, UA-rich DNA was custom synthesized by exchanging uracil instead of thymine in only head sequence of TA-rich DNA. From this, sense and antisense template of UA DNA is prepared, finished double-stranded UA-rich DNAs were obtained by sense and antisense template at a molar ratio of 1:1.2 in the annealing buffer (10mM tris-HCl, 1mM EDTA , pH8.0, 50mM NaCl) for final concentration 2uM. The mCG-rich DNA constructs were obtained by 4 h methylation reaction using the CpG methyltransferase M.
To present different views on the effect of the methyl group on dsDNA attraction, we observed whether the methyl group pattern affects the attraction between dsDNA. We designed 100% TA with multiple thymine sequences (TTT), 25% TA (CG-rich), 25% TA with multiple cytosine sequences (CCC). For this reason, we prepared a customized sense and antisense CCC sequence template and obtained dsDNA by annealing as UA-rich dsDNA. In addition, we designed complementary DNA rich in 60 bp, 80 bp, 100 bp TA by reducing the number of thymine in the main sequence to measure the length-dependent attraction of dsDNA.

Polyamine
Ensemble FRET Measurement
Spectrofluorometer
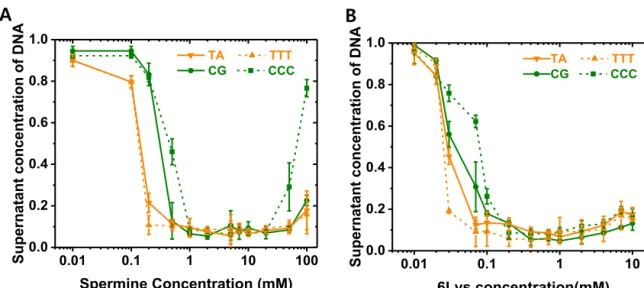
Based on this assumption, we titrated the concentration of the DNA molecules in the supernatant after precipitating them by varying concentration of the polyamine molecules. The concentration of DNA in the supernatant was determined by measuring the absorbance at 260 nm. These results suggest that the content of C5 methyl group, rather than the sequence composition of the DNA, determines the condensation force of linear polyamine molecules.
As expected, we observed enhanced contrast between them (Figure 3.3), consistent with previous computational prediction.[12] Thus, the effect of methyl content is maximized when we have consecutive methyl groups on the same DNA chain, likely due to the linear structure of polyamine molecules. Shorter DNA molecules saw a dramatic reduction in condensation power in the polyamine solution. (Figure 3.4.). A) Normalized concentration of DNA supernatant using length-dependent samples after measurement of DNA precipitation in different concentration range of spermine (B) in different concentration range of 6 lysine. In the Scatter plot of the average FRET from binding and bending events, we could predict when many binding events occurred in a simultaneous trace, bending events were rare and vice versa.
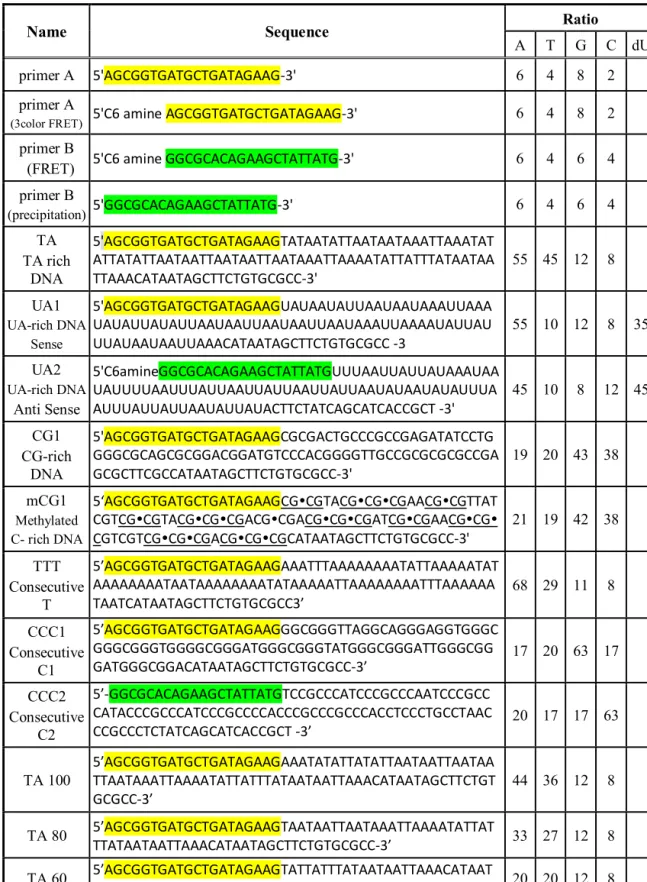
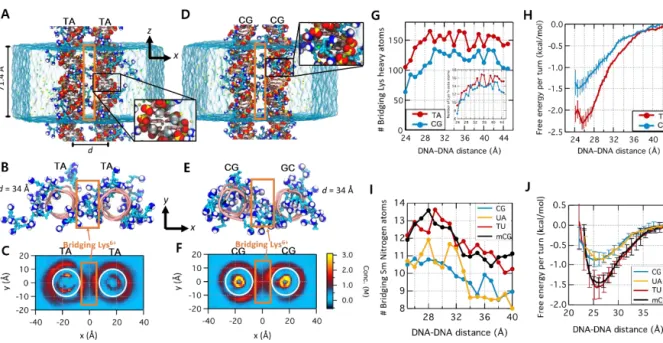
The methyl group in TA- and mCG-rich DNAs pushes the bulky polyamine molecules out of the DNA grooves, resulting in a lower polyamine density within the boundary of the double helix (Figure 4.1 C, F). To neutralize the phosphate charges, the emitted polyamines are located at the boundary of the DNA (Figure 4.1 C, F, G, I), leading to a stronger attraction between dsDNAs. These MD simulation results allow us to verify that steric repulsion of the polyamine cation molecules by the C5 methyl group increases the dsDNA affinity.
Data Analysis
DNA precipitation
The conditions for double-stranded DNA precipitation by the polyamines spermidine and spermine have been determined experimentally and compared with theoretical predictions. We performed a DNA precipitation experiment to further validate our results. In our experiment, we assumed that there is a difference in the supernatant concentration of DNA due to the strength of DNA condensation, determined by methylation. In this experiment, we prepared 500 ml tubes usually containing equimolar DNA and precipitation buffer (Tris HCl pH 8 20 mM, EDTA 2 mM, NaCl 50 mM) mixed with each polyamine (spermine/6lysine) at different concentrations from 0 mM to 100mM. (Figure 2.8) Then, after 5 minutes of incubation at room temperature, centrifuge the entire concentration range of the tubes for 5 minutes at 14,000 rpm, and measure the supernatant concentration of each DNA sample using absorption spectroscopy.
Single molecule FRET measurement
Schematic design of multi-color FRET measurement
By switching between green and red laser every hundred milliseconds, we can measure (A) state and (B) state almost at the same time. In the green excitation mode of 532nm, we can observe the parallel binding events and antiparallel binding events.
Analysis of single molecule traces
Ensemble FRET
DNA precipitation
In the red box in Table 2 we can check that parallel binding events are almost 2 to 3 times more common than anti-parallel binding events in the same condition, consistent with previous research[12]. Furthermore, we were able to establish that there is an anticorrelation between bending and binding events, by comparing the ratio between the binding fraction, the bending fraction to the level of the TA-rich constructs and their concurrent events (Figure 3.5). As we expected, the cross-correlation of the two FRET signals showed a negative correlation for ~9 sec, when averaged over 100 traces of single molecules selected in an unbiased manner (Figure 3.7 A). Furthermore, the scatter plot of the average FRET levels across the individual molecules also revealed the negatively correlated tendency of the two dynamics (Figure).
We quantified two parameters: the density of the charged elements of polyamines (Lys6+: lysine heavy atom, spermine: nitrogen atom) at the bridge region and the attractive potential energy curve between dsDNAs. The potential energy curve was measured by the umbrella sampling method by fitting the ensemble distance population to the Boltzmann distribution. Bridging Lys6+ molecules are Lys6+ inside the orange boxes shown in panels A–F. H) Interaction free energy of the two DNA helices as a function of d for two different sequences.
Recent super-resolution imaging showed that hypermethylated chromosomes result in silencing of the hypermethylated genes. [20] Chromosomal domains were shown to segregate into two compartments depending on their TA content. Yoo, J., et al., Direct evidence for sequence-dependent attraction between double-stranded DNA controlled by methylation. Ha, T., et al., Investigating the interaction between two separate molecules: fluorescence resonance energy transfer between a single donor and a single acceptor.
Hohng, S., et al., Maximizing the information content of single-molecule FRET experiments: multicolor FRET and FRET combined with force or torque. FAIRLAMB, Levels of polyamines, glutathione and glutathione-spermidine conjugates during growth of the insect trypanosomatid Crithidia fasciculata.
Multi-Color single molecule FRET
Discussion
The number of bridging polyamines was greater for the TA-rich DNAs than CG-rich DNAs at all distances (Figure 4.1 G, I). Such methylation-dependent polyamine density can be explained by the steric repulsion by the C5 methyl group. Due to this difference in the charge distribution in the system, TA- or mCG-rich DNAs exhibit greater attractive potential (Figure 4.1 H, J).
However, from our study on the effect of methyl group on DNA condensation, we can assume that not only DNA methylation but also polyamine methylation affects the DNA condensation force from steric repulsion of methyl groups against each other. From this hypothesis of steric repulsion synergy among methyl groups, we performed DNA precipitation to measure condensation force between dsDNAs molecules using tri-methylated 6lysine and our methyl group-dependent DNA samples. In the first measurement, we could see the possibility of methyl group synergy in DNA condensation.
Both TA- and CG-rich DNA with tri-methylated 6lysine settled faster at the bottom of the tube at lower concentration than with only 6lysine (Figure 4.2 B,C). In addition, we also observed the same trend for methyl group-dependent dsDNA condensation force: TA-rich DNA settled to the bottom faster than CG-rich DNAs at lower concentration of both 6K and 6KA. When binding event occurred, significantly reduced bending events were observed (Figure 4.4 A), we could calculate obvious negative cross-correlation graph using only 10 traces.
Conclusion
Rau, A comparison of DNA compaction by arginine and lysine peptides: a physical basis for arginine-rich protamines. Deng, H., et al., Structural basis of polyamine-DNA recognition: spermidine and spermine interactions with genomic B-DNAs of different GC content probed by Raman spectroscopy. Puglisi, An Oxygen Scavenging System for Improving Dye Stability in Single-Molecule Fluorescence Experiments.
Hohng, Single-molecule three-color FRET with both negligible spectral overlap and long observation time.
![Figure 1.1. A graph of FRET efficiency following distance(R) between a donor and acceptor fluorophore molecules [33]](https://thumb-ap.123doks.com/thumbv2/123dokinfo/10529284.0/16.892.276.631.711.1002/figure-graph-efficiency-following-distance-acceptor-fluorophore-molecules.webp)