Significant differences in the growth parameters were observed in both generation 1 and 2 plants when plants were grown under enriched and ambient CO2 conditions. We hypothesize that the Rhodes grass parent plants grow below ambient temperature and enriched CO2.
Introduction
Climate change, the causes and impacts
2) Environmentalists believe that both temperature rise and climate change are interrelated and ultimately occur continuously, but in a natural manner of cyclical events without any human involvement. 3) Scientists and researchers consider that global warming and climate change are happening inevitably due to human activities and negatively revolutionizing the earth's atmosphere (Goldberg et al., 2019; Mehnert, 2016).
Greenhouse gases
- Impacts of greenhouse gases on the earth-atmosphere system
- Carbon dioxide
Over a wide range of time, greenhouse gases have diversified significantly and lead to substantial changes in Earth's history (Jeffry et al., 2021). According to the Intergovernmental Panel on Climate Change (IPCC), radiative forcing is responsible for the influence of greenhouse gas or other climate factor on the radiative energy that affects the earth's surface (Kweku et al., 2017).
CO 2 enrichment and plant growth
- Effect of CO 2 enrichment on stomatal conductance, size and density
- CO 2 enrichment and photosynthetic rates
- Effect of CO 2 enrichment on leaf surface area and chlorophyll content
- Effect of CO 2 enrichment on plant morphological parameters
C4 plants have advantages over other plants in surviving in high CO2 habitats (Rudov et al., 2020). No significant change in total chlorophyll content was observed in alfalfa plants with increasing CO2 (Ksiksi et al., 2018).
C3 vs C4 responses to CO 2 enrichment
Transpiration is correlated with the evaporative cooling effect, so over time the evaporative cooling effect or water loss decreases (Ksiksi et al., 2018). Another study by Navie et al. 2005) stated that C3 weeds have a competitive advantage over C4 grasses at higher atmospheric CO2 levels (from 360 (ambient) to 480 (enriched) ppmv).
Maternal effects on plants
- Classifications of maternal effects
Likewise, these maternal effects can impair the fitness of the offspring by transferring few inappropriate traits, and this can lead to the evolutionary imbalance (Johnsen et al., 2005). This effect involves maternal parental adaptation to adapt the offspring phenotypically to the local conditions to achieve maximum offspring fitness (Marshall et al., 2008; Itonaga 2011).
Asexual propagation in plants
- Asexual propagation by cutting plant part
- Asexual propagation using maternal branch
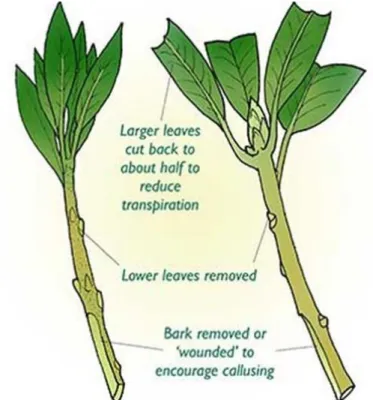
In contrast, hardwood or a mature branch of the tree is used for propagation in woody deciduous plants (Dhillon et al., 2011; Islam et al., 2011). The cut is made at the base, just below the cutting node at an oblique angle followed by a straight cut, away from the top bud. The cutting angle is very significant to identify the position, geotropism and phototropism of the propagating plant (Husen & Pal, 2007).
The oblique cut at the lower part is intended to provide a maximum area with cutting medium contact, so that root formation can be induced quickly under the influence of growth hormones that have already been supplemented with the rooting medium (Husen & Pal, 2007). On the other hand, the straight cut near the upper part of the cut is wrapped by the waxy material which is meant to reduce the chances of water loss due to transpiration. The most commonly used practice is to grow stem cuttings of conifers, gymnosperms and some oaks during the monsoon season.
The oblique cut in the lower part is made to promote callus formation, while the lower leaves, thorns are removed to avoid transpiration loss of water (Bonga, 2016). Furthermore, the positive properties of mother plants are transferred exactly to the newly grown plant, and in this way it provides the basis for creating several clones of the same healthy plant (Husen, 2004).
Chloris gayana
- Utilization of C. gayana
- Concerns about using the C. gayana
- Gulf countries and cultivating C. gayana
The nodes and internodes are present on the stem with tufted culms that are creepy or erect (Hussain et al., 2021). This phenomenon protects the field soil from pathogenic pests and promotes biodiversity (Abu-Alrub et al., 2018; Gupta & Arora, 2015). Experimental studies have demonstrated successful soil rejuvenation in Australian soil devastated by mining (Fox et al., 2004).
Moreover, it is resistant to several acidic or basic chemicals and metals, but not to manganese and magnesium (Allah & Bello, 2019; Faji et al., 2021). The field preparation involves repeated plowing to create suitable seed bed and immediate sowing (Arshad et al., 2016). It is adapted to a wide range of weather conditions, but it can tolerate occasional frost or extreme cold (Abebe et al., 2015; Manalil et al., 2020).
It was found that soil properties were improved and showed that treated wastewater can be significantly used for irrigation (Mazahrih et al., 2018). The government bodies are developing new strategies to upgrade the desalination system to meet the irrigation and domestic water needs (Hussain et al., 2021).

Methods and Materials
- Experiment setup
- Pot preparation and seed germination
- Growing Generation 1 plants
- Assessment of growth parameters in Generation 1 plants
- Evaluation of growth parameters in Generation 1 plants
- Propagation of Generation 2 through replantation
- Assessment of growth parameters in Generation 2 plants
- Experimental Design and Statistical Analyses
- Analyses performed
- Treatments Tested
Random individual plants were selected from each growth chamber (CO2 enriched and CO2 ambient temperature) and plant height, number of leaves and shoot/root ratio assessed at regular intervals. The plant shoot to root ratio was measured after the second month of growth and the assessment was made at regular intervals up to the fourth month of plant growth. For this purpose, the soil pots were prepared by filling the fertile soil compost and branches with their roots were removed from Generation 1 plants and replanted in the newly prepared soil pots.
27 plants were propagated in two successive groups of plants, i.e. replanted pots were divided into two CO2-enriched and CO2-supplied groups under controlled growth conditions in (BINDER Growth Chamber, KBW#720). Planted Generation 1 branches were able to grow Generation 2 plants under controlled CO2 supplies. Plant height, number of leaves, shoot/root ratio, chlorophyll content, stomatal conductance and photosynthetic rate were measured in 2nd generation plants.
Random individual plants were selected from each growth chamber and scored for plant height, number of leaves, and stem/root ratio. Stomatal conductance, photosynthetic rate, and chlorophyll content were measured at regular intervals using the same equipment and methodology as for Generation 1 plants.
Results
Growth pattern assessment - Generation 1 plants
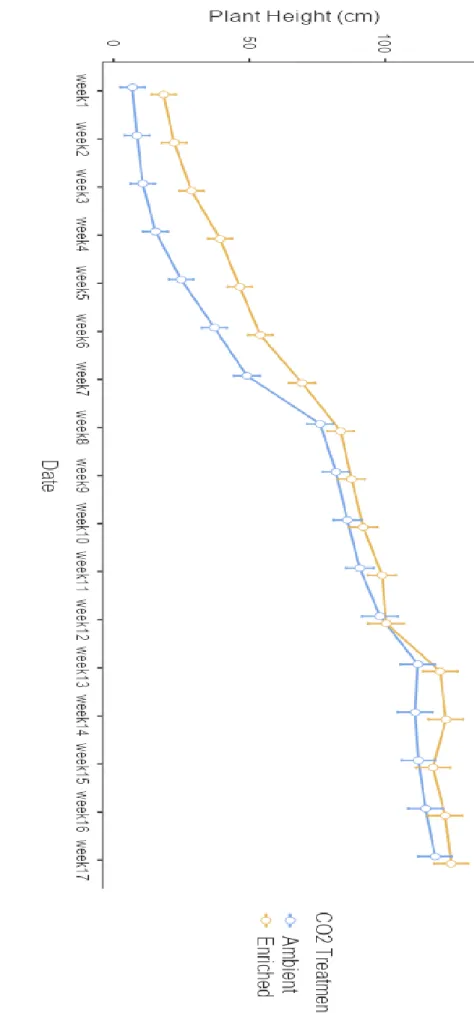
- Plant height
- Number of leaves
- Chlorophyll content
- Stomatal conductance
- Photosynthetic rate
- Shoot/Root ratio
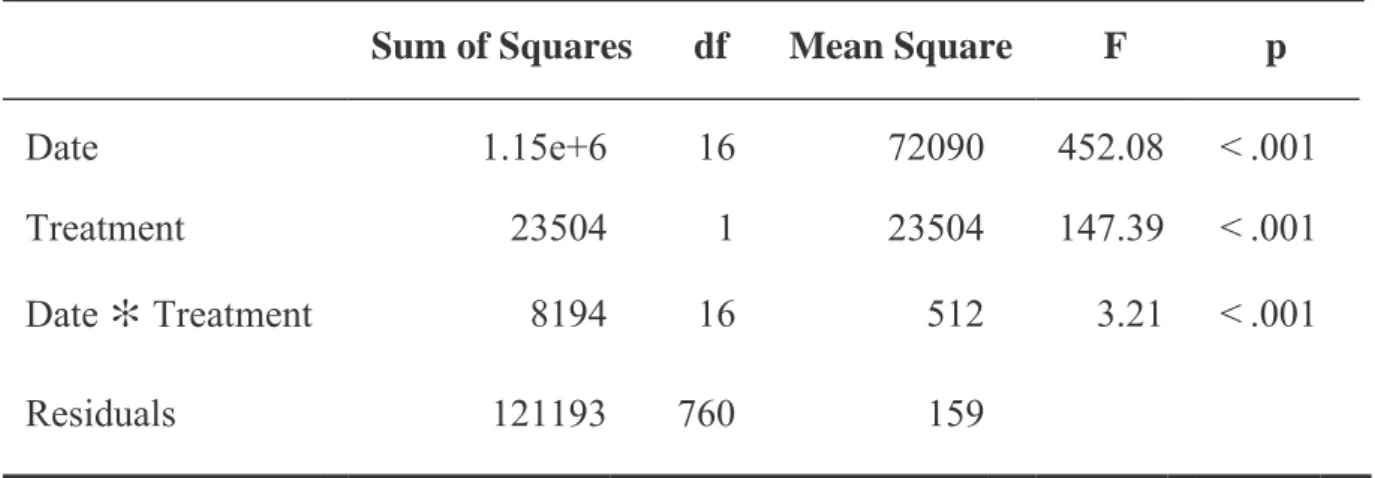
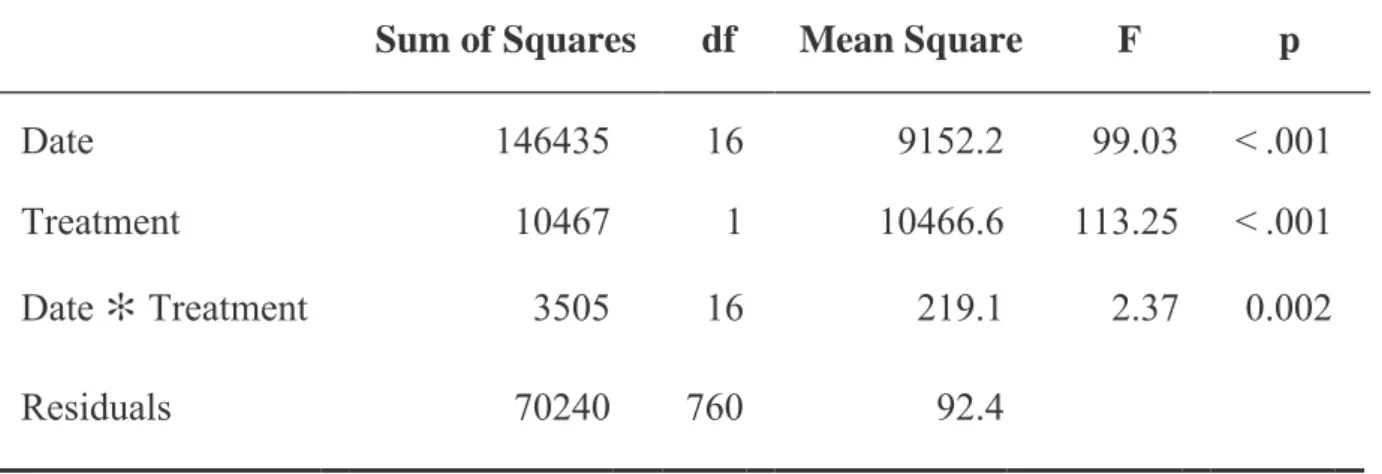
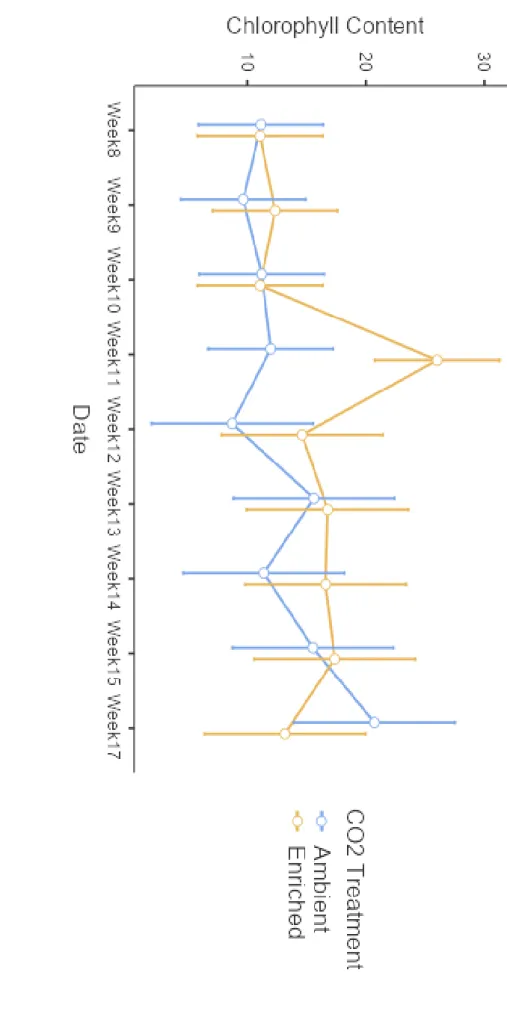
In general, chlorophyll content in plants grown under CO2-enriched conditions was greater than or equal to plants grown under ambient CO2 (Figure 7). Rhodes grass grown under CO2-enriched conditions reached the highest chlorophyll content at week 11 with an average of 26 units, while weeks 8 and 10 had the lowest chlorophyll content with an average of 11 units. Rhodes grass grown under ambient CO2 conditions had the highest chlorophyll content at week 17 with an average of 21 units and had the lowest chlorophyll content at week 12 with an average of 9 units.
The average rate of photosynthesis varied between 72 and 75% in Rhodoska grass grown in a CO2-enriched environment (Figure 9). In addition, the mean of plants grown under external CO2 at month 3 with a mean of 25.5 was significantly lower than plants grown under external CO2 at month 4 and enriched CO2 at month 3, which averaged 36 and 42, respectively. 6. However, plants grown under atmospheric CO2 at month 4 with a mean of 36 were significantly higher than plants grown under enriched CO2 at months 2 and 4 with means of 19.2 and 16.1.
Moreover, Rhodes grass grown under enriched CO2 in month 3 had an average of 42.6 and it was significantly higher than plants grown under enriched CO2 in months 2 and 4 with an average of 19.2 and 16.1. In general, the Rhodes grass grown under ambient CO2 conditions had a higher average dry shoot/root ratio compared to those grown under enriched CO2 conditions (Figure 11).
Growth pattern assessment – Generation 2 plants
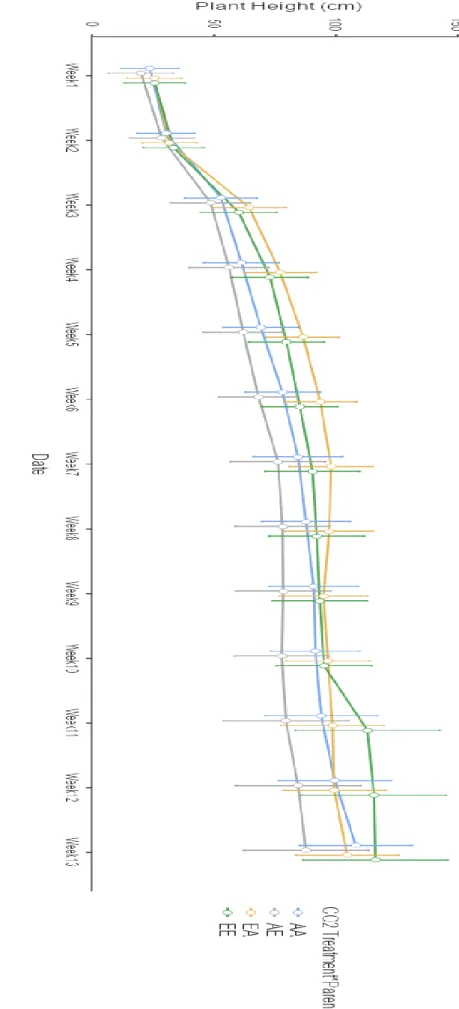
- Plants height
- Number of leaves
- Chlorophyll content
- Stomatal conductance
- Photosynthetic rate
- Shoot/Root ratio
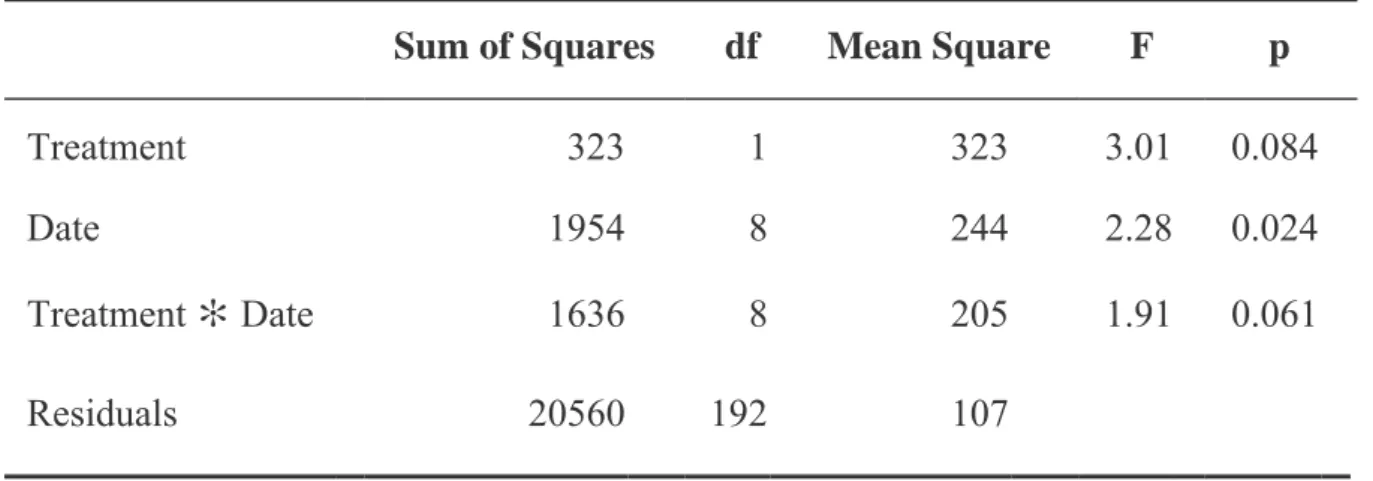
As shown in (Figure 13), the number of leaves generally increased during the experimental period and reached a maximum at week 13. A sharp decrease was observed at week 6 in group stomatal conductance (AA), which had a value of 227 mmol/m2s in week 5 (considered the maximum value) and became 111 mmol/m2s in week 6 (Figure 15). After analyzing the data, a significant difference (p<0.05) was observed between all experimental groups of plants (Table 12).
As shown in (Figure 16), plants whose parent comes from ambient CO2 were lower in photosynthesis rate than plants whose parent comes from enriched CO2 first. In the 4th month, the highest shoot/fresh root ratio with an average of 18.5 was observed in the (EE) group. While in month 2 the lowest Fresh Shoot/Root ratio with an average of 5.50 was for group (AE).
The highest shoot/dry root ratio with an average of 11 was observed in group (EE) at month 4. While the lowest shoot/dry root ratio was at month 2 with an average of 3.13 in group (EA ).
Discussions
The studies revealed that C3 plants under enriched CO2 showed more physiological development than those grown under the ambient CO2 level. The plant height and number of leaves were higher in enriched CO2 than in ambient CO2 as shown in the results. In the experiment, the chlorophyll content in plants grown under enriched CO2 conditions was higher than in plants grown under ambient CO2 in most weeks.
The dry stem/root ratio of plants under ambient conditions was higher than that under enriched conditions. The results are quite relevant to a previous study where an increased level of CO2 promoted plant height compared to those grown under ambient levels of CO2 (Ksiksi and Youssef, 2010). Therefore, maternal effects may confer effects on a plant's metabolism or photosynthetic rates (Tanaka et al., 2006).
The results of the current study are in line with the research by Ksiksi et al. 2018), who also reported increased stomatal conductance under CO2-enriched conditions. However, a possible reason for the non-significant differences in some parameters in generation 2 plants can be justified based on the plant's abiotic stress tolerance behavior.
Conclusion
Fluctuations in CO2 in Free Air CO2 Enrichment (FACE) inhibit photosynthesis, plant growth and yield. Responses of a C3 and C4 perennial grass to CO2 enrichment and climate change: Comparison between model predictions and experimental data. Characterization of physiological response of leaves of the high-yielding rice cultivar Takanari to CO2 enrichment in free air.
Effects of CO2 enrichment on photosynthesis, growth and biochemical composition of seagrass Thalassia hemprichii (Ehrenb.) Aschers. Growth and carbon ratios of mature Picea abies trees during 5 years of outdoor CO2 enrichment. Effects of CO2 enrichment on growth partitioning of Chloris gayana in the arid environment of the UAE.
Photosynthesis, plant growth and N allocation in transgenic rice plants with reduced Rubisco under CO2 enrichment. The effect of CO2 enrichment on the growth of a C3 weed (Parthenium hysterophorus L.) and its competitive interaction with a C4 grass (Cenchrus ciliaris L.).