This culminated in exploring the mechanism of action of DCUN1D1 in prostate cancer using changes in protein expression measured by western blot analysis. We also observed that knockdown of DCUN1D1 in prostate cancer led to dysregulation of the ubiquitination and WNT/β-catenin pathways.
Introduction
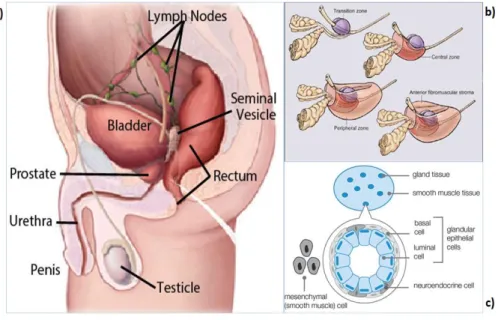
Prostate Cancer
- Background
- Prostate cancer screening and diagnosis
- Prostate cancer risk stratification
- Prostate cancer staging
- Treating prostate cancer
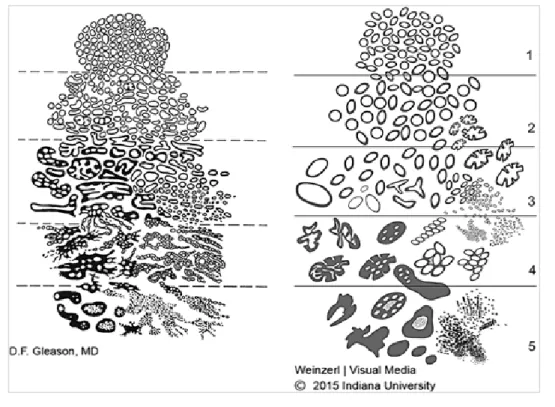
However, the TNM staging system provides guidelines for PCa classification based on clinical and pathological information (see Figure 5 below for schematic diagrams depicting the different classifications) (Edge SB et al., 2010). Tumor size (T) is classified based on the ability to determine the presence of PCa via DRE or examination after surgical removal of the prostate gland (Edge SB et al., 2010).
Proteomics
- Introduction to proteomics
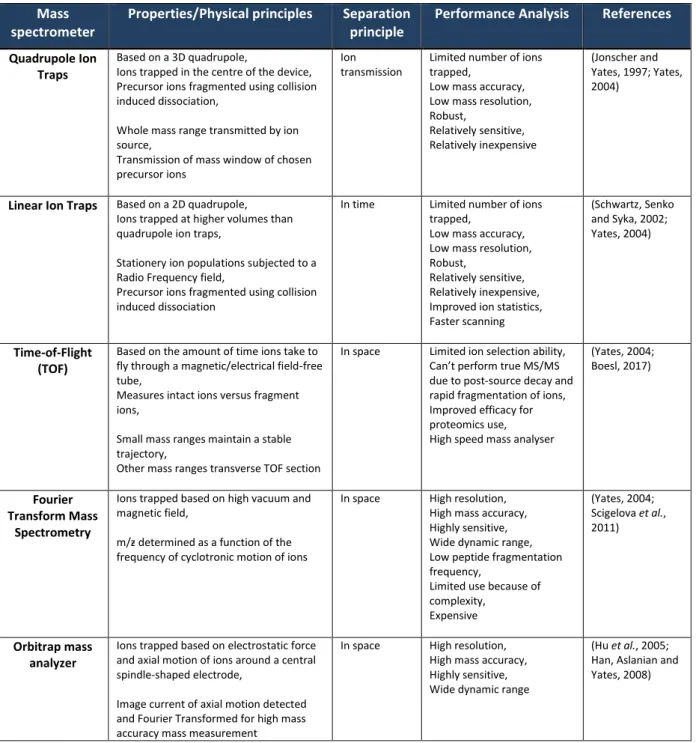
- Mass spectrometry
However, Fourier transform mass spectrometry has made the greatest contribution to mass precision measurement so far (Scigelova et al., 2011). Among the challenges to understanding the proteome is the ability to identify peptides/proteins, and databases of previously identified peptide sequences have played a key role in MS-based proteomics (Ma, 2010; Perez-Riverol et al., 2015 ).
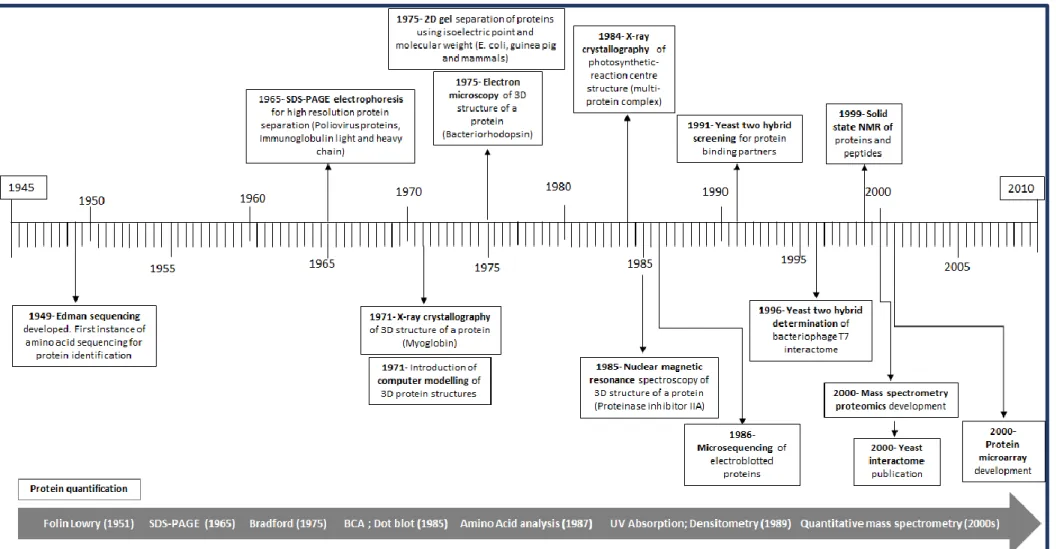
Posttranslational modifications, cancer and proteomics
Post-translational modifications are extensively covered in the Hallmarks of Cancer because of the wide range of cellular activities they affect, but also because of the dynamic benefits they provide during tumorigenesis, including the rate of signal transduction. MS-based proteomics have had a significant impact on the study of PTMs due to the high sensitivity and specificity of PTM detection.
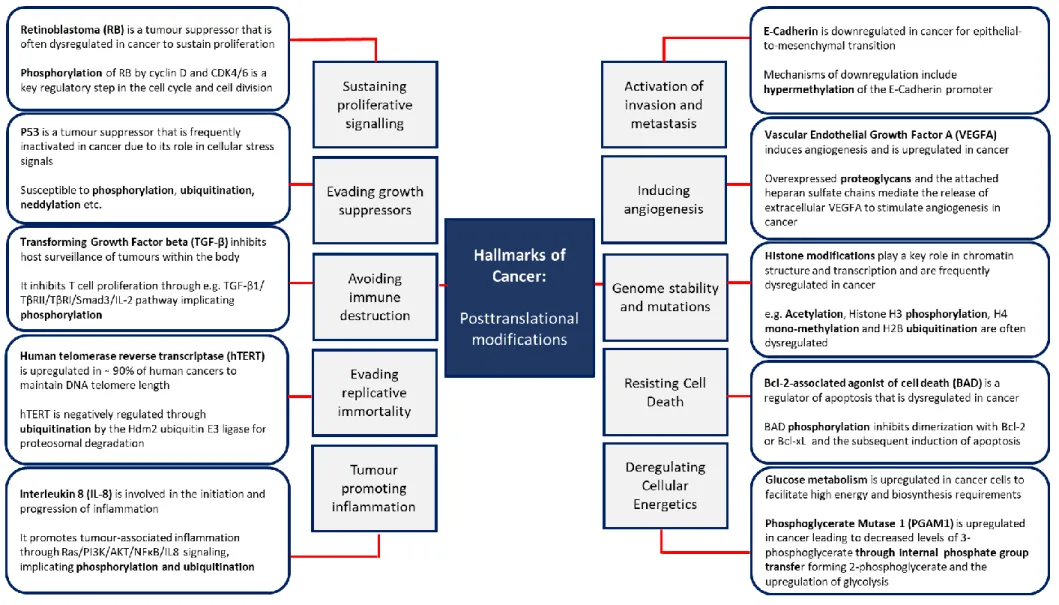
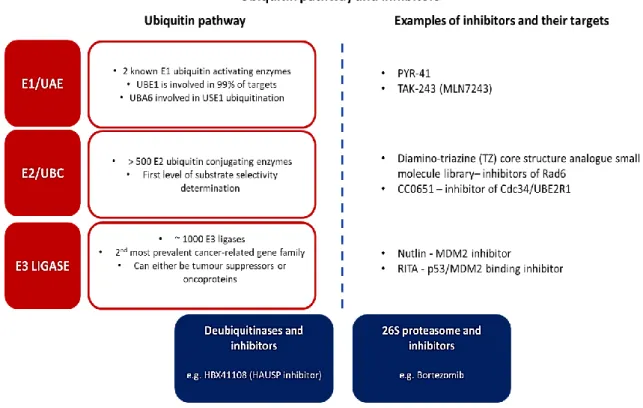
The ubiquitin pathway and related molecules
- Ubiquitination
- Ubiquitin and cancer
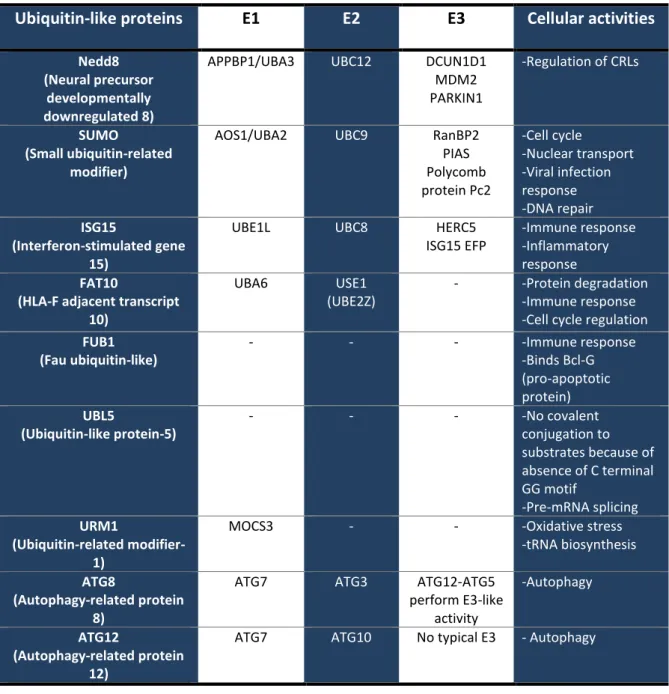
- Ubiquitin-like proteins
- Insights into ubiquitination using proteomics
This class of E3 ligases is the focus of this study because the cullin family of proteins undergo neddylation prior to conjugation to the other components of the CRLs (Kamura et al., 1999). Furthermore, deubiquitinases are also implicated in cancer due to their regulation of deubiquitination, including their role in substrate-specific deubiquitination (Wei et al., 2015; Singh and Singh, 2016).
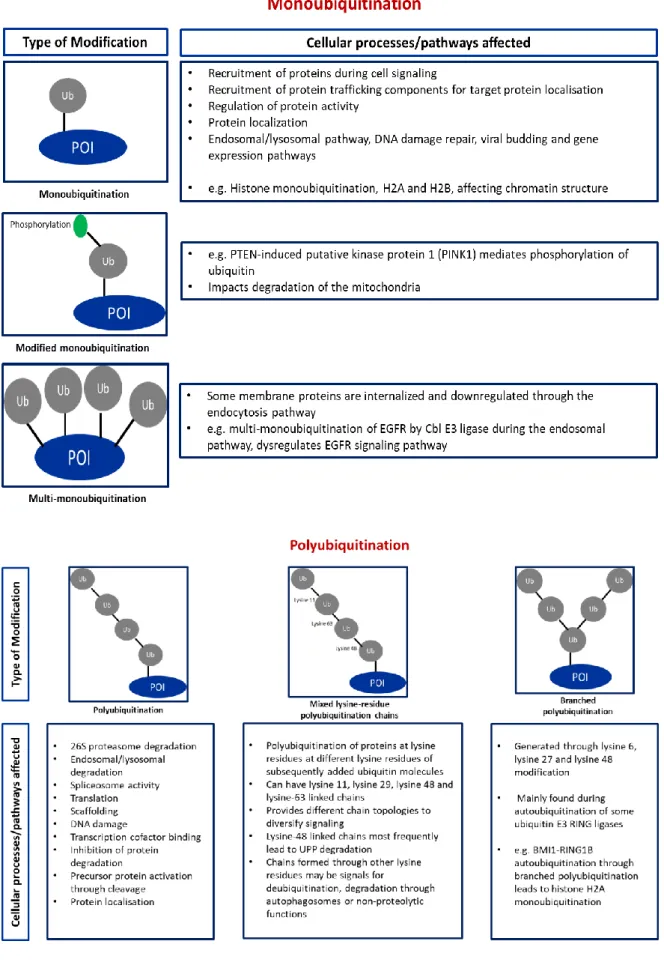
Neddylation
This would lead to the accumulation of proteins normally degraded by the 26S proteasome (Liao et al., 2011). Furthermore, using dual affinity purification (TAP) overexpression of labeled NEDD8 in Hela cells and the Q-Star Pulsar XL mass spectrometer, several ribosomal proteins were identified as targets for neddylation (Table 7) (Xirodimas et al., 2008).
DCUN1D1
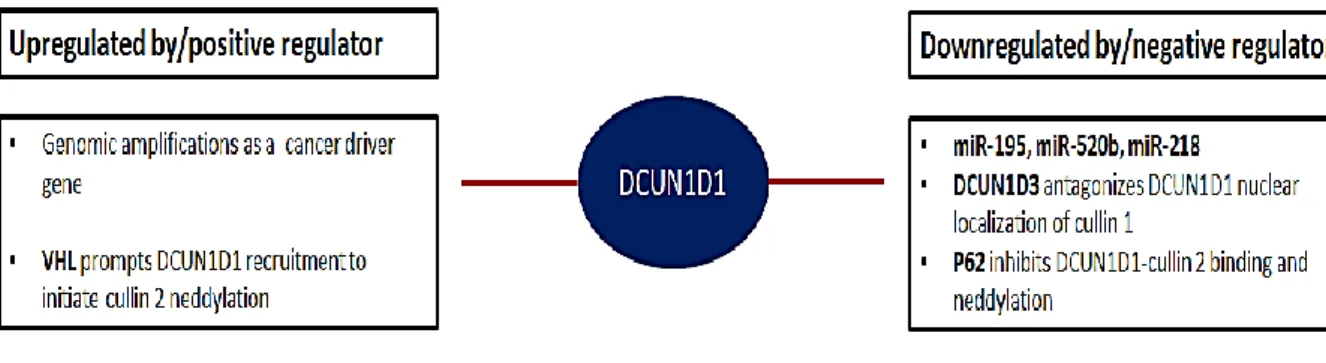
Furthermore, DCUN1D1 is inhibited by antagonism by DCUN1D3, p62 has been shown to inhibit DCUN1D1 binding to cullin 2 and VHL regulates the recruitment of DCUN1D1 to initiate cullin 2 nedylation in response to HIF1α binding (Figure 16) (Huang et al. This study aims to use different proteomics approaches to determine the mechanism of action of DCUN1D1 and to screen for potential inhibitors of DCUN1D1 using a proteomics approach.
Study Aims and Objectives
IPMS analysis of DCUN1D1 substrates
Introduction
Hypothesis
Aims and Objectives
Materials and Methods
- Cell culture and cell maintenance
- Calcium Phosphate Transfection
- Cell lysis and co-immunoprecipitation
- Tryptic digestion and C18 Stage-Tip clean-up
- LC-MS/MS
- Data processing and analysis
- Software/Tools used in the study
The co-precipitates then bind strongly to the cell monolayer and enable uptake of DNA by the cells (Wigler et al., 1979). They are then organized based on controlled vocabularies that generate standardized information for analysis by the user (Mellacheruvu et al., 2013).

Results
- Overall sample data analysis
- Filtered sample data findings to identify true interactors of DCUN1D1
Interestingly, Xirodimas et al., 2008 described several ribosomal proteins as neddylation substrates, but the neddylation E3 ligase responsible for this function was not identified. It also functions as an exchange factor in terms of the F-box binding proteins recruited for substrate recognition within the relevant CRL complex (Schmidt et al., 2009). Neddylation has also been shown to recruit histone deacetylases to repress NFκB-mediated gene expression ( Gao et al., 2006 ).
As previously described, histone molecules are components of the nucleosome complex consisting of 2 molecules each of H2A, H2B, H3 and H4 (Luger et al., 1997; Mariño-Ramírez et al., 2005).
Discussion
Interestingly, FUS has been shown to be an E3 ligase for the Erbb3 receptor-binding protein, Ebp1, and has been implicated in PCa progression, the regulation of AR and AR gene transcription, as well as PCa hormone resistance (Zhang et al. Gannon et al., 2008; Oh et al., 2010). Neddylation has previously been implicated in inflammatory responses through regulation of T cell activity via the ERK pathway, and Neddylation has been shown to regulate NF-κB expression ( Gao et al., 2006 ; Jin et al., 2013 ). This is most likely because among the cullin family of proteins, cullin 3 has been widely associated with viral/host interactions and the regulation of the endosomal pathway, while TRIM21 has been characterized as an Fc cytosolic receptor under viral entry and an E3 ligase that regulates innate and adaptive immunity (Yang et al., 2009;
Recently, the tumor microenvironment and its role in tumorigenesis has been established (Hanahan and Weinberg, 2011; Balkwill, Capasso, and Hagemann, 2012; Quail and Joyce, 2013; Belli et al., 2018).
SILAC for quantification of differentially expressed proteins
Introduction
This can be complicated by a variety of factors, including the fact that it requires the addition of peptide/protein at a known concentration that can be detected and identified during MS runs, bearing in mind that molecules respond differently to MS, variances due to sample preparation, and run works by MS (Lindemann et al., 2017; Ankney, Muneer, & Chen, 2018). However, these challenges can be mitigated by normalization using peptide, protein, or peptide concatemer standards, experimental design, and different algorithms (Cox et al., 2014; Ankney, Muneer, & Chen, 2018). In light of the above, we performed relative quantitative proteomics using SILAC to quantify proteins differentially expressed after DCUN1D1 knockdown in PCa cells.
Hypothesis
Aims and Objectives
Materials and Methods
- Cell seeding and SILAC incorporation
- Sample preparation
- Filter Aided Sample Preparation and Tryptic Digestion
- C18 Stage-Tip clean-up
- LC-MS/MS
- Data processing and analysis
- Software/Tools used in the study
Media for the “light” and “heavy” samples were prepared and then 1% penicillin (5000 µg/ml/streptomycin 5000 µg/ml) (Lonza, USA) was added prior to filter sterilization of the media using filters of 0. 22 µm. (Techno Plastic Products, Switzerland) (Figure 31). For the SILAC labeling uptake efficiency test, 100 µg of each lysate was prepared, while 50 µg of each "light" and "heavy" lysate were prepared. After incubation, the samples were centrifuged at 14,000 x g for 10 minutes, with the flow-through containing digested tryptic peptides.
We used Perseus version 1.6.5.0 to perform data filtration and statistical analysis of Maxquant output data (Tyanova et al., 2016).
Results
- SILAC labelling incorporation efficiency and mixing ratio analysis
- SILAC MS/MS data analysis
Analysis of duplicate samples resulted in the identification of 1443 unique peptide amino acid sequences from 3743 identified spectra. CCT3 and TCP1 are components of the protein folding complex called the chaperone-containing T-complex, with TCP1 involved in the folding of 10% of the cellular proteome ( Leitner et al., 2012 ). We found RPS10 which is a component of the 40S ribosomal subunit and the B subunit of eukaryotic translation initiation factor 3 (EIF3B).
The proteins involved in processing in the ER are shown in the upper right corner of the associations.
Discussion
Interestingly, a number of "Cancer Safeguards" were implicated in pathways such as proliferative signaling e.g. Regarding the list of significantly dysregulated proteins after knockdown of DCUN1D1 in DU145 PCa cell lines, we found 33 proteins in the LH2 sample with 14 down-regulated and 19 up-regulated. Identification of dysregulation of some of the 40S small ribosome subunits and eukaryotic initiation factors.
TCP1 has been demonstrated to be involved in the folding of 10% of the cell proteome and found to be upregulated in prostate tissue samples using tissue microarray analysis (Leitner et al., 2012).
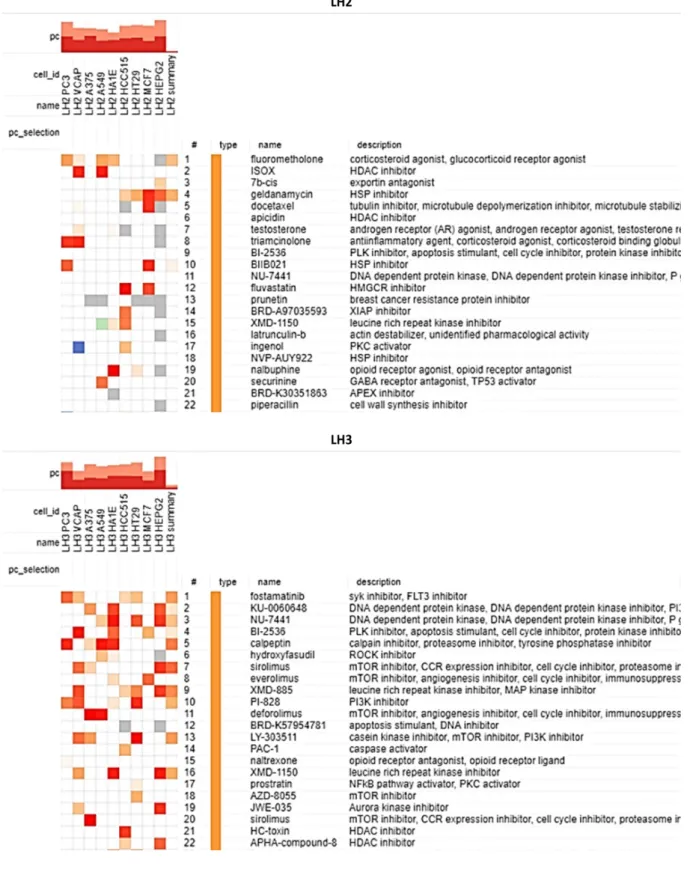
Advanced Connectivity Map analysis for identifying DCUN1D1 inhibitors using
- Introduction
- Hypothesis
- Aims and Objectives
- Materials and Methods
- Results
- Discussion
Significantly, of the 22 main compounds observed, we found the following compounds in common among the samples: BI-2536 (PLK inhibitor/apoptosis stimulator/cell cycle inhibitor), NU-7441 (DNA-dependent protein kinase inhibitor ) and XMD-1150 (leucine-rich repeat kinase inhibitor). Heatmaps showing the top 9 cell lines of the reference data set and the ranked list of compounds for sample LH2 (top) and sample LH3 (bottom). Heatmaps showing the top 9 cell lines of the reference data set and the ranked list of compounds in sample LH2 (left) and sample LH3 (right).
Analysis of the results in PCa cell lines found talniflumate, atorvastatin, GR-135531, W-9, RX-821002 and palmitoylethanolamide shared in the PC3 output.
Preferential cullin neddylation and dysregulation of the WNT pathway by
- Introduction
- Hypothesis
- Aims and Objectives
- Materials and methods
- Cell lines and culturing
- Protein quantification and western blot analysis
- Results
- DCUN1D1 knockdown in PCa cells decreased global neddylation, ubiquitination
- DCUN1D1 shows preferential neddylation activity of cullin proteins in PCa
- The WNT pathway was inhibited following knockdown of DCUN1D1 in PCa
- Blockage of DCUN1D1 activated the AKT pathway in PCa
- Discussion
It also suggests that dysregulation of the neddylation pathway was key to the mechanism of action of DCUN1D1 in PCa. Western blot showing decreased expression of the cullin-related proteins RBX1 (upper panel) and CAND1 (lower panel). Therefore, the blockade of DCUN1D1 resulted in the deactivation of the WNT pathway, confirming its role in the mechanism of action of DCUN1D1.
Therefore, the activation of the AKT pathway may be a survival response to the inhibitory effects of DCUN1D1 knockdown in PCa.
General Discussion, Conclusion and Future Work
General Discussion
Interestingly, blockade of DCUN1D1 in PCa cells produced a signature strongly associated with kinase inhibitors. This would be the first description of DCUN1D1-mediated preferential neddylation of cullin proteins in PCa. These activities overlap with the observations we made on DCUN1D1 and imply this in the mechanism of action of DCUN1D1.
According to the strength of the evidence obtained in our laboratory at the level of transcriptomics and proteomics, we demonstrated using western blot that the WNT/β-catenin pathway plays a role in the mechanism of action of DCUN1D1 in PCa.
Conclusion
Analysis of DCUN1D1 binding partners identified cullin proteins as target proteins of DCUN1D1, which together with previously published studies on DCUN1D1, we hypothesize may be the primary targets of DCUN1D1. We also identified neddylation substrates such as ribosomal proteins with which DCUN1D1 is associated. However, it was not clear whether this could be an E3 ligase or part of a broader mechanism, such as through its implication in eukaryotic translation or dysregulation of ribosome biogenesis in PCa. Analysis of the drugs strongly associated with the DCUN1D1 knockdown signature in PCa revealed aspects of the involved pathways as well as strong connections to kinase inhibitors, raising interesting questions around the role of DCUN1D1-mediated kinase signaling pathways in PCa.
We validated the role of cullin 3, cullin 4B and cullin 5 down-dylation in the mechanism of DCUN1D1 in PCa, showing preferential decrease in cullin down-dylation of cullin 1, 3, 4A, 4B and cullin 5.
Future Work
1992) 'Prostate specific antigen density: a way to distinguish benign prostatic hypertrophy from prostate cancer.', The Journal of Urology, 147(3), pp. 2014) 'New insights into the ubiquitin E3 ligase mechanism', Nature Structural & Molecular Biology. Measurement of Prostate-Specific Antigen in Serum as a Screening Test for Prostate Cancer.”, The New England Journal of Medicine, 324(17), pp. prostate-specific angen to improve prostate cancer differentiation from benign prostate disease:. 1999) “Local Control of Prostate Cancer By Transrectal High Intensity Focused Ultrasound Therapy: Preliminary Results.”, The Journal of Urology, 161(1), pp. 2008) ‘The Atg8 and Atg12 ubiquitin-like conjugation systems in .
Pacific Symposium on Biocomputing, 123(2), s. 2014) 'RNF168-medieret H2A-neddylering antagoniserer ubiquitylering af H2A og regulerer reparation af DNA-skader', Journal of Cell Science, 127(10), s. Oncology: Official Journal of the American Society of Clinical Oncology, 17(3), s. 2012) 'The TMPRSS2: ERG Rearrangement, ERG Expression, and Prostate Cancer Outcomes: A Cohort Study and Meta-analyse.', Cancer. 2007) 'Cullin4B / E3-ubiquitin ligase regulerer negativt β-catenin', Journal of Biosciences, 32(september), s. 2012) 'Paradoxical Roles of Tumor Necrosis Factor-Alpha in Prostate Cancer Biology', Prostate Cancer, s. 2012. .