Co-infection with the human immunodeficiency virus (HIV) and the rise of drug resistance pose significant threats to the control of TB (Raviglione and Smith, 2007; Chindelevitch et al. 2015), especially in sub-Saharan Africa. A small population (2–5%) of treated TB patients go on to develop relapses, further complicating TB epidemics (Weis et al. 1994; Tuberculosis Trials Consortium, 2012). Retreatment cases are often associated with increased costs, which may prove problematic for developing countries with limited national TB budgets (Panjabi et al. 2007).
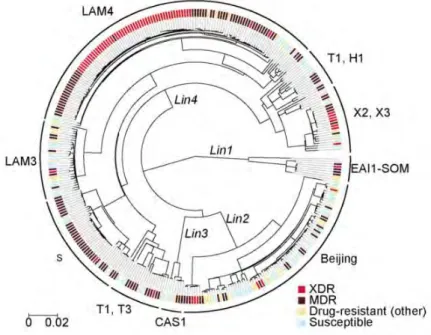
These include spacer oligo typing (Spoligotyping) and mycobacterial interspersed repetitive unit variable number tandem repeat (MIRU-VNTR) typing (Supply et al. 2000; Supply et al. 2001). Gagneux et al. 2006), with Lineages 2 (Beijing) and 4 (Euro-American) being the most geographically widespread and virulent (Coscolla and Gagneux, 2014). The overrepresentation of specific genotype families among cases of drug resistance and transmission is of paramount importance with regard to tuberculosis control regimens (Marais et al. 2006).
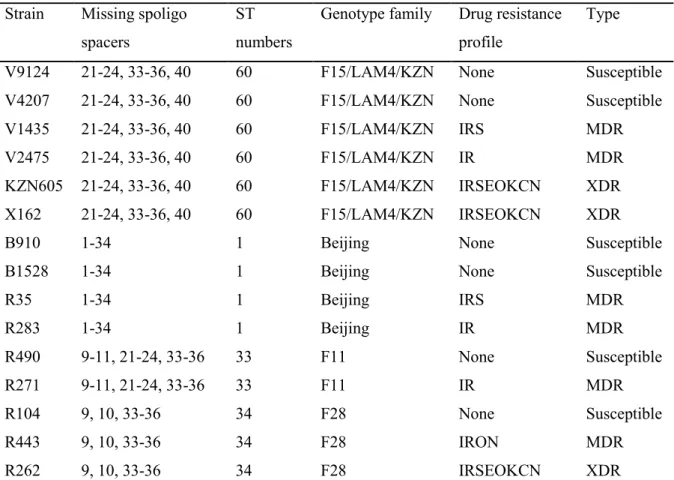
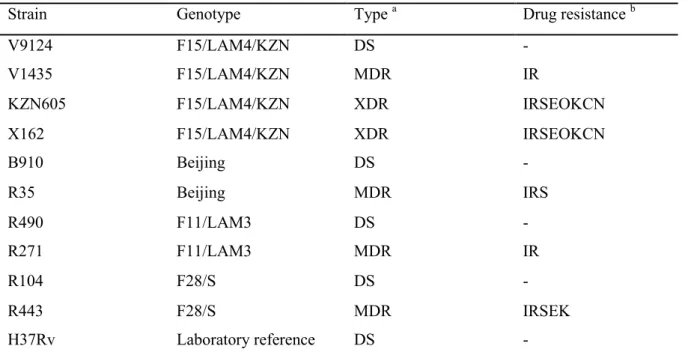
7 The F15/LAM4/KZN genotype was responsible for the Tugela Ferry XDR-TB outbreak, in which 52 of 53 HIV-co-infected patients died within a median time of 16 days (Gandhi et al. 2006) . A C-T nucleotide substitution at codon 491 of the rrs gene not associated with streptomycin resistance is actually a useful phylogenetic marker for the F11 genotype (Victor et al. 2001).
Mechanisms of drug resistance
The wide distribution of Beijing strains (Bifani et al. 2002) can be accredited to the increased virulence in macrophages and animal models (Manca et al. The S315T katG mutation is more frequently detected in clinical isolates and is also associated with a high level of INH resistance (MIC > 1 µg/ml) (Vilcèze et al. 2007; Fenner et al. 2012) Combinations of mutations in the regulatory and coding regions of inhA have recently been shown to induce high-level INH resistance and cross-resistance to ETH (Machado et al. 2013).
Resistance to ethambutol is commonly associated with the embB mutations, particularly those in codon 306 (Telenti et al. 1997; Sreevatsan et al. 1997), but some studies have also reported their occurrence in ethambutol-susceptible isolates (Ahmad et al. 2007). . This is made possible by the pyrazinamidase/nicotinamidase enzyme encoded by the pncA gene (Konno et al. 1967; Scorpio and Zhang, 1996).
By altering the level of 16S rRNA, these drugs act to inhibit protein synthesis (Alangaden et al. 1998). Drug resistance remains a complex process with some resistant strains harboring uncharacterized mutations (Zhang et al. 2013).
Fitness, epistasis and compensatory evolution
Virulence and immunogenicity
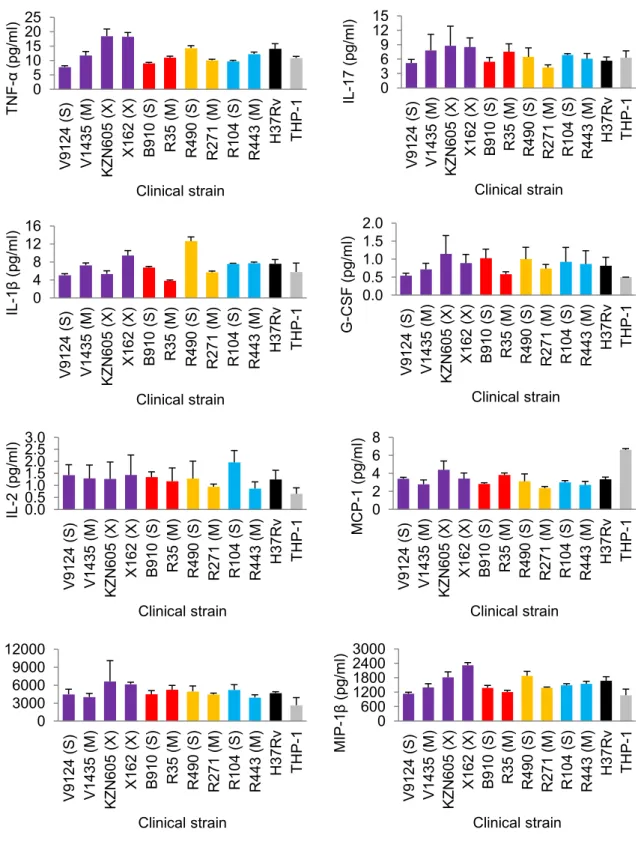
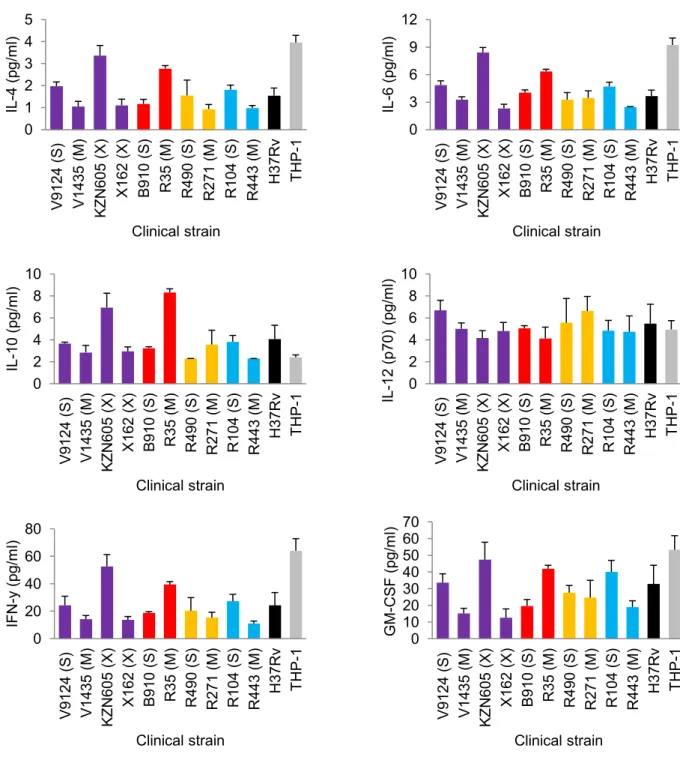
Several studies have used macrophage models to assess intracellular fitness of drug-resistant bacteria. 2004) measured the independent and competitive fitness of 3 spontaneous RIF-resistant mutants in a U937 macrophage cell line, and showed reduced fitness compared to the parental strain. Similarly, Rey-Jurado et al. 2011) assessed the fitness of katG S315T mutant and MDR isolates in J774.2 murine macrophages, and showed reduced fitness relative to susceptible isolates. 15 which emphasizes the importance of infectivity as an indicator of extrapulmonary dissemination (de Viedma et al. 2005).
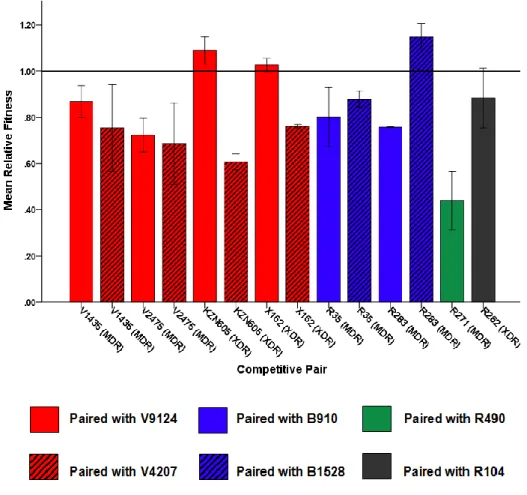
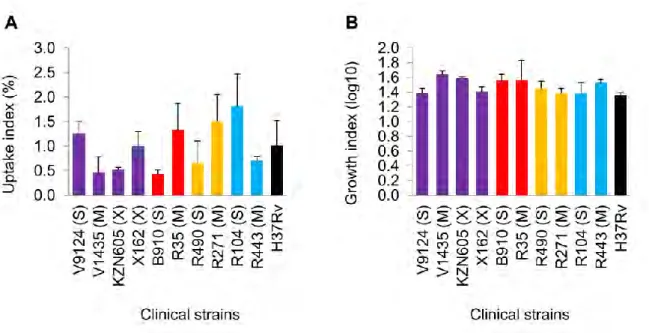
A later study by Barczak et al. 2005) looked at immune responses during dual infection with varying starting ratios of CDC1551 (hyperimmunogenic and hypovirulent) and W-Beijing isolate HN878 (hypoimmunogenic and hypervirulent). Interestingly, both strains had similar growth rates in liquid media, suggesting that the observations under in vivo condition were not due to differences in fitness. Similarly, increased in vitro fitness of drug-resistant F15/LAM4/KZN strains (Naidoo and Pillay, 2014) does not necessitate increased virulence as shown in a recent study (Smith et al. 2014).
Their study showed decreasing virulence in increasingly drug-resistant strains (susceptible > MDR > XDR), but it should be mentioned that all strains were virulent enough to kill SCID mice and showed high levels of necrosis relative to the Erdman strain (Smith et al. 2014). Although it may not be clear how accurately in vitro findings correlate with in vivo conditions, laboratory studies can still reveal important information regarding resistance and fitness-compensating mutations that may be conferred in the natural environment (Bjorkman and Andersson, 2000).
Research aim
Objectives
Scope of thesis
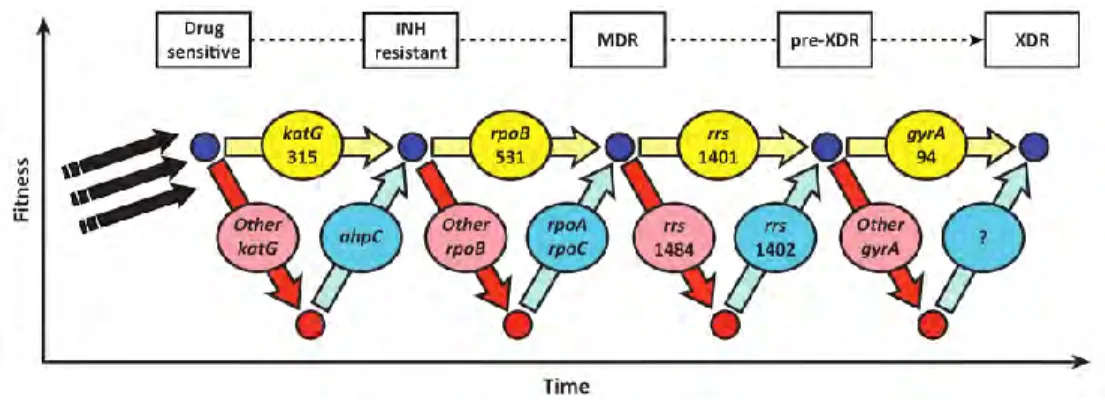
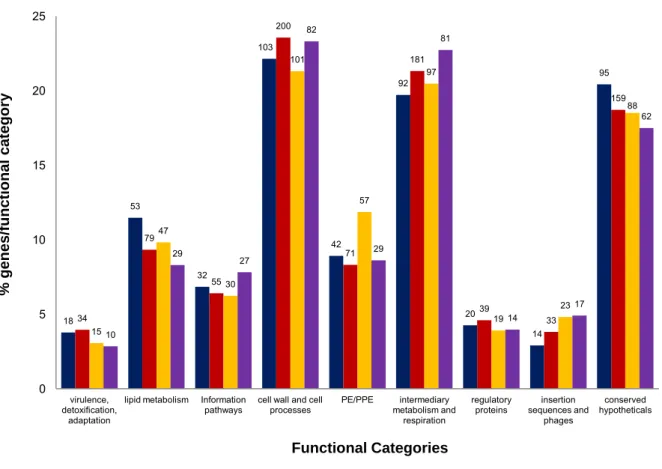
Compensatory mutations, however, can help restore the fitness of drug-resistant strains (Sherman et al. 1996; de Vos et al. 2013). Non-synonymous SNPs (nSNPs) were classified into functional groups according to Tuberculist (Lew et al. 2011). This gene is a member of the dormancy regulon and is strongly up-regulated under hypoxic conditions (Sharpe et al. 2008).
A recently characterized silent mutation, mabAg609a (Ando et al. 2014) was detected in our Beijing MDR strain. Mutations in drrA and ctpB (G23S)—a gene believed to encode a putative copper transporter (Knapp et al. 2015), were uniquely present in drug-resistant KZN strains. 55 Clinical strains containing identical resistance-conferring mutations can demonstrate variable MICs (Kim et al. 2003).
Similarly, the fitness of rifampicin-resistant strains harboring rpoB mutations can be restored by rpoC compensatory mutations (de Vos et al. 2013). Drug-resistant strains sometimes undergo fitness costs, depending on the resistance-encoding mutations they harbor and their genetic background (Gagneux et al. 2006). Genomic DNA was isolated by the cetyltrimethylammonium bromide (CTAB) method with some modifications (van Soolingen et al. 2001).
Spoligotyping was performed as previously described (Kamerbeek et al. 1997) using a commercially available kit (Ocimum Biosolutions, Hyderbad, India). Minimal inhibitory concentrations (MICs) were determined in a 96-well plate format using Alamar Blue (Invitrogen) as previously described ( Franzblau et al. 1998 ).