Differences between inoculated and uninoculated plants of Acacia nilotica grown under both ambient and high CO2 conditions. Visual differences between Acacia nilotica and Acacia sieberana plants grown under elevated CO2 conditions when inoculated or uninoculated.
Introduction and Literature Review
Rising carbon dioxide levels
An area of particular concern is the combined effects of rising CO2 levels and temperature on the productivity (and thus nutrient status) of native plant communities. Various authors have found a range of combined effects of elevated CO2 and elevated temperature (RAWSON found a less than additive effect, indicating a decline in the response to elevated CO2 with increasing temperature.
The effects of elevated CO 2 on plants
The effect of elevated CO2 on transpiration ranges from negligible to nearly a 70% reduction, depending on the species (BREMER et al. 1996). For example, if Rubisco efficiency is higher under elevated CO2, less N would be required per unit dry matter gain (NORBY et al. 1986).
Ecosystem responses to elevated CO 2
It is therefore not obvious how increased CO2 and climate change will affect the quality and quantity of soil organic matter, or the degree of nutrient limitation (HUNT et al. 1996). There is limited evidence that increasing CO2 will increase osmotic adaptation and thereby increase the tolerance of some plants to dehydration (POLLEY et al.1996).
Leguminous trees and nitrogen fixation
The degree of competition for soil available nitrogen by associated non-legume crops is one factor that determines the proportion of nitrogen obtained from nitrogen fixation (HARTWIG et al. 1996). Seasonal changes in nodulation and nodule specific activity (lower in the dry season) produce marked variation in fixation rates (LANGKAMP et al. 1982; BARNET et al. 1985).
The effects of elevated temperature and nitrogen on plant growth
CO2 enrichment can moderate adverse effects of temperature extremes (ROGERS and DAHLMAN 1993; BOWES et al. 1996). Elevated CO2 may allow plants to survive at lower nutrient levels, but the plant growth response to increased nutrient availability is greater under elevated CO2 (BAZZAZ 1990; WONG et al. 1992).
Competition between C 3 and C 4 plants in elevated CO 2
However, C4 plants may be favored in warmer, drier environments (COLEMAN and BAZZAZ 1992; CAMPBELL and HART 1996) independently of CO2. From this theory, it is predicted that plants from productive habitats will be the fastest to increase competitive effects in response to increased resource availability, i.e.
The effects of elevated CO 2 on other biological processes
Herbivores may be adversely affected if biomass production remains unchanged and nitrogen percentages decrease under elevated CO2 conditions. WILSEY (1996) found no large increase in aboveground biomass in response to elevated CO2, ARNONE et al. 1995) found no major differences in leaf nutrient quality between elevated and external CO2 treatments.
Plant acclimation to elevated CO 2 levels
SCHAFFER et al.(1996) found that CO2 enrichment may have compensated for the effects of root restriction on plant growth, as there was no effect of root chamber size on plant dry weight at elevated CO2. BERNSTON et al. (1993, cited in ARNONE 1996) showed that increasing the nutrient additions can eliminate the effects of small pot size.
Photosynthetic Response Curves (or A/C I)
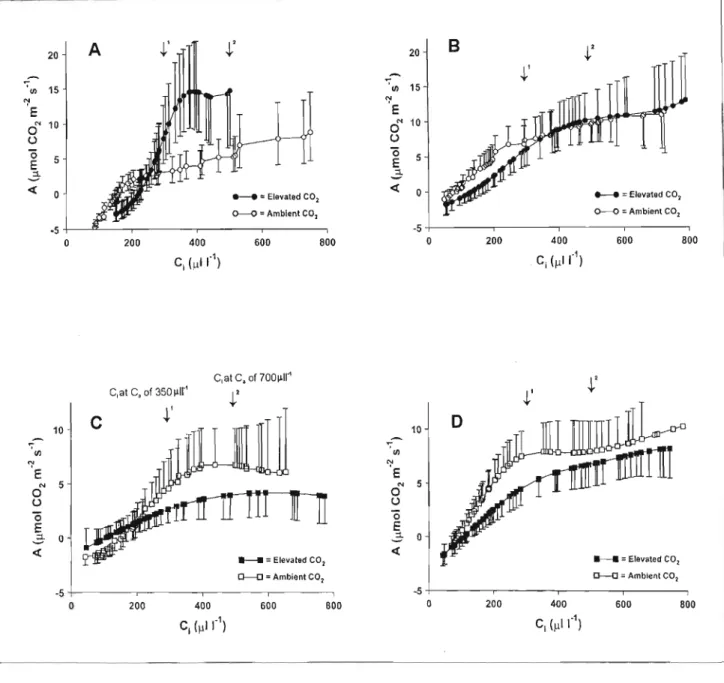
2, the ratio of C, toC, at C, = 350 I-lW1 was slightly lower for plants grown in ambient compared to plants grown under elevated CO2, although stomatal conductance (gs) did not differ significantly. SAGE (1994) also reported on a number of studies in field-grown plants since 1986 in which long-term exposure to CO2 produced little change in the net CO2 assimilation response, indicating a lack of adaptation to elevated CO2 in natural environments. The best way to determine the effects of elevated CO2 using A1Cj data would involve determining the ratio of A from plants grown in elevated C to plants grown in normal C, when both are measured at the same C , if photosynthetic adaptation to elevated CO2 has occurred, this assimilation ratio will vary from one over a wide range of C·I' values, a value greater than one indicating greater CO2 assimilation under elevated CO2 and a ratio less than one indicates downregulation under elevated CO2.
Stomatal Conductance
See earlier Competition between C3 and C4 plants in elevated CO2), However, it has been observed that the CO2 photosynthetic response curves of various C4 species show that photosynthesis is not always saturated at an environmental concentration of 600 1J1I-1 (POORTER 1993). It appears that in some species, the C4 pathway is not as tightly controlled as previously suggested, and therefore allows a response to CO2 (POORTER 1993).
Possible effects of global warming in KwaZulu-Natal
If such a plant were fast growing, would be useful for fuelwood, would improve nitrogen availability in the soil and would improve the nutritional status and condition of the soil (ALEXANDER 1989; BELSKYet al. 1989; TIEDEMANN and KLEMMEDSON 1973;TIEDEMANN and KLEMMEDSON 1986) and would also be useful in reducing soil erosion, this would be very coincidental. The improved condition of the soil under these trees would also improve the growth and quality of the grass (whether it is a C3 grass in the shaded conditions with elevated CO2, or a C4 grass at elevated temperatures). Temperature influences plant growth by affecting the biochemical processes that control resource acquisition, growth, and energy budget.
Objectives and Introduction to the Present Investigation
The direct assessment of the impact of increasing CO2 is very difficult due to the long-term nature of CO2 effects and the countless potential interactions between CO2 and other environmental factors that can affect physiological and ecological contexts (NORBY 1996). The best approach is to use FACE or even larger enclosures, but these are very expensive to set up and expensive to operate due to the sophisticated equipment involved (TIBBITIS and LANGHANS 1993) and the large amounts of CO2 emitted each day.
Plant growth in response to CO 2 enrichment with two different Rhizobium inoculation treatments
Introduction
Therefore, the aim of this study was to determine the effect of elevated CO2 on plant growth in two Acacia species. These variables were expected to give some idea of the relative success of these two species under a future elevated CO2 regime.
Materials and Methods
The time it takes a given bubble to travel a certain distance gives an indication of the CO2 flow rate. Plants were inoculated by watering them with an infusion of the soil from under A. This resulted in the closure of some of the leaves, reducing the leaf area in some cases.
Results
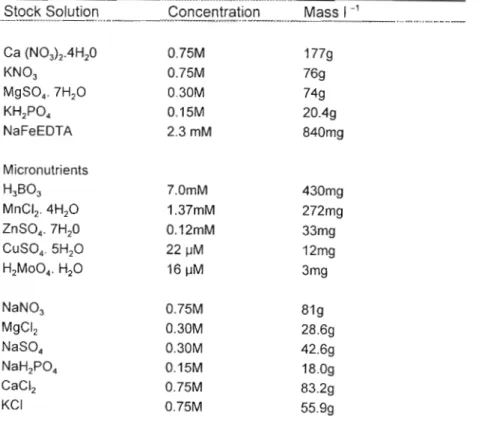
Final plant heights in both species were greater in plants grown in elevated CO2 and with rhizobia. Visual differences between Acacia nilotica and Acacia sieberana grown under ambient conditions, either inoculated or uninoculated with rhizobium root nodule bacteria. The differences between Acacia ni/otica inoculated and uninoculated plants grown under both ambient and elevated CO2.
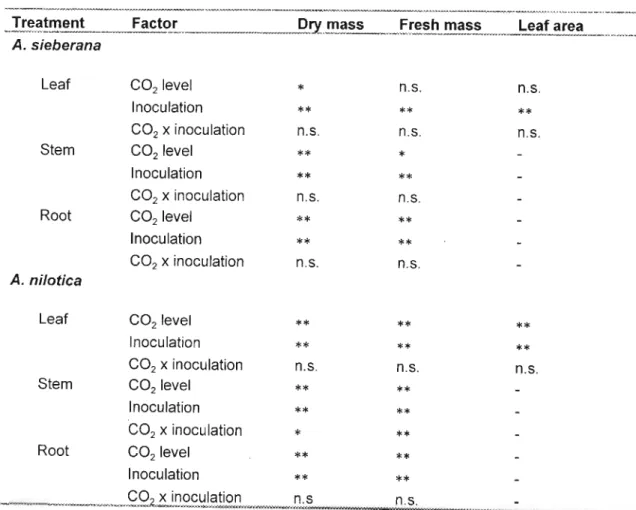
Discussion
Root-shoot ratios were greatest in plants grown without rhizobial inoculation in both CO2 treatments and were on average higher in ambient CO2 treatments than elevated CO2 treatments for both species (Table 2.3). eel. 1996). This can alter the competitive balance of species by modifying leaf light capture patterns (BAZZAZ and MCCONNAUGHY 1992; Luoand MOONEY 1995), which may be particularly important during seedling establishment (WAND et al. 1996). However, elevated CO2 can also improve plant water status and thereby affect RSR (HUNT et al. 1996).
Conclusions
This is because there is a lack of understanding of the mechanisms that control the overall response of plants to CO2 concentration (MITCHELL et al.). This suggests that when species are differentially infected, nitrogen-fixing capacity will be important in understanding shifts in species composition in response to increased CO2 emissions, even when soil fertility is too low to support a community-level response (REYNOLDS 1996 ). It is clear that nitrogen-fixing species will be favored in a CO2-rich world, and the results obtained in this study confirm this assumption.
Introduction
Materials and Methods
Results
Analysis of variability (ANOVA) in stomatal conductance revealed only one obvious significant difference between the two species (Table 3.1). Only the results of the uninoculated plants grown in elevated CO2 seemed to differ from the other treatments. This may have been an artifact of the experimental procedure used to determine stomatal conductance as this treatment had very variable results.
Discussion
In this study, photosynthesis was reduced in plants grown without rhizobial inoculation (low-nitrogen related) in both ambient and elevated CO2, except for the uninoculated A.nilotica treatment (Figure 3.2). In this case, the plants grown under elevated CO2 (uninoculated) had significantly lower stomatal conductance than all the other plants/treatments. Reductions in stomatal conductance and the associated increase in A due to CO2 enrichment (the exact mechanism is unclear) are the main factors that increase WUE at elevated C, (EAMUS and JARVIS 1989; . POORTER 1993; SAGE 1994).
Conclusions
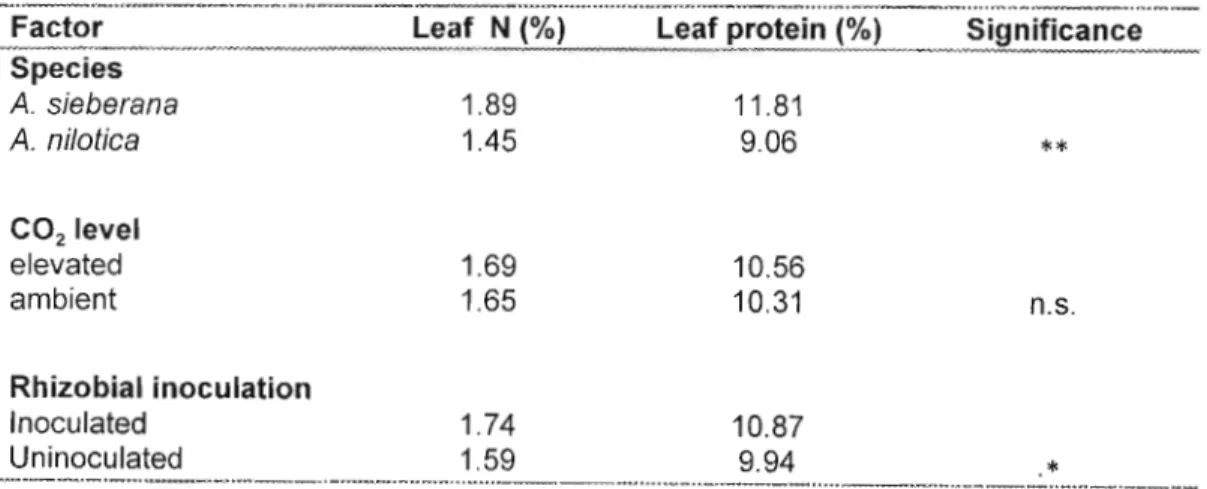
Thus, while photosynthesis was unexpectedly reduced, it was offset by some other mechanism to result in a significant increase in plant mass under elevated CO2 after termination of the experiment (Chapter 2). A simple explanation for the increase in growth (but not photosynthesis) could be that dark respiration was less in plants grown at elevated CO2 - for reasons that remain unclear. The effects of elevated CO2 and rhizobial inoculation on Acacia leaf nitrogen concentration and possible implications.
The effects of elevated CO 2 and rhizobial inoculation on Acacia leaf nitrogen concentration and possible implications
- Introduction
- Materials and Methods
- Results
- Discussion
- Conclusions
Therefore, the distribution and presence of root tuber bacteria may play an important role in the well-being and efficiency of CO2 utilization in elevated CO2. Increased soil exploration (DAY et al. 1996) and nitrogen uptake are the main mechanisms by which seedlings can respond to elevated CO2 under nitrogen/nutrient limitations. The results of this study, however, were contrary to expectations, as plants grown in an elevated CO2 environment did not have significantly lower leaf nitrogen percentages compared to plants grown in a CO environment.
- Introduction
- Materials and Methods
- Results
- Discussion
- Conclusions
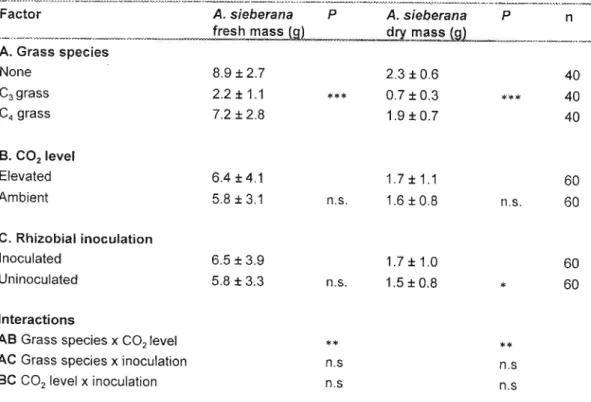
When grown with the C4 grass, the presence of both factors (increased CO2 and rhizobial inoculation) resulted in the greatest Acacia mass, and the absence of both, the lowest Acacia mass (Table 5.5). The C3 grass accumulated significantly more mass than the C3Acacia, although this mass decreased in the presence of the tree seedling and increased with increased CO2 and inoculation. It can be expected that the C3 grass will also do better than the C4 grass in the shade of the Acacia, a specific habitat adaptation.
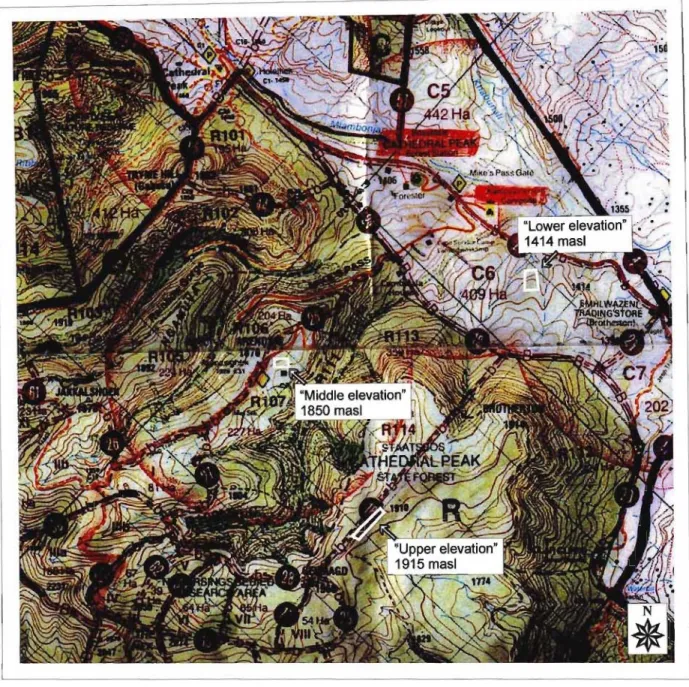
The effect on growth and survival of transplant seedlings of Acacia nilotica and Acacia sieberana at three different
- Introduction
- Materials and Methods
- Results
- Discussion
- Conclusions
Acacia sieberana grown at different heights showed no significant differences between the treatments (Table 6.1). The height of A.nilotica had increased significantly more by. At the end of the experimental period, the mass of the plants at the lowest altitude (1414 meters above sea level) was greatest in both species (P < 0.001). Baboons may have interfered with the plants at the higher elevation and damaged some of them.
NIJS, I.& IMPENS, I.(1996) Effects of increased CO2 concentration and climate warming on photosynthesis during winter in Lolium perenne. 1996) Effects of atmospheric CO2 enrichment and root limitation on leaf gas exchange and growth of banana (Musa). Plant water relations and the effects of elevated CO2: a review and suggestions for future research.