Biometric and eddy covariance methods to study the carbon balance of Larix principis-rupprechtii forest in Qinling Mountains, China. Currently, the most important techniques for measuring NEE are the eddy covariance technique and the biometric technique. The eddy covariance technique has several advantages: (1) it is non-destructive and has a low workload, (2) it provides observations at the ecosystem level, and (3) it yields continuous records that address time scales of every half hour up to the length of the data record [16,20].
This paper presents the first set of results from a long-term project measuring CO2 exchange between forest and atmosphere using vortex covariance and biometric methods in a L. principis-rupprechtii forest in the Qinling Mountains.
Materials and Methods 1. Study Area
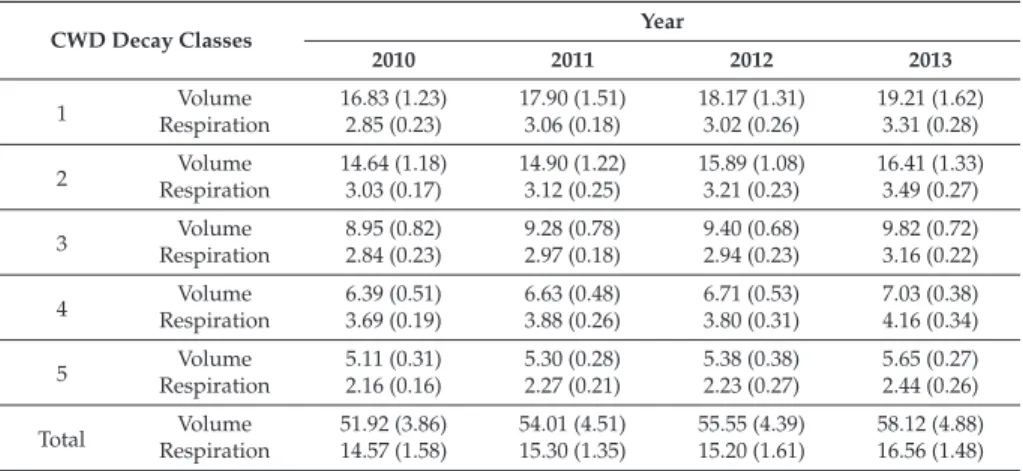
Soil respiration in the trench plot was measured by the same soil respiration method. To scale up the chamber measurements of CWD respiration to the standing level, we calculated the volume of the five decay classes of CWD in the graph. To scale up the room measurements of stem respiration to the stand level, we estimated the total sapwood volume per unit of soil area in the plot.
To scale up room measurements of leaf respiration to the stand level, we estimated the total leaf dry biomass per unit of soil area in the plot.
Results
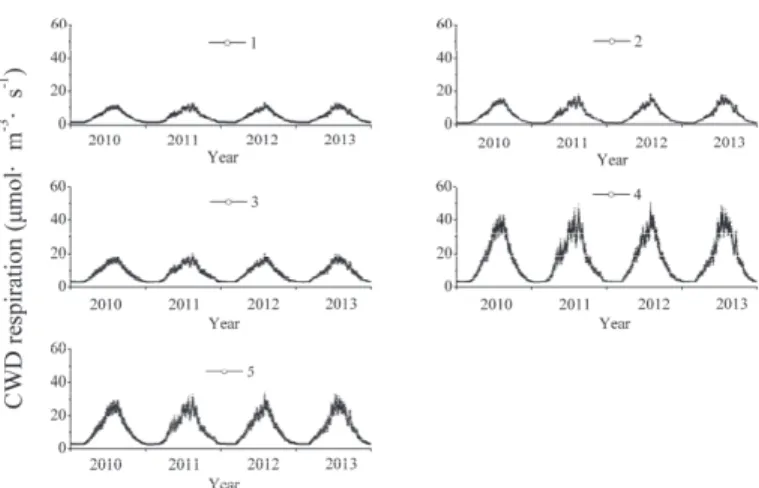
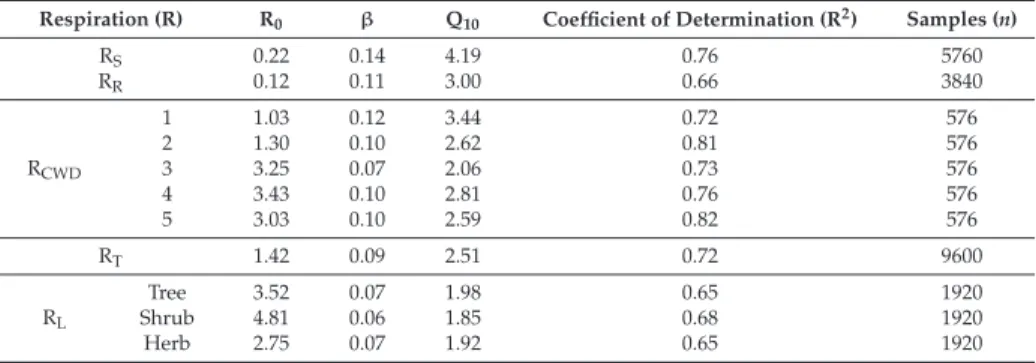
There was a strong exponential correlation between respiration of different CWD decay classes and temperature (Table 1). The lowest leaf respiration in trees, shrubs, and herbs occurred in March, increased sharply until July, and then decreased until November (Figure 4). Cumulative leaf respiration per unit ground area for trees, shrubs, and herbs showed that tree leaves have higher respiration than shrub and herbaceous leaves (Table 3), which mainly corresponds to the highest leaf biomass for trees.
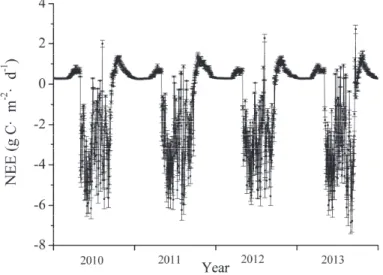
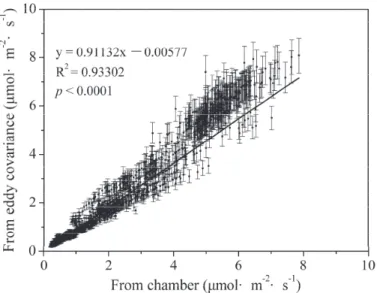
Average daily ecosystem respiration based on chamber measurements and eddy covariance measurements are shown in Figure 7 .
Discussion
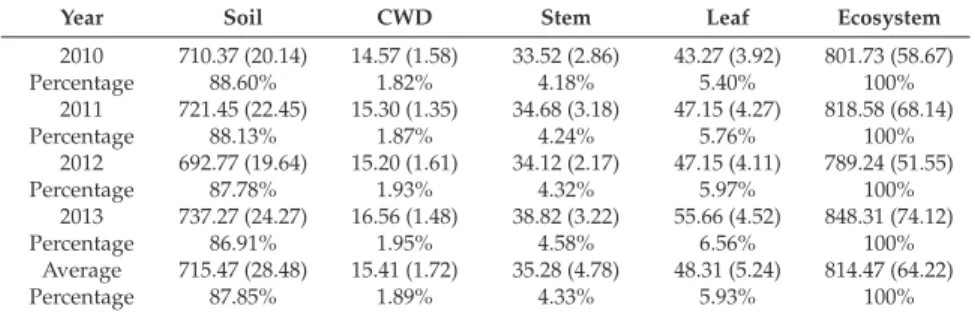
Daily mean ecosystem respiration measured by eddy covariance and chamber methods is shown in Figure 7. Our study showed that annual mean ecosystem respiration measured by chamber method was 5% higher than that measured by eddy covariance. Our study showed that the NEE measured by the eddy covariance method was 14% higher than that measured by the biometric method, which is consistent with Wang et al.
However, there were still sources of error for measuring the NO based on vortex covariance measurements.
Conclusions
Comparison of eddy covariance and chamber-based methods for measuring CO2 flux in a temperate mixed forest.Tree Physiol. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest.Tree Physiol. Biometric and eddy-covariance-based estimates of carbon balance for a warm-temperate mixed forest in Japan.Agric.
The use of eddy covariance to infer net carbon dioxide uptake by the Brazilian rainforest.Glob.
Carbon Mass Change and Its Drivers in a Boreal Coniferous Forest in the Qilian Mountains, China
Introduction
Carbon mass change and its drivers in a boreal coniferous forest in the Qilian Mountains, China. These climate change-induced species conversions will lead to a long-term loss of carbon stock in montane forest ecosystems [ 15 ]. The response of mountain forests to climate change depends greatly on relative changes in temperature, precipitation, CO2 and their interaction effect [10]; the relative contributions of individual historical drivers can be assessed through high-resolution climate data and modeling [16,17].
However, analyzes of the driving factors of carbon storage changes in recent years in high latitudes and high altitude areas are still lacking [18]. The sensitivity to climate change of carbon storage in montane forests is based on initial climate variability; Thus, the increase in average annual temperature at high altitudes results in the sensitivity of tree growth and changes in forest productivity [3]. Investigating the sensitivity of carbon flux to climate change will reduce the uncertainty of past drivers of climate change and help guide forest management.
For a better understanding of the effects of climate change on boreal forests, it is necessary to quantify the drivers of carbon changes in boreal forests during the last 50 years. Montane boreal forest is the dominant forest type, and it plays important roles in the hydrology and biogeochemistry of the area due to high altitudes m) and minimal human-induced ecological degradation [23]. Ecosystem stability of the montane boreal forest in the Qilian Mountains supports downstream economy and production because the forest serves as the most important water conservation entity in the area.
Better understanding of the role of climate and atmospheric CO2 in determining carbon flux in this montane boreal forest, and the relationship between the nuclear power plant and biomass is critical for guiding ecological policy to optimize carbon storage. To address this knowledge need, we used climate and CO2 concentration data to reconstruct carbon storage in living biomass for the montane boreal forest in the Qilian Mountains, China from 1964–2013.
Materials and Models
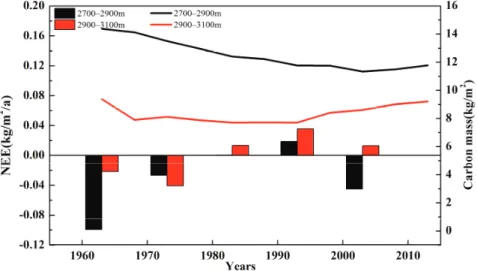
Because the climate variables are strongly interrelated, a comparison of the effects of climate change on carbon fluxes based on the change of a single parameter in the data was incomplete [6,16]. Indeed, in the simulation period, the NPP and carbon mass of the montane boreal forest increased. The temperature sensitivity of biomass allocation may be an important but not obvious regulator of the carbon cycle in the boreal forest [19,56].
Response of tree recruitment to climatic variability in the alpine treeline ecotone of the Qilian Mountains, northwest China.For. Carbon pools of semi-arid Picea crassifolia forests in the Qilian Mountains (northeastern Tibetan Plateau).For. Gis-assisted modeling of the spatial distribution of Qinghai spruce (Picea crassifolia) in the Qilian Mountains, Northwest China based on biophysical parameters.
Estimation of aboveground biomass of montane forest in the upper Heihe River basin using landsat-tm data.Int. RFF represented an average of 4–13% of total soil respiration, and forest floor removal increased mineral soil moisture content. The forest floor mass is an estimate based on a companion study in the same pits [35] and the mass from the actual collars in this study.
Like mineral soils, forest soil respiration rates are negatively related to substrate C:N ratio [ 4 ] and three of our four stands had relatively high forest soil C:N ratios. If anything, the increased soil moisture in the no-litter treatment may have partially compensated for the reduction in RS associated with the absence of decaying forest floors. If the difference in soil respiration rates between no-litter collars and control collars were entirely due to the absence of contributions from forest floor respiration, we would expect a similar difference to be evident in the removable bag treatment.
Biotic and abiotic factors regulating forest floor CO2 flux across a range of forest age classes in the southern Appalachians. Pedobiology.
Carbon Sequestration in Protected Areas
Discussion 1. Carbon Content
This conservation depends on the amount of existing biomass, which depends on the age, diameter and height of the trees. These differences can be attributed to several factors such as the quality of the studied area, and the density and age of the trees, among others. For the production of coetane masses, the content is variable and depends on the age, diameter, height and density of the trees in a given location.
Forest ecosystems store carbon and are considered as an option to mitigate the effects of increasing atmospheric CO2 loading [59–62]. As mass age increases, increments decrease [45], and as a result, carbon sequestration potential is also affected. Compared to protected forests, sustainably logged forest areas capture more carbon [65], which is mainly a result of managing the age factor within the masses.
The differences between these studies and the estimated rate in this study can be attributed to factors such as: forest management, location of the studied area and species. The use of high-resolution satellite images combined with analysis of physical aspects of the terrain enabled detailed zoning directly related to the density of the forest mass and its amount of biomass and carbon. In this case, because it is a protected forest where forestry activities are limited, the productivity of the mass is lower and consequently the rate of carbon sequestration decreases.
The quantified carbon additionality is a parameter that depends on several factors, for which the age of the forest mass is considered to be one of the most important, and in the young and vigorous forests the sequestration rate is higher than in forests with advanced stages of development. Temporal evolution of carbon budgets in Appalachian forests of the United States from 1972 to 2000. For.
Nonlinear Variations of Net Primary Productivity and Its Relationship with Climate and Vegetation
The MODIS annual NPP algorithm relies on the summation of the daily estimate of gross primary productivity (GPP) calculated globally minus growth and maintenance respiration [14]. The best estimated result was then considered in the following analysis of the NPP relationship with climatic factors and vegetation phenology. This method did not require the independence and normality of the time series data [47], which has been widely used in trend analysis [19].
Similarly, 88.12% of all pixels showed an increasing trend on the Tibetan Plateau, where the increasing magnitude mostly occurred at 0–0.4 × 10–2 kg C m–2a–1 (Figure 4f). To clarify the characteristics of NPP trends, the average NPP and breakthrough year trends at the national and regional levels were calculated during the study period (Figure 5). At the regional level, 62.97% of the total pixels in northern China showed a negative relationship between NPP and temperature, while precipitation, solar radiation, and LOS generally showed a positive relationship with NPP (Table 1).
More than 57% of the total pixels in South China indicated a positive relationship between the NPP and climatic factors as well as LOS. Specifically, more than 83% of the total pixels for temperature and about 58% of the total pixels for precipitation showed a positive relationship with the NPP, while in general an opposite relationship occurred between the NPP and solar radiation. Moreover, the effect of climatic factors and LOS on NPP was 0.63, explaining a total of 56% of the variation in NPP.
For example, the Miami model is one of the first global empirical models to use only temperature or precipitation in the model [55], while the Thornthwaite Memorial model determines NPP for a specific location as the actual evapotranspiration functions [36]. The sparse distribution of climate data has limited the detail and accuracy of the relationship between climate factors and plant productivity. Climate impacts on net primary productivity trends in natural and managed ecosystems of the central and eastern United States.Agric.
Improvements to the global gross and net terrestrial primary production dataset. Remote Sens.
![Figure S2). The Kriging method was used for the spatial interpolation of climate data across the study area [41].](https://thumb-ap.123doks.com/thumbv2/1libvncom/9201242.0/79.723.264.462.458.524/figure-kriging-method-used-spatial-interpolation-climate-study.webp)