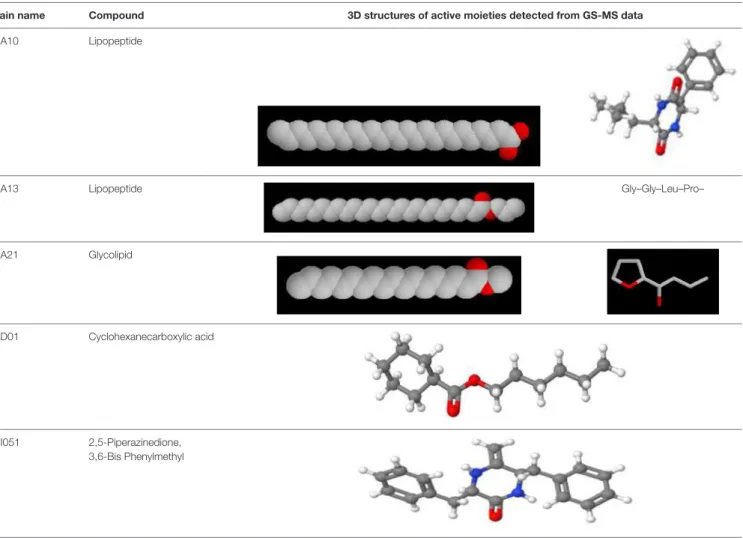
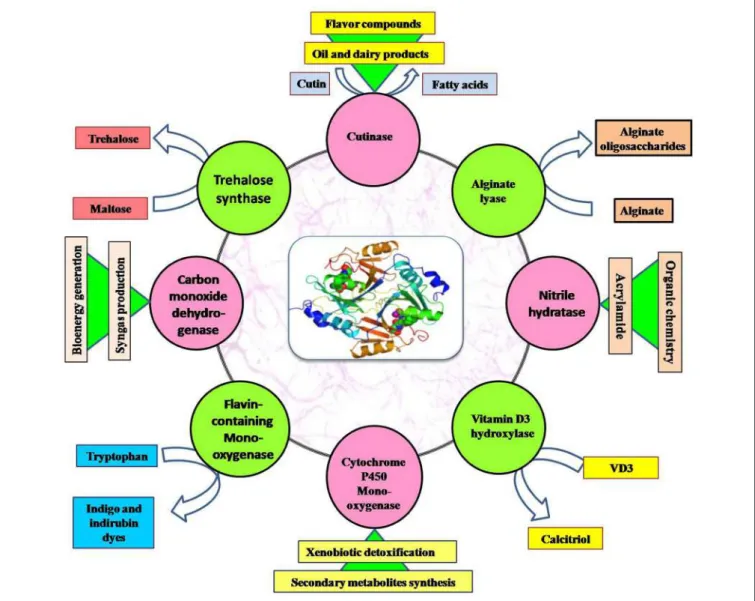
Another important secondary metabolite, carotenoids (tetraterpenoid) is produced by Dietzia natronolimnaea HS-1 (Gharibzahedi et al., 2014). Monofunctional enzymes can either act on poly(M) or poly(G), and bifunctional enzymes prefer poly(MG) (Tondervik et al., 2010).
INTRODUCTION
ACTINOBACTERIA AS THE OLDEST AND MOST PROMISING RESOURCE
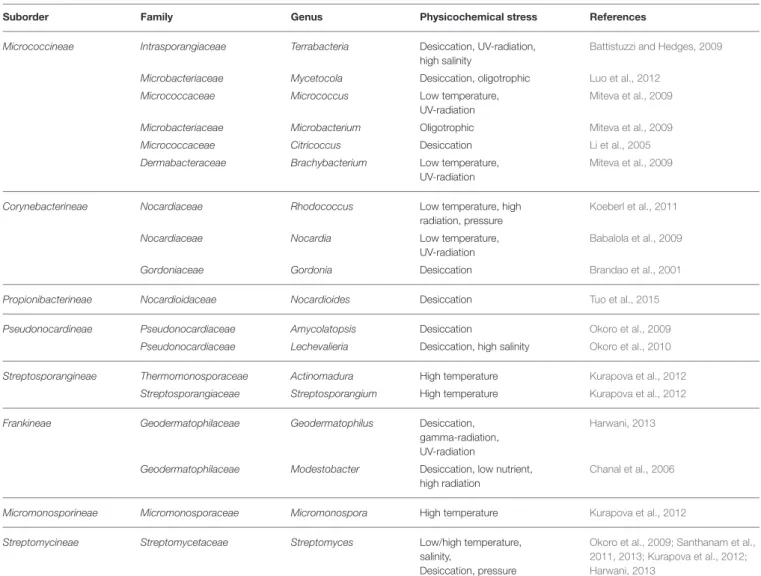
NEW SOURCE FOR EXTREMOPHILIC ACTINOBACTERIA
ARID HABITATS AND EXISTENCE OF BIOGEOGRAPHICAL BARRIERS
Arid zones are the interface between frequently vegetated semi-arid zones and biologically unproductive hyperarid deserts (Neilson et al., 2012). In addition, habitats other than soil are also considered as new resource areas with limited water availability (Azua-Bustos et al., 2012).
XEROPHILIC STRAINS ISOLATED FROM ARID AREAS
On the contrary, native desert bacteria can improve plant health in desert agroecosystems (Koeberl et al., 2011). Even within the hyperarid margin, fine variations in physicochemical parameters can have a strong effect on the taxonomic diversity of actinobacterial communities (Neilson et al., 2012).
BIOLOGICALLY ACTIVE METABOLITES REPORTED FROM XEROPHILIC
In this regard, a moderately thermophilic strain of Streptomyces fumigatiscleroticus 315 HE578745 that was isolated from the desert soil was experimentally shown to be xerotolerant (Lubsanova et al., 2014). These strains exhibited plant growth promoting activity and intrinsic water stress tolerance (-0.05 to -0.73 MPa) (Yandigeri et al., 2012).
ACTINOBACTERIA
Another Streptomyces strain isolated from the Chilean highland soil of the Atacama Desert produces novel aminobenzoquinones that show inhibitory activity against bacteria and dermatophytic fungi (Schulz et al., 2011). NRPS genes were widely distributed among these taxa, while PKS-I genes were detected in fewer genera (Meklat et al., 2011).
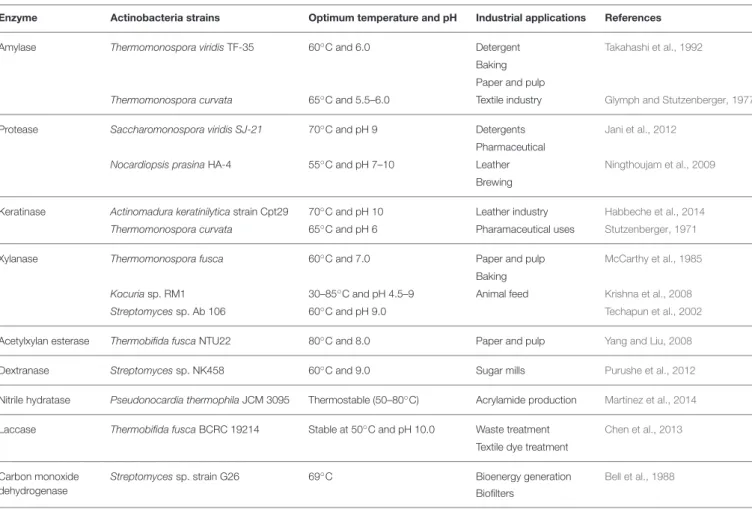
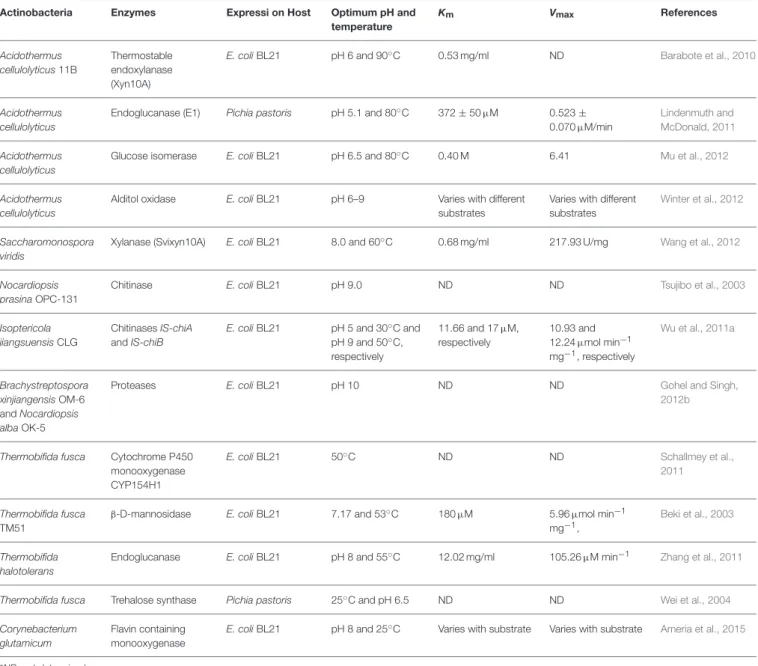
ENZYMES REPORTED FROM XEROPHILIC ACTINOBACTERIA
ACTINOBACTERIA FROM ARID REGIONS OF IRAN AND THEIR POTENTIAL
BIOTECHNOLOGICAL ACTIVITIES
Although studies of the world's deserts are increasing, information on the diversity of Actinobacteria in the arid regions of Iran is scarce. Until now, only four new species of Actinobacteria, which belonged to the genera Nocardiopsis, Kribbella and Promicromonospora, have been reported from the semi-arid soil of Iran (Hamedi et al., 2011; Mohammadipanah et al.
DISCUSSION
Only a few members of Actinobacteria have been introduced from the arid regions of Iran, and their secondary metabolite potential is still being investigated. Further focus on indigenous actinobacteria from Iranian deserts would increase our knowledge of their occurrence, distribution, ecology, taxonomy and biotechnological potential.
FUTURE PERSPECTIVE
Radiation protection, sulfur cycle or regulation of the community structure through quorum sensing and the production of bioactive compounds are among the ecological functions of these bacteria in such ecosystems (Bull and Asenjo, 2013). In desert habitats, the availability of water and organic substrates are among the main parameters limiting the ability of bacteria to maintain their metabolic functions (Saul-Tcherkas et al., 2013).
AUTHOR CONTRIBUTIONS
Drought and radiation are determinants of life, while habitat availability, temperature, pH and toxicants (high local concentrations of elements such as arsenic) are some of the major determinants. Organic substrates may derive from the chemical profile of plant root exudates, leading to variability in the associated bacterial composition of the dry soil (Saul-Tcherkas et al., 2013).
Life on the hyperarid edge: new bacterial diversity in the arid soils of the Atacama Desert, Chile. Extremophiles. Diversity of cultivable actinomycetes in the hyper-arid soils of the Atacama Desert, Chile. Antonie van Leeuwenhoek nov., isolated from hyper arid soils. Int.
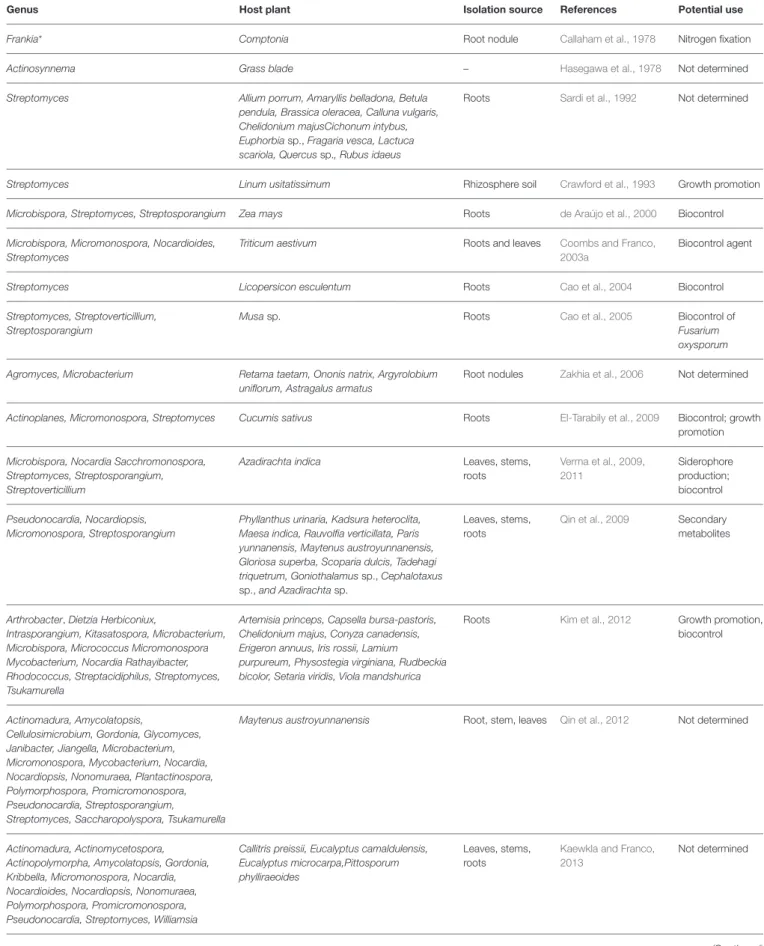
PLANT-ASSOCIATED ACTINOBACTERIA
Microbial communities present in soil represent the richest reservoir of biological diversity on our planet (Berendsen et al., 2012). Individual isolates representing the genera Arthrobacter, Dietzia, Herbiconiux, Kitasatospora, Mycobacterium, Nocardia, Rathayibacter and Tsukamurella were also found (Kim et al., 2012).
MICROMONOSPORA AND NITROGEN FIXING NODULES: A UNIVERSAL
Actinobacterial species identified in the genera Agromyces, Curtobacterium, Microbacterium, Micromonospora and Streptomyces have been reported from nodule tissues (Sturz et al., 1997;. With the exception of the study of Valdés et al. 2005), the isolation of Micromonosporaal from nodules also followed same isolation protocols as that of legumes, using yeast-mannitol agar as isolation medium (Vincent, 1970).
DISTRIBUTION, LOCALIZATION AND GENETIC DIVERSITY OF
The isolation of micromonospora strains usually requires selective isolation procedures to promote its slow growth, but in all the above examples the same isolation protocol as that used for the isolation of rhizobia was used (Cerda, 2008; . Rodríguez, 2008; Carro, 2009 ; Alonso de la Vega, 2010; Trujillo et al., 2010). Actinorhizal plants sampled to date in Mexico, Spain, Canada and France include the species Alnus viridis, Casuarina equisetifolia, Coriaria myrtifolia, Elaeagnus x ebbingei, Hippophae rhamnoides, Myrica gale and Morella pensylvanica et al., 05; .
MICROMONOSPORA IN NITROGEN FIXING NODULES
However, for this plant, at least one species of Micromonospora was found from every nodule sampled (Carro et al., 2012a). In a recent study, Carro et al. 2013a) examined several actinorhizal plants and recorded the number of strains and species of Micromonospora found.

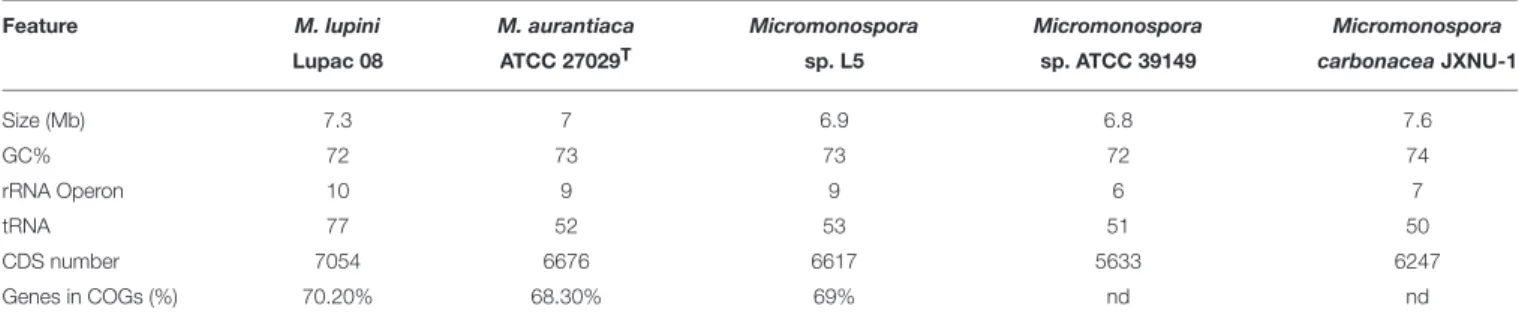
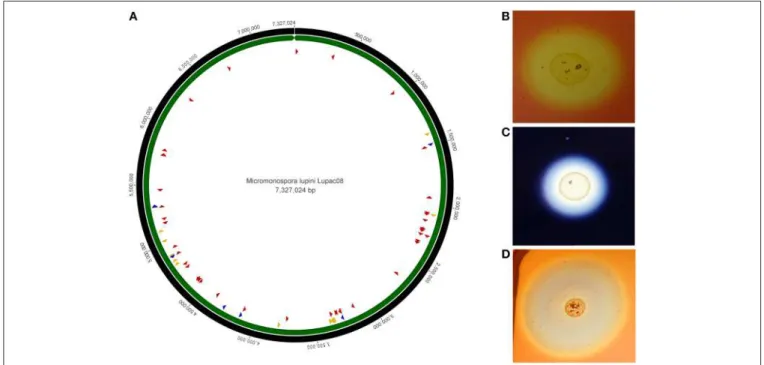
GENOME FEATURES OF
MICROMONOSPORA ISOLATED FROM NODULES
MICROMONOSPORA LUPINI LUPAC 08: A FRIENDLY BACTERIUM HIGHLY
EQUIPPED WITH PLANT CELL WALL DEGRADING ENZYMES
Plant polymer-degrading enzymes are thought to be involved in internal plant colonization (Compant et al., 2005). BH72 affected the colonization of the roots when exposed to the plant's root exudates (Shidore et al., 2012).
MICROMONOSPORA, A PLANT GROWTH PROMOTER WITH WIDE APPLICATION IN
THE MICROMONOSPORA METABOLOME AND ITS POTENTIAL ROLE IN
PLANT-MICROBE COMMUNICATION SIGNALS
In this work, it was suggested that some of these compounds could act as communication molecules to establish the symbiotic interaction between Franki and the host plant (Udwary et al., 2011). These compounds can act as communication molecules between the microorganism and the plant to allow bacterial colonization (Udwary et al., 2011).
CONCLUDING REMARKS
Based on these results, the authors suggested that different metabolites were formed under the two tested conditions and that these compounds are responsible for the activation of different defense mechanisms in the plant (Conn et al., 2008). In addition, some metabolites may be required for nutrient uptake (Barry and Challis, 2009; Rungin et al., 2012). All these areas remain to be studied in the interaction of micromonospore and nitrogen-fixing plants.
ACKNOWLEDGMENTS
Generic diversity of limestone isolates consisted of Streptomyces (15 phylotype groups), Micromonospora (4), Amycolatopsis (3), Arthrobacter (3), Kitasatospora (2), Janibacter (1), Nocardia (1), Pseudonocardia (1) and Rhodococcus (1). The results show that 86% of the strains isolated from the Hundung limestone deposit sites possessed biosynthetic gene clusters, of which 40% exhibited antimicrobial activities.
Introduction
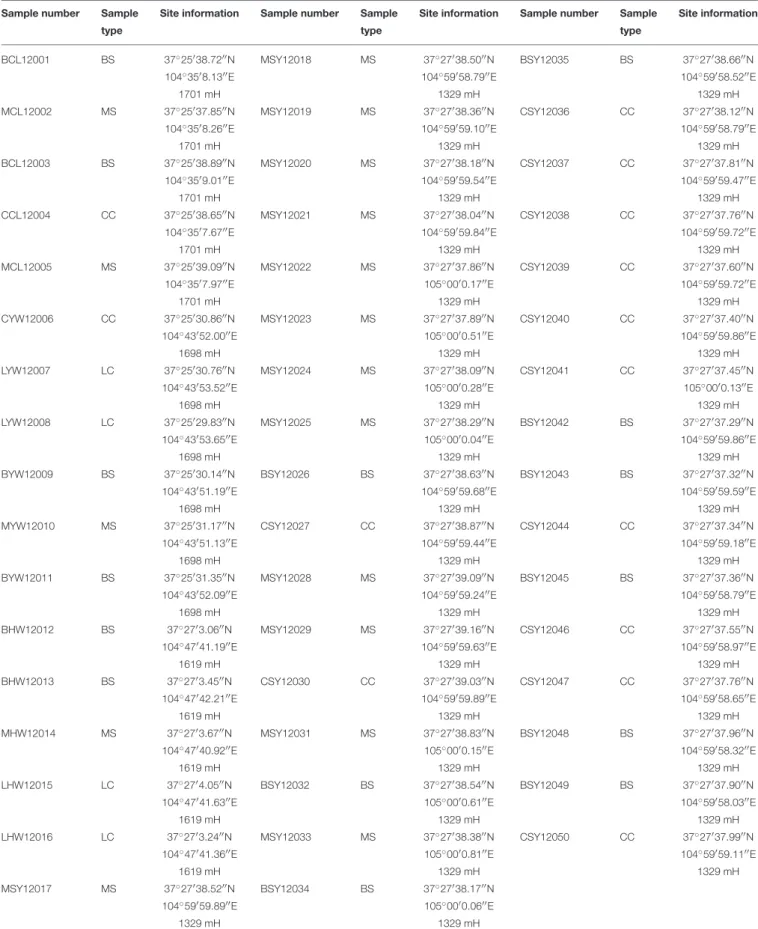
Out of the 137 actinobacterial isolates, 118 were found to have at least one of the three biosynthetic gene clusters (PKS-I, PKS-II, NRPS). This paper reports the actinobacterial diversity profiling of the Hundung limestone depositional sites using ARDRA as the molecular tool for preliminary classification.
Materials and Methods
Actinobacteria involved in the deterioration of artworks are considered serious risk factors if environmental changes promote their massive spread (Groth et al., 1999b; Portillo et al., 2009). Three sets of degenerate primers were used for amplification of PKS-I, PKS-II, and NRPS-specific domains ( Metsä-Ketalä et al., 1999 ; González et al., 2005 ).
Results
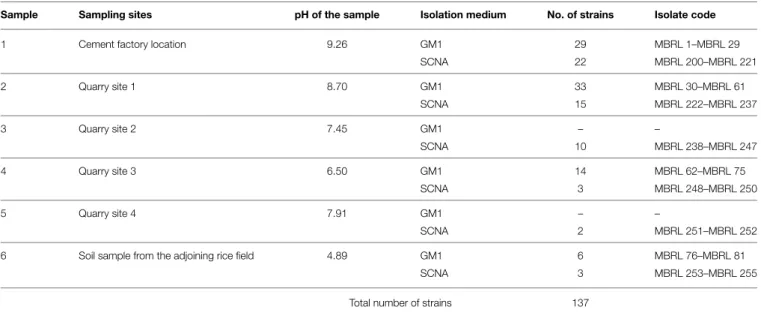
Forty-five actinobacterial strains from the Hundung limestone habitat showed biocontrol activities against at least one of the rice fungal pathogens. The remaining 75 strains had two or more of the biosynthetic gene clusters: 9 strains possessed both PKS-I and PKS-II; 8 had both PKS-I and NRPS; 36 had both PKS-II and NRPS while 22 strains had all three sets of biosynthetic genes. Table 4 shows the amplification profile for biosynthetic genes in Actinobacteria Hundung (see Supplementary Table S4 for the complete PCR profile of biosynthetic genes).
Discussion
For the repertoire of the actinobacterial diversity in caves, Nakaew et al. 2009) added the genera Nonomuraea, Actinocorallia, Catellatospora, Microbispora and Sprillospora. In our studies, 118 of the 137 actinobacterial isolates were found to have at least one of the three biosynthetic gene clusters. The results of the various experiments suggest that 86% of the strains isolated from Hundung limestone rocks possessed biosynthetic gene clusters, of which 40% exhibited antimicrobial activities.
Author Contributions
Acknowledgments
Supplementary Material
This number is much lower than for the cultivated genera (60 genera) (Abdelmohsen et al., 2014a).
MATERIALS AND METHODS
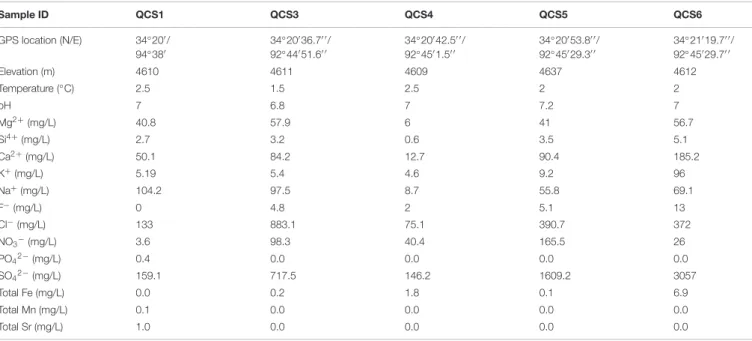
Previously, one 16S rRNA gene-based microbial study showed the presence of Actinobacteria in the cold springs of Wuli, QTP (Li et al., 2012). However, the actinobacterial diversity in these cold springs may be underrepresented due to the use of universal bacterial primers (Cottrell and Kirchman, 2000; Jiang et al., 2010a). Alpha diversity indices, such as Simpson, Shannon, Equitability and Chao 1, were calculated using the PAST software package (Hammer et al., 2001).
RESULTS
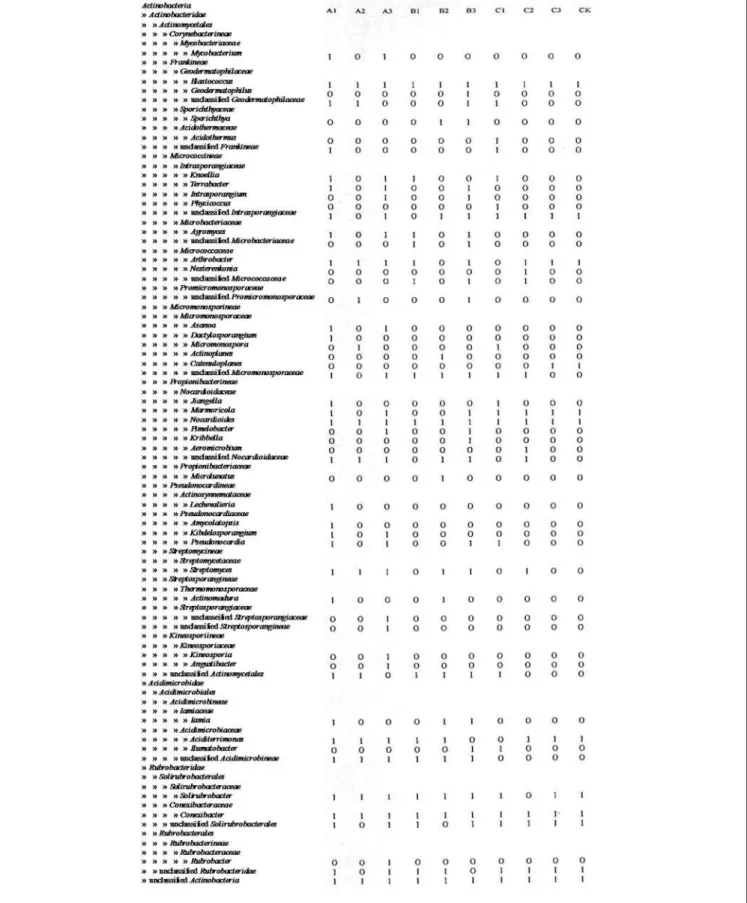
The underlying coal or gas hydrate may provide abundant nutrients, which support diverse actinobacterial communities in the cold springs studied (Santos et al., 2008; Jiang et al., 2010a). Actinobacterial clones obtained from Tibetan salt lakes were mainly classified with Micrococcales, Propionibacteriales and Frankiales (Jiang et al., 2010a). In contrast, actinobacterial communities in Tengchong hot springs were mainly associated with unclassified Actinobacteria, Rubrobacterales and Frankiales (Song et al., 2009).
SUPPLEMENTARY MATERIAL
This comparison represents one of the largest sequencing efforts of Actinobacteria in volcanic caves to date. Samples from Helmcken Falls Cave (Canada) were prepared, processed and observed at the University of British Columbia (UBC) BioImaging Facility (Cheeptham et al., 2013). The raw pyrosequencing reads were processed using version 1.34 of Mothur's software package (Schloss et al., 2009).

RESULTS AND DISCUSSIONS
The subgroup of Actinobacteria was identified with RDP classifier (Maidak et al., 2001) and used for further analysis. Similar spherical particles were previously reported by Cuezva et al. 2013) who suggested vaterite as their mineralogical phase. Furthermore, studies of Actinobacteria in other environments have been shown to show endemism (Wawrik et al., 2007; Valverde et al., 2012).
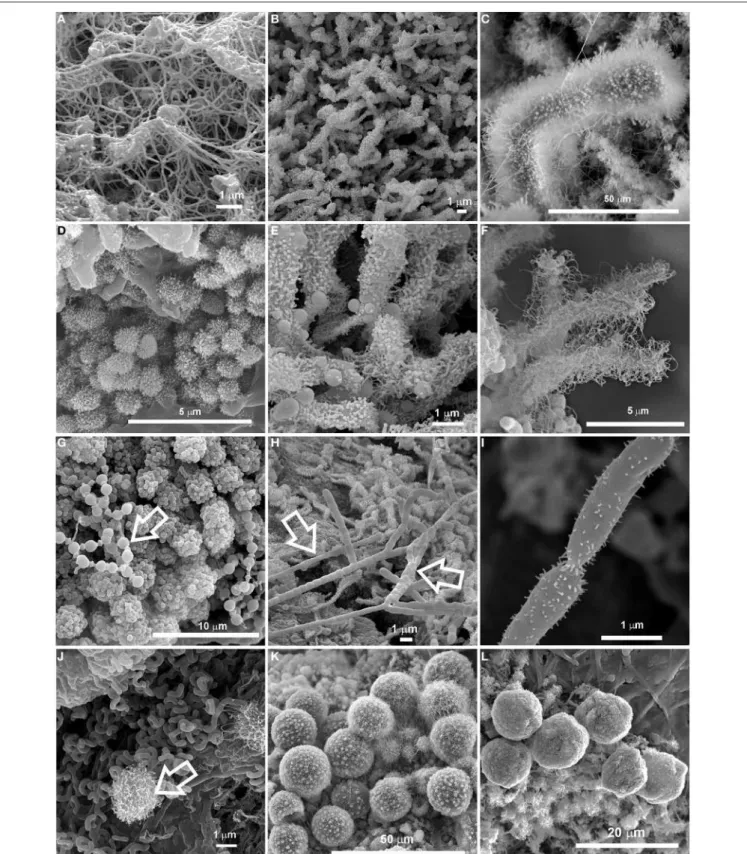
CONCLUSIONS
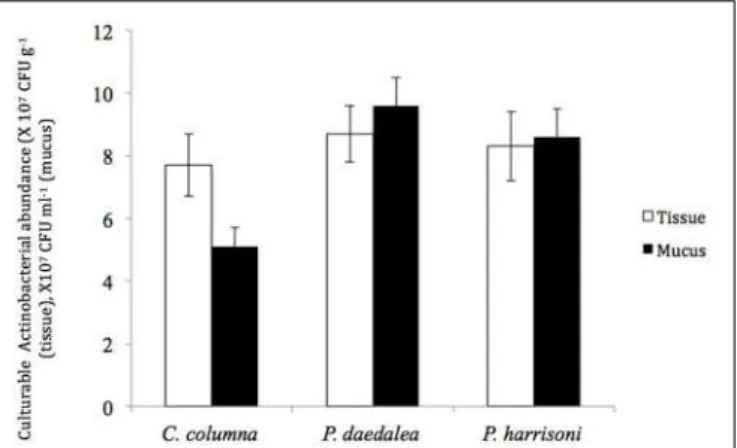
Molecular analysis of the benthic microbial community in the Zavarzin thermal spring (Uzon caldera, Kamchatka, Russia). Differences in abundance and diversity of Actinobacteria were detected between mucus and tissue of the same coral host. In addition, temporal and spatial variations in the abundance and diversity of culturable actinobacterial communities were detected.
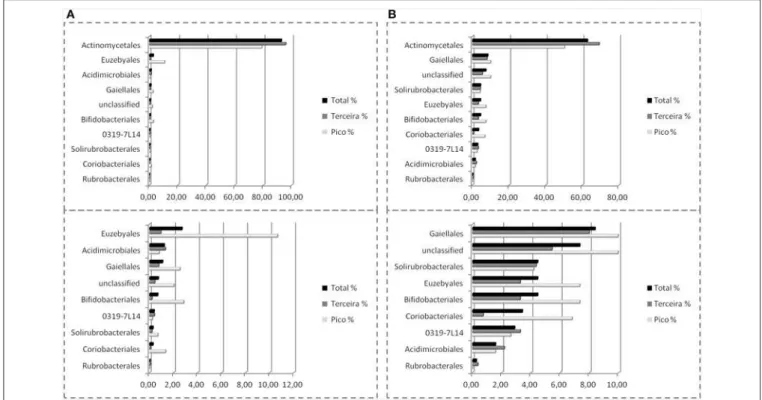
MATERIALS AND METHODS Sampling and Sample Processing
It was important to also quantify the total number of microbes in the coral tissue and slime examined. Significant differences in the number of culturable Actinobacteria were obtained between mucus and tissue samples from the same coral. Brevibacterium and Kocuria were the most dominant actinobacterial isolates in the coral tissue and slime samples examined.
CONCLUSION
Guppy and Bythell, 2006) either by degradation of the coral mucus with extracellular proteases (Bourne and Munn, 2005) or by a reduction in the antibiotic content of the coral mucus (Ritchie, 2006). suggested by Reshef et al. 2006) may explain the changes in P actinobacterial abundance and diversity. The first primer set, Com2xf/Ac1186 (Schäfer et al., 2010), was used to detect most Actinobacteria species . The 16S rRNA gene sequences were taxonomically assigned using the Ribosomal Database Project II Naïve Bayesian rRNA classifier (RDP; Wang et al., 2007).
RESULT
The structure of the bioactive compound(s) was analyzed using mass spectrometry (MS) and nuclear magnetic resonance (NMR) spectroscopy (Booth et al., 1976; Hamza et al., 2013). The maximum activity was found against MRSE ATCC 35984 (inhibition zone of 35 mm diameter), followed by A. The detailed antimicrobial profiles are shown in Table 1. These morphological features are consistent with grouping the strain into the genus Streptomyces (Williams et al. ., 1989).
RESULTS AND DISCUSSION Phenotypic Analyses of Strain
An independent T-test analysis was also performed to compare the effect of the extract against cancer and normal cell line. In this regard, it has prompted the needs to perform the profiling of the chemical components of the MUM256 extract. The phylogenetic tree of the deduced AA sequences of PKS II segments and ketosynthase genes was generated using the neighbor-joining method through MEGA programs (Kumar et al., 2004).

RESULTS AND DISCUSSION
The AA composition was also predicted to underpin the function of the type II PKS and ketosynthase of our interest. All these three iterative KS domains of PKS type II gene clusters of actinomycetes were modeled using comparative modeling method. The secondary structures of type II PKS and ketosynthase domains of 3D models were created using a (PS)2 is an automated homology modeling server (Chen et al., 2006).