These would both contribute greatly to our understanding of floral specialization and the mechanisms involved in angiosperm radiation. Specialized pollination by large spider-hunting wasps and self-incompatibility in the African milkweed Pachycarpus asperifolius. Despite its widespread use, the traditional concept of pollination syndromes has recently been criticized (Ollerton 1996; Herrera 1996; Waser et al. 1996), resulting in rigorous discussions of floral specialization in the literature (Johnson & Steiner 2000;).
The research of Waser et al. 1996) led to renewed interest in the levels of specialization in plant pollination systems, and a number of researchers began to critically examine the applicability of pollination syndromes as a conceptual framework for floral evolution. In the Iridaceae, two South African species of Ferraria were recently found to be pollinated exclusively by wasp-like (eumenine and masarine) wasps (Goldblatt et al. 2009). Until recently, specialized pollination systems powered by spider-hunting wasps (Pompilidae) were completely unknown among angiosperms.
Specialized pollination by Hemipepsis wasps on rewarding plants was only recently rediscovered when a study by Ollerton et al. 2003) in the middle zone of KwaZulu-Natal found that three milkweeds, Miraglossum pilosum, M. The pollination niche and its role in the diversification and maintenance of the South African flora.
Palp-Faction: An African Milkweed Dismembers Its Wasp Pollinators
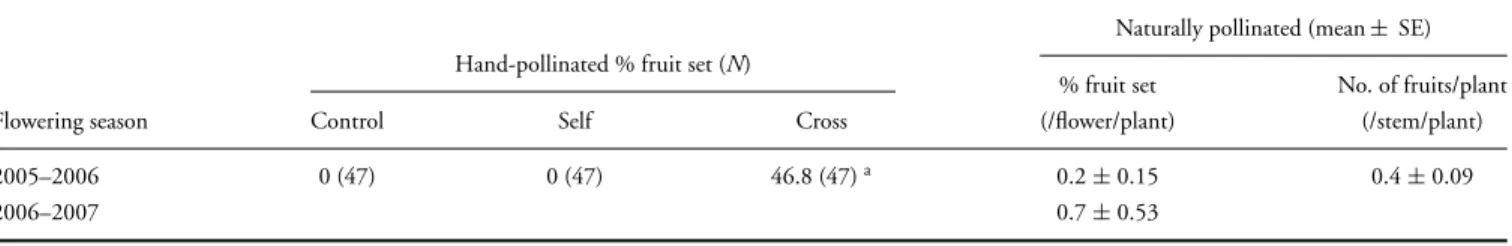
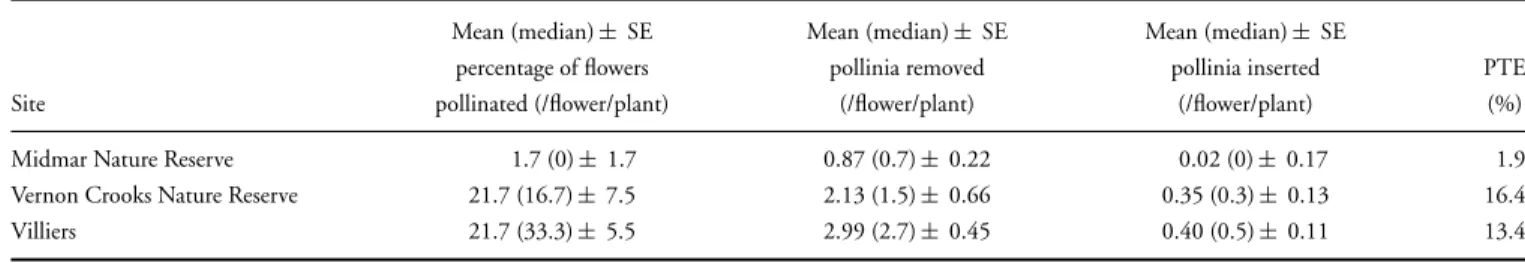
Pollinator Effectiveness: Cage Experiments. In the cage experiment, 40 pollinia (in 20 pollinaria) were removed and 17 pollinia were introduced (see Table 2 for means per ßfunction). These plants were tagged and the number of flowers on each plant was recorded at the beginning of the flow- Chapter 3. Voucher specimens of the studied species are deposited in the NU Herbarium (University of KwaZulu-Natal, Pietermaritz- burg).
A revised classification of the Apocynaceae s.l. Diversity and temporal change in the effective pollinators of Asclepias tuberosa. 1979. The genus Hymenoptera described by Linnaeus in the genera Sphex, Chrysis, Vespa, Apisand Mutilla. Biological Journal of the Linnean Society 12: 45 – 84.
SUPPLEMENTARY DATA
Compounds are listed in order of increasing retention time (using an Alltech EC-WAX column) within each compound class. Mass fragments for unknowns are listed with the molecular ion marked * first (if known), followed by the base peak and other fragments in decreasing order of abundance.
The importance of scent and nectar filters in a specialized wasp-pollination system
INTRODUCTION
Pollinator‐mediated selection on floral traits typically arises from the morphology, behavior and sensory modalities of particular pollinator species (Harder & Johnson 2009 ). Evolutionary shifts among pollinators can therefore diversify flower forms and result in reproductive isolation (Grant 1949; Stebbins 1970; Johnson 2006). Understanding the mechanisms responsible for hastening a shift from one pollen vector to another is thus of particular importance, as this can often represent the first stage of plant speciation.
To this end, a number of studies have investigated the functional importance of floral morphology or color for pollinator attraction in closely related species (Bradshaw & Schemske 2003; Irwin &. The adaptive importance of floral scent has also been suggested by studies demonstrating links between Scent composition and different types of pollinators (e.g. Knudsen & Tollsten. Chen et al. 2009; Shuttleworth & Johnson 2009a,b,c) suggest that differences in the production of specific floral volatiles in some systems , can change their attractiveness and initiate shifts between different types of pollinators (Kessler et al. 2008).
Groups of closely related species that show differences in pollinators and floral scent, rather than morphology or color, are ideal for investigating the role of scent evolution in pollinator transitions. Hyacinthaceae) represents a promising study system, as the ten species have morphologically unspecialized flowers with exposed nectar and yet apparently have highly specialized and diverse pollination systems (Shuttleworth & Johnson 2009a; this study). In a previous study, we showed that pollinator attraction in the specialized pompilid-wasp pollination systems of Eucomis Autumnalis and E.
Preliminary observations of several closely related Eucomis species (E. bicolor, E. humilis, E. schijffii, E. montanan and E. vandermerwei) indicated that they differ dramatically from the wasp-pollinated species in scent chemistry and are pollinated by scavengers. We thus hypothesized that the divergent pollination systems in Eucomis are rather mediated by differences in floral scent chemistry. The objectives of this study were (i) to determine whether some Eucomys species depend on scavengers for pollination, (ii) to compare floral scent, color, morphology and nectar traits of these fly-pollinated species with wasp-pollinated Eucomys species to determine which traits are associated with divergent evolution, and (iii) to determine whether a pollinator shift can be induced by experimental manipulation of floral scent.
MATERIAL AND METHODS
The quantification was based on 68 synthetic standards that were injected and thermally desorbed under identical conditions to the samples (Shuttleworth & Johnson 2009a). To determine whether differences in floral scents are related to pollination system, we plotted scent data for the four Eucomis species (data for the wasp-pollinated species are presented in Shuttleworth & Johnson (2009a)) in two dimensions with NMDS using Primer Clarke. & Warwick 2001; Clarke & Gorley 2006). Differences in the odor profiles between species and pollination systems were tested using ANOSIM (Clarke & Gorley 2006), a nonparametric permutation procedure based on the similarity matrix underlying the ordination (Clarke & Warwick 2001).
Observed R's were compared to the distribution of R generated by up to 10,000 random permutations of the sample labels to. Volatiles characterizing the scent of each species and each pollination system were examined using the "similarity percentages" (SIMPER) function of Primer (Clarke & Gorley 2006). SIMPER identifies compounds that contribute most to the average similarity within a particular group (a species or pollination system).
To determine the effects of sulfur compound production by non-fly-pollinated Eucomis flowers on pollinator preference, we added a 1:1 mixture of dimethyl disulfide (DMDS) and dimethyl trisulfide (DMTS) to bee-pollinated inflorescences. Eucomis species and then recorded the behavior of flower visitors. A small vial (10 ml) with a cotton wick and containing either 100 ml DMDS, 100 ml DMTS and 800 ml white mineral oil (to slow evaporation) or 1000 ml white mineral oil alone (for control) was attached then at the base of each. inflorescence and hidden from view by leaves. Experimental inflorescences were placed about 1 m apart and insects visiting the flowers in each inflorescence were observed.
The absence of sulfur compounds in the natural odors of the two inflorescences was confirmed by GC–MS after the experiments were completed (sufficient time was given for any remaining sulfur compounds to evaporate prior to analysis). Flowers on both experimental and control inflorescences were emasculated to avoid contamination of gene pools. The inflorescences were moved to new positions three times during the 3.5 h of this experiment, but the control and scent treatments were not switched between plants.
The sulfur and control bottle were switched once between blooms and the bloom was moved once to new positions during the 2 h of this experiment.

RESULTS
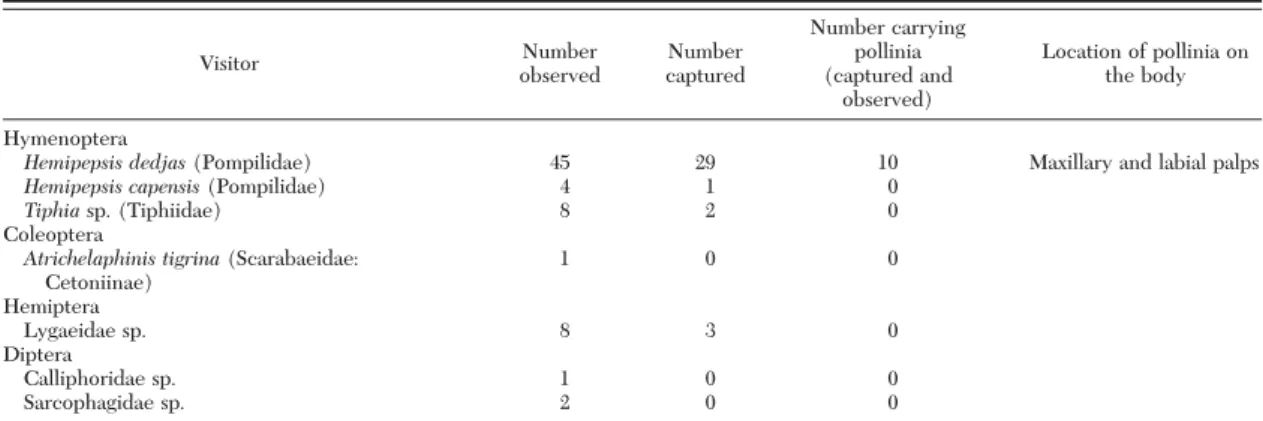
In biological experiments, inflorescences of species normally pollinated by wasps with experimentally added sulfur compounds attracted large numbers of flies (mostly calliphorids and sarcophagids) that would not normally visit these flowers (binomial test comparing fly visits to experimental and control inflorescences, p .0.001 for both Eucomis species; figure 4 and electronic supplementary material, table S4). Flies that landed on the experimental flowers behaved differently: some settled on the inflorescences and then periodically explored the flowers and drank nectar, while others (especially Sarcopha. Very few flies crawled to the base of the inflorescence to explore vials from which the sulfides came). .
DISCUSSION
Flower colors of the four Eucomis species in the color space of bees and flies. a) Loci in the bee-colored hexagon (in this case used as a model for wasp vision; Chittka 1992). The distance between loci in the hexagon correlates with the distinguishability of the colors by bees (or in this case wasps). Specialization in the wasp-pollinated species Eucomis is based on floral scent (Shuttleworth &. Johnson 2009a), and this also appears to be the case in the fly-pollinated species.
2009cSpecialized pollination in the African milkweed Xysmalobium orbiculare: a key role for floral scent in the attraction of spider-hunting wasps. The distribution of the guild is centered in the moist highland grasslands of eastern South Africa. Non-metric multidimensional scaling (NMDS) of scent data for 15 guild members and 16 congeners (obtained through GC-MS analysis of headspace samples and supplemented with published data) provided little evidence for convergent evolution in the overall scent composition of guild members.
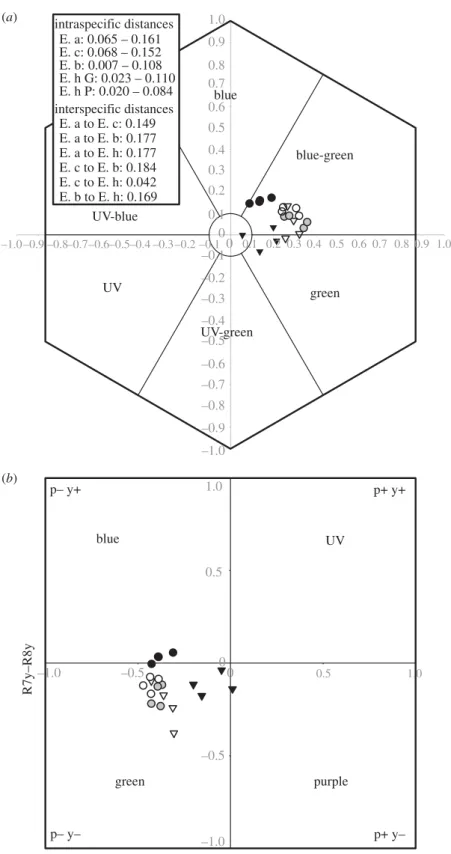
Convergence in floral spectral reflectance was evident in guild members; in particular, spectra of guild members clustered in the blue to blue-green region of the hymenopteran color hexagon and were significantly closer to the color of background vegetation than those of congeneric species. Non-metric multidimensional scaling (NMDS) analysis of the floral scent profiles of guild members and congeneric (non-wasp-pollinated) species revealed little evidence for overall convergence in floral scents of wasp-pollinated species ( Fig. 3 ). Loci for these spectra cluster close to the background vegetation in the green to blue-green region of the color hexagon (Fig. 5).
The distribution of the guild is clearly centered in the moist grasslands of eastern South Africa (Fig. 2). However, the flower colors of the guild members clustered close to the background in the blue to blue-green range of the color hexagon and were significantly closer to the color of the background vegetation than they were in congeneric species (Fig. 5). The lack of convergence in guild members' floral scent profiles (Fig. 3) is intriguing.
Cryptic flower coloration in the guild is confirmed by the close proximity of guild member loci to background vegetation in the color hexagon (Fig. 5). Indeed, the members of the guild are all inconspicuous in the landscape, and many of the smaller asclepiads (such as Aspidiglossum glanduliferum, Periglossum angustifolium and Miraglossum species) are best found by following foraging wasps, as first noted by Weale (1873). Specialized pollination in the African milkweed Xysmalobium orbiculare: a key role for floral scent in the attraction of spider-hunting wasps.