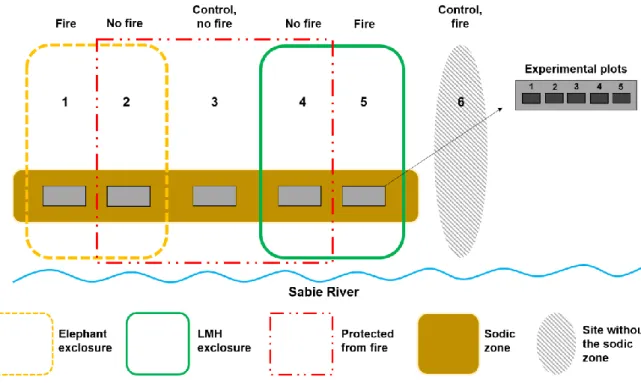
prof. William Bond for his valuable comments on the application of the Tea Bag Index, as well as Prof. Sally Archibald for her insightful input on nutrient cycling in grassy biomes; Appendix A: Supplementary Figure to Support Chapter 4: Experimental Layout of the Nkuhlu Enclosures with Fire and Without Fire Treatments.
List of symbols
Mean number of fungus-infested (a) and termite-damaged (b) tea bags in the three herbivore treatments. Whiskers show the standard error of the mean based on results obtained from 250 tea bags (125 green and 125 rooibos) over a three-month incubation period.
List of tables
Introduction
- Preface
- Rationale
- Format of study
Using the internationally standardized TBI approach as described by Keuskamp et al. 2013) has been tested and evaluated as a viable means of quantifying degradation in an African context. This unit has been accepted and published in the South African Journal of Science (Erasmus et al., 2021; Appendix D).
Literature review
- Semi-arid savannas
- Herbivory as a driver of savanna dynamics
- Conclusions
Available soil C is a major determinant of microbial biomass and activity (Bardgett et al., 1998; . Eldridge et al., 2019). Tea bags are subsequently able to act as suitable proxies for natural waste (Keuskamp et al., 2013).
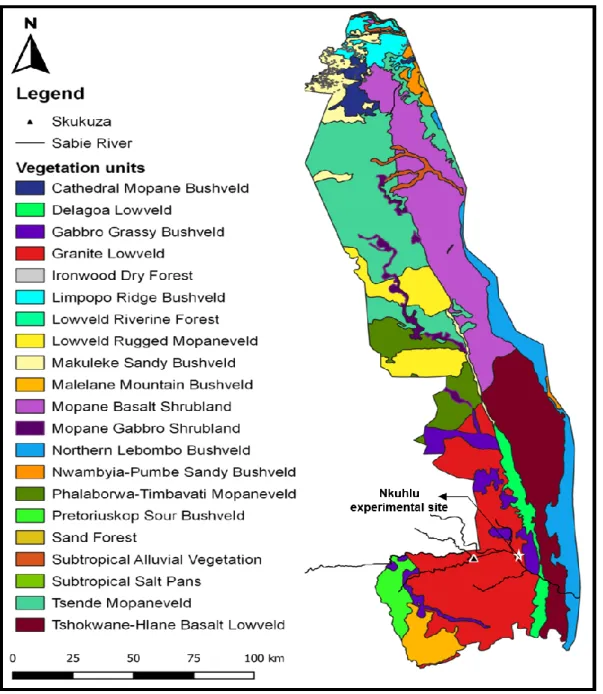
Study Area
- Locality and description
- Geology
- Topography
- Soil
- Introduction
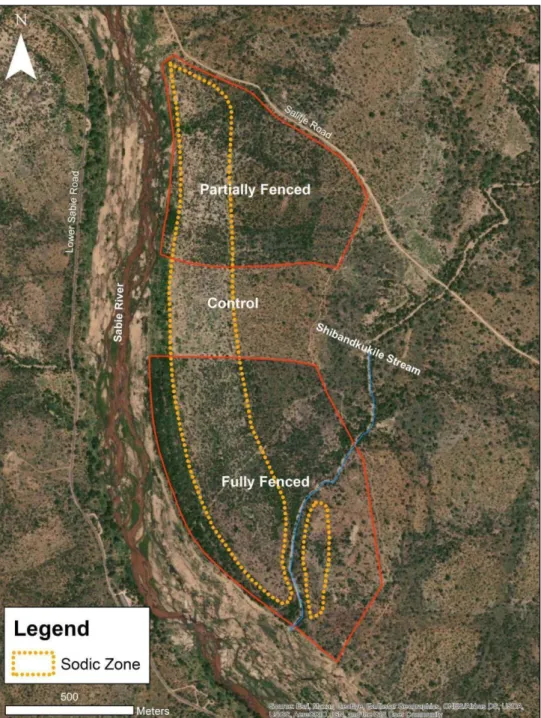
- Experimental design .1 Nkuhlu experimental site
- Data analyses
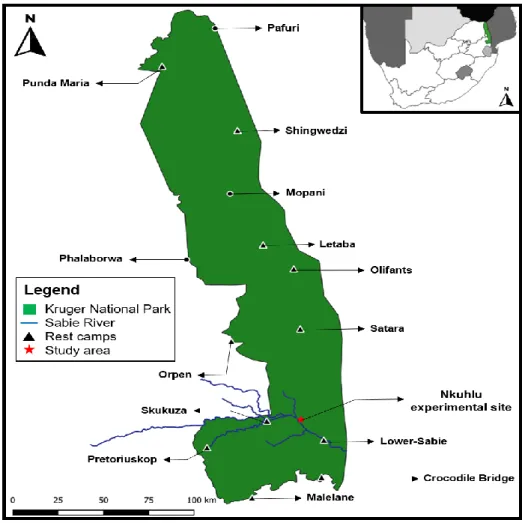
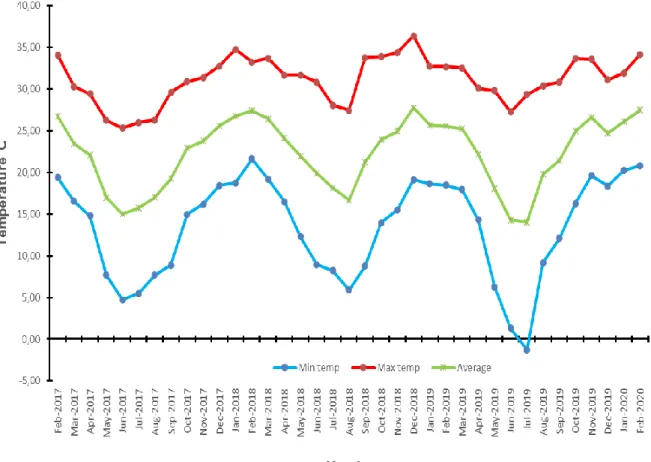
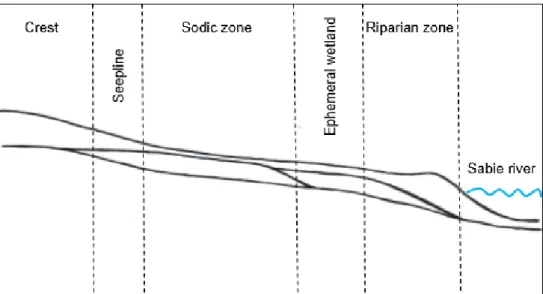
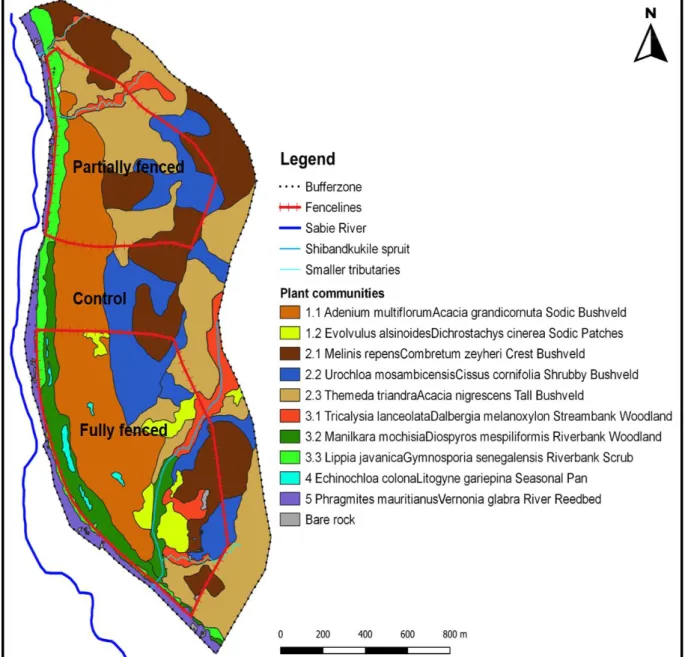
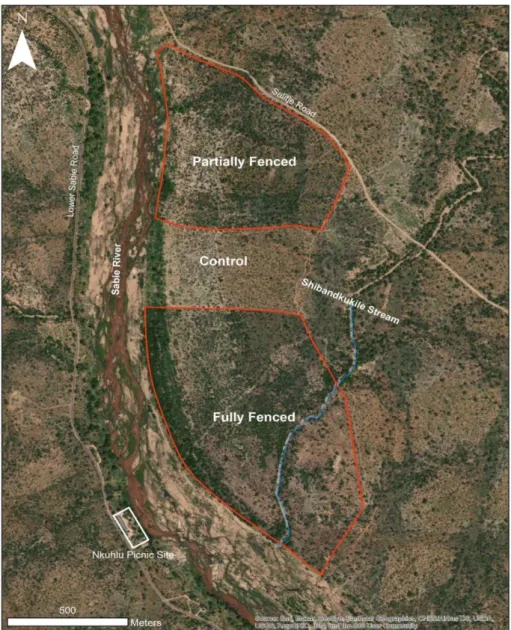
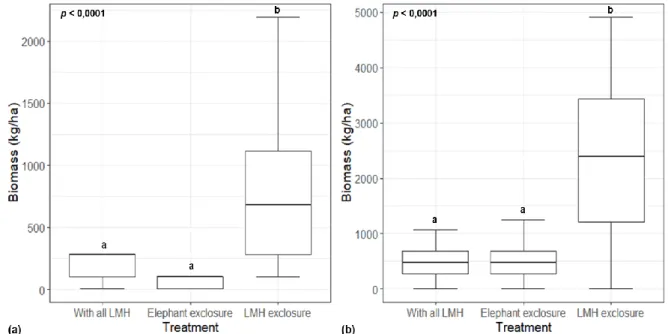
The study site, located within vents, is located below the waterline, at the base of the catena (Fig. 9) and is characterized by a sodium-rich, deep duplex soil called the sodic zone (Fig. 10) (Venter et al., 2003; Khomo & Rogers, 2005; Grant & Scholes, 2006; Siebert & Eckhardt, 2008). Primary productivity was estimated for each of the three herbivore treatments (Figure 14) using a disc pasture meter (DPM) calibrated in lowveld savanna.
Extension and validation of the Tea Bag Index as a viable approach to quantify decomposition in a semi-arid African savanna
- Preamble
- Introduction
- Materials and Methods
- Results
- Discussion
- Conclusion
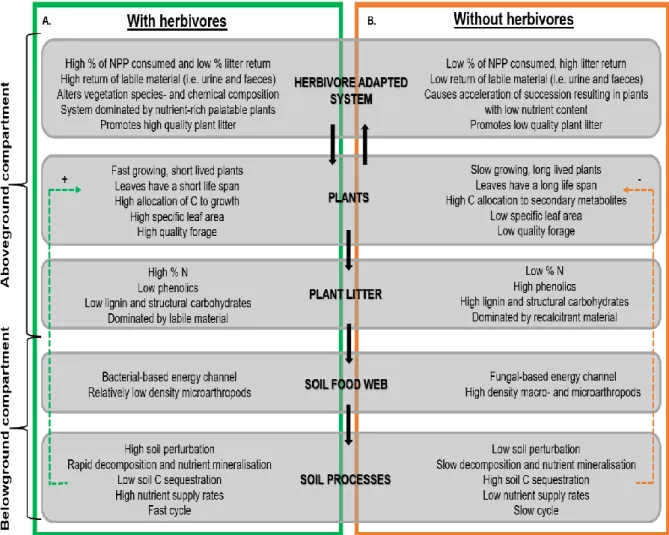
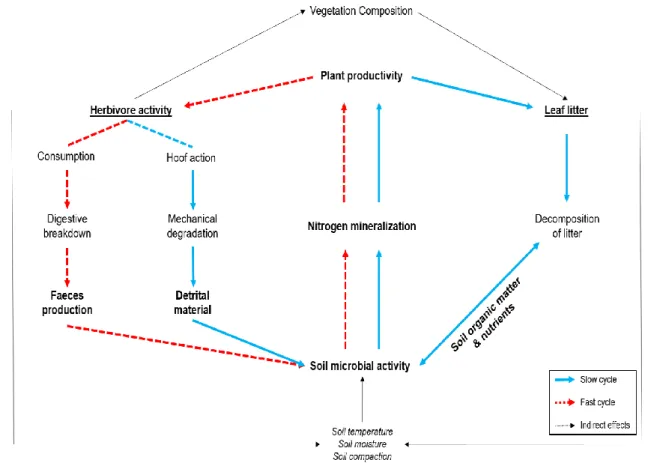
TBI has been developed and used in systems where decomposition is primarily dominated by microbial activity (Keuskamp et al., 2013; Sundsdal, 2019; Teo et al., 2020). Arthropods, including meso- and macrofauna, are an essential component of trophic structure (Coleman et al., 2017). In general, aboveground disturbances, such as those associated with herbivory activity, have a negative effect on arthropod communities (Bardgett et al., 2002).
The magnitude of the disturbances is believed to be greater in densely packed herbivore communities (Vandegehuchte et al., 2017). Fungal communities follow similar biogeographical patterns to plants and are mainly influenced by climatic factors (Vermeire et al., 2021).
Decomposition rates of standardised organic material across herbivore treatments
- Preamble
- Introduction
- Materials and Methods
- Results
- Discussion
- Conclusion
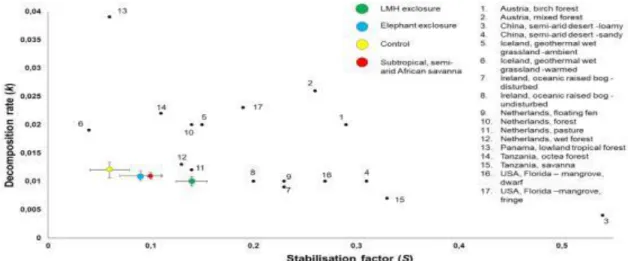
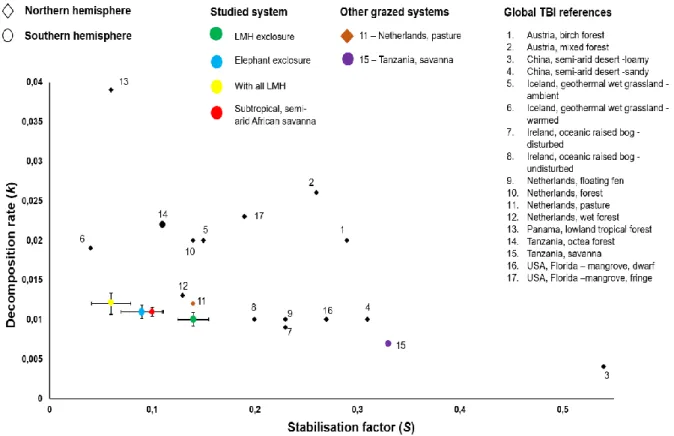
Methodology corresponded to the extended, site-specific version of the TBI approach as described in Teatime in Kruger (Erasmus et al., 2021). All measured k and S values were within the global TBI reference value range derived from Keuskamp et al. This supports the general view that litter stabilization (S) is more dependent on system-specific environmental and climatic factors, reflecting the influence of site-specific drivers (Keuskamp et al., 2013; Becker & Kuzyakov, 2018).
Furthermore, the study area is located in a moisture-limited system (561 mm per year), which further inhibits microbial activity and subsequent decomposition (Grant & Scholes, 2006; Keuskamp et al., 2013; Becker and Kuzyakov, 2018; Petraglia et al. , 2019 ). It is suggested that in productive ecosystems, the beneficial effects of naturally occurring LMH may outweigh the negative effects (Vandegehuchte et al., 2017).
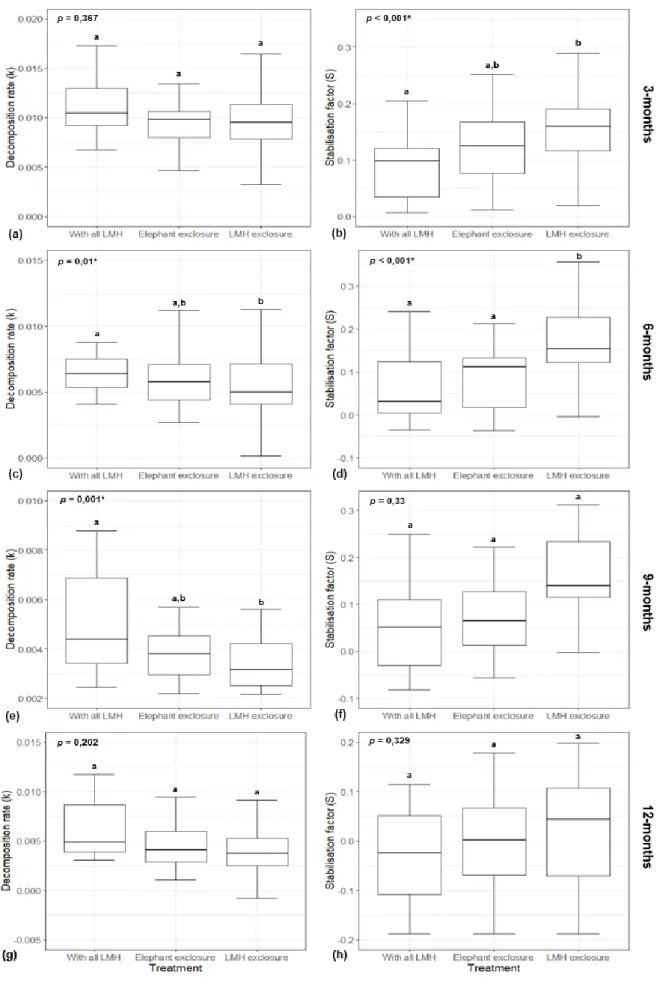
A detailed account of potential decomposition rate and stabilisation factor at several incubation lengths across herbivore treatments
- Preamble
- Introduction
- Materials and Methods
- Conclusion
Existing soil analyzes of the study area were undertaken for two herbivore treatments (i.e. With herbivores and No herbivores) (Van Coller et al., in prep). Data were obtained from Van Coller et al. in prep) and is presented here courtesy of the authors. This changes the chemical composition of root tissue, and by extension underground harmful material and also root exudation (Bardgett et al., 1998).
The degradation is mainly regulated by the chemical composition of the detrital substrate (Keuskamp et al., 2013; Eldridge et al., 2019). Increased SOM results in fertile soil with higher productivity potential (biomass production) as observed with the LMH exclusion (Van Coller et al., 2013; Van Coller & Siebert, 2015).
Soil microbial community structure across various herbivore intensities
- Introduction
- Materials & Methods .1 Soil sampling
- Results
- Conclusion
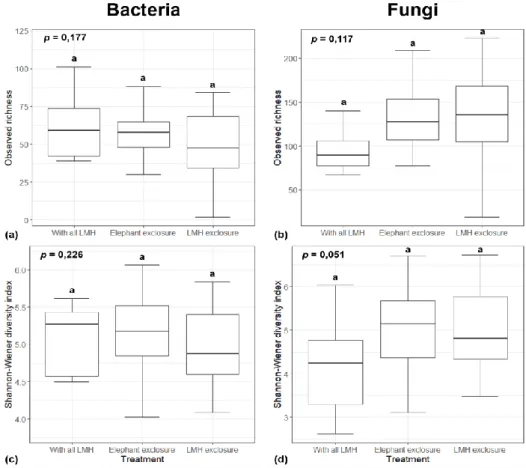
The denoized reads were assigned taxonomy by the Silva 16S rRNA taxonomy (Release 132) ( Quast et al., 2013 ) using a trained classifier of the V3–V4 region. This could potentially be attributed to a lack of ITS sequence updates and/or curation in the UNITE database (López-García et al., 2021). In contrast to these findings, Vermeire et al. 2021) reported considerable variation in bacterial and fungal richness for both ridge and riparian areas located above and below the soot zone.
More herbaceous and woody cover associated with elephant excreta compared to an area with all LMH present (Van Coller et al., 2013) may potentially allow relatively higher soil moisture by limiting evaporation. Similar to the findings of Vermeira et al. 2021), bacterial and fungal community composition showed a high degree of resistance to changes caused by herbivores, especially compared to above-ground changes in vegetation structure.
Linking above- and belowground systems: Linking changes in vegetation structure to belowground processes
Preamble
Introduction
Existing physico-chemical soil data from the study area were used to clarify and support patterns of change in above- and below-ground communities and processes.
Materials and Methods
Species that could not be identified on site were collected and later compared with specimens housed at the AP Goossens herbarium (PUC) at North-West University, Potchefstroom Campus for accurate identifications. Plant trait frequencies were calculated as a percentage of the number of individually counted herbaceous species per treatment as a means of accounting for the potential influence of certain properties on degradation. Furthermore, annuals tend to fall within the acquisitive end of the leaf economy spectrum, which relates to higher leaf N.
Although most of the fixed N is used in the production of seeds, the remaining N within root nodules is released into the soil as the legume dies. To test for significant variation in biomass of the herbaceous layer between three different treatments of different herbivory intensity, data were analyzed using R version 4.1.0 (R Core Team, 2021).
Results
Because biomass data (based on DPM and herbaceous clippings) did not meet the assumption of normality (p < 0.05), a Kruskal–Wallis one-way analysis of variance (ANOVA) was applied to the data to test for significant variation among herbivore treatments. . Exclusion for elephants, elephants (and giraffes) excluded; LMH exclusion, all LMH excluded; p, p < 0.05 indicates significant variance between groups as assessed by the Kruskal-Wallis test. Exclusion for elephants, elephants (and giraffes) excluded; LMH exclusion, all LMH excluded; p, p < 0.05 indicates significant variance between groups assessed by paired t-test.
With all LMH (control), all herbivores are present; Exclusion of elephants, elephants (and giraffes) excepted; LMH exclusion, all LMH excludes. With all LMH (control), all herbivores are present; Exclusion of elephants, elephants (and giraffes) excepted; LMH exclusion, all LMH excluded.
Discussion
High quality root litter, along with improved root secretion, further promotes microbial activity (Bardgett et al., 1998). Studies conducted by Goldfarb et al. 2021) revealed that soil chemical properties are important determinants of bacterial community composition. In addition, bulk density indirectly affects CEC in soils, especially those with a high clay fraction (Xu et al., 2012).
Consequently, fungi have been observed to follow similar biogeographical patterns than those of vegetation (Bui et al., 2020). Existing soil data support this as NO3 (commonly used as a source of N by fungi) (Ngwene et al., 2013), was found to be significantly lower under herbivore exclusion (Figure 5f).
Conclusion
- Main findings
- The way forward
Since the majority of studies have been conducted in the Northern Hemisphere, lower values obtained from the study site can be mainly attributed to the combined effect of global factors such as climate and edaphic properties. In addition, seasonality was found to have a major impact on α-diversity measures, with fungal richness and diversity showing significant variation between dry-season herbivore treatments. Despite the general lack of significant variation between herbivore treatments and across different seasons, the most consistent effect of herbivory appeared to be associated with significant changes in the relative abundance of both bacterial and fungal taxa.
Such information can aid in the development of surface and subsurface conservation strategies specifically aimed at preserving subsurface heterogeneity as a means of managing nutrient cycling. The levels of herbaceous biomass increase in the absence of herbivory, which in turn corresponds to the highest fungal α-diversity and stabilization (S).
Interactive effects of ungulate herbivores, soil fertility and variable rainfall on ecosystem processes in a semiarid savanna. Plant traits explain the distribution of soil microbial communities in two contrasting parts of the world. Teatime in Kruger: tailoring the application of the Tea Bag Index approach to an African savannah.
Effects of grazing and rainfall variability on root and shoot degradation in a semiarid grassland. Long-term effects of fire frequency and season on herbaceous vegetation in savannas of Kruger National Park, South Africa. Responses of the functional structure of the soil microbial community to livestock grazing in the Tibetan alpine grassland.
Teatime in Kruger: Tailoring the application of the Teabag index approach to an African savanna.
Teatime in Kruger: Tailoring the application of the Tea Bag Index approach to an African savanna
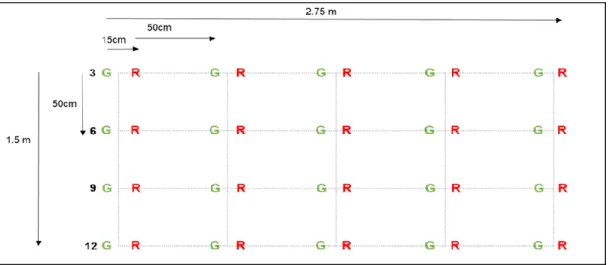
In addition, detritivore activity in the form of termites has a significant impact on the lifespan of the tea bags. Adequate recovery (78%) with small differences between green tea bags and rooibos tea bags will lead to safe conclusions from estimates of TBI. 1000 non-woven polypropylene tea bags (500 green and 500 rooibos) buried in pairs (15 cm apart) Green and rooibos tea bags buried in pairs (15 cm.
Rows contained five pairs of tea bags (five columns) buried 50 cm apart at a depth of 8 cm. Towards harmonization of leaf litter decomposition studies using standard tea bags – a field study and model application.
Decomposition Rates of Organic Material across Herbivore Treatments in a Nutrient-Rich Semi-Arid Sodic Savanna
Herbivory is a disturbance agent and acts as one of the main drivers of system dynamics in African savannas (Grant and Scholes, 2006; Van Coller et al., 2013; Tuomi et al., 2019). Many studies have reported the ability of large mammal herbivores (LMH) to change the vegetation structure and edaphic properties of the system (Grant and Scholes, 2006; Van Coller et al., 2013; Van Coller & Siebert; 2015; Wigley et al. , 2020) . In South African savannas, edaphic factors are stronger drivers of intensively used patches (IUPs) than animal biomass itself (Grant and Scholes, 2006; Van Coller et al., 2013).
Sodic patches are ecologically important as they provide a positive nutrient feedback loop promoted by the cumulative activity of herbivores (Grant and Scholes, 2006; Van Coller et al., 2013). Such losses can affect several fundamental processes including soil-based decomposition and stabilization of detrital plant material (Becker and Kuzyakov, 2018; Tuomi et al., 2019; Wigley et al., 2020).