Where the work of others was used, this is acknowledged in the text. This study investigates the effects of forest fragmentation and matrix type on the diversity and distribution of epigaean invertebrates in selected Afromontane forests of midland KwaZulu-Natal. This range of species in the mist belt mixed Podocarpus forests of midland KwaZulu-Natal will provide an accurate indication of forest condition in summer when abundance data are used.
It also contains 14.1% and 14.2% of the South African mammal and bird species respectively (Geldenhuys and MacDevette, 1989).
Sampling design and non-analytical methods
Results
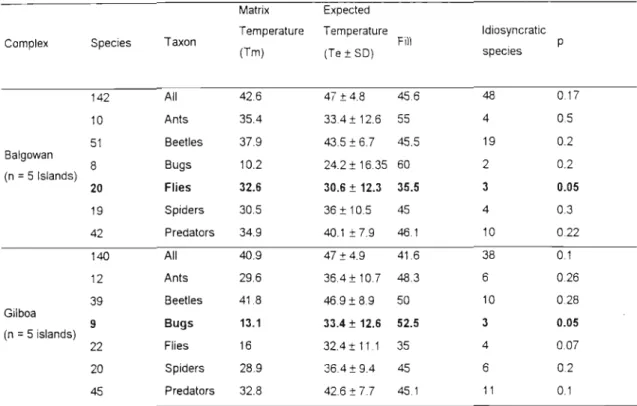
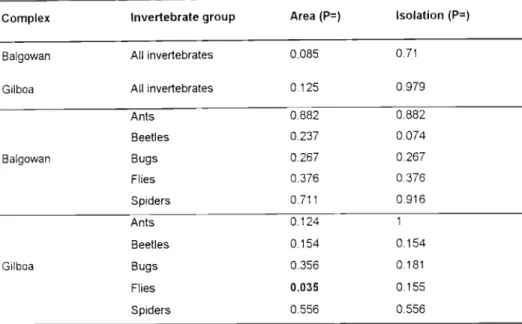
The Lomolino model showed no significance of nestedness when sites were ranked by area or isolation, with the exception of the Gilboa fly community which was significant (P < 0.05) when sites were ranked by area (Table 4). These results are counterintuitive, as some degree of nestedness appears to be the rule in nature (Wright etI found that of the 279 archipelagos analyzed, there were a few archipelagos in which the communities studied were significantly non-nested. Idiosyncratic species could variously recolonize some islands from which they have been locally extirpated, reach their range limits in the middle of the archipelago, or become victims of competitive exclusion or products of local speciation (Patterson and Atmar, 2000).
The anti-breeding nature of epigean invertebrates in this study may have been due to invasion of Podocarpus mixed forest fragments in the fog belt from matrices by species from the genus list, with greater numbers invading from the grassland matrix as a natural result of the higher number of invertebrate species found here. The variation around the boxed matrix extinction threshold limit produced by the temperature calculation program (cf. Atmar and Patterson, 1993) for epigean invertebrates in this study was high (fewer nested) compared to truly isolated species such as terrestrial isopods and terrestrial snail communities in the Cyclades Islands, Greece (Sfenthourakis et al., 1999), which were highly nested and driven to extinction. While it is easy to obtain a measure of island area, any measure of isolation must be biologically meaningful at the level of the organisms being studied, which is very difficult to do (Lomolino, 1996), especially with such a diverse group (with different responses to external factors) as a community of epigeic invertebrates.
If an extinction-driven community structure of epigaeic invertebrates is observed and due to the similarity of the communities throughout all forests, and the lack of nests, I expect that convergence of epigaeic invertebrate communities has already occurred in the smaller fragments, with patterns of convergence is clouded due to subsequent and continuous invasions from the matrix. In the next chapter, I investigate how individual species respond to the fragmentation and matrix transformation of the Karkloof/Balgowan/Gilboa forests, and attempt to identify ecological indicators of forest condition from the epigaeic invertebrate community. Settlement and dispersal of colony-founding queens of the tree ant, Crematogaster ashmeadi, in a longleaf pine forest.
The distribution of the samango monkey (Cercopithecus mitis erythrarchus Peters, 1852 and Cercopithecus mitis labiatus I. Geoffroy, 1843) and forest history in southern Africa. Eyeless arthropods from the Galapagos Islands, Ecuador: composition and origin of the cryptozoan fauna of a young, tropical, oceanic archipelago.
The diversity and species-area relationships of epigaeic invertebrate assemblages in isolated mistbelt mixed Podocarpus forest fragments
Introduction
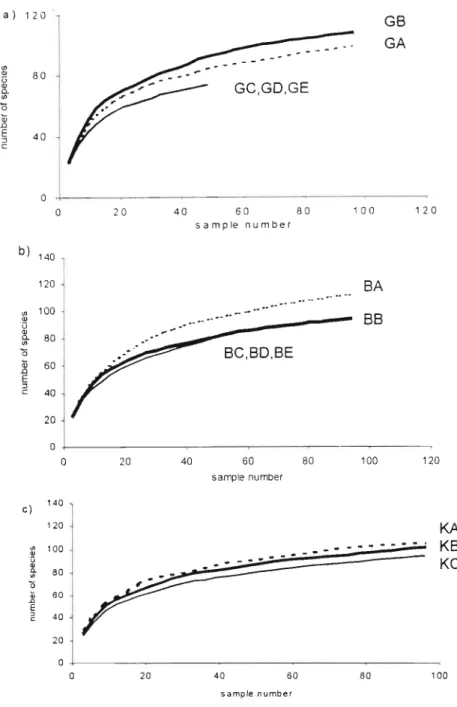
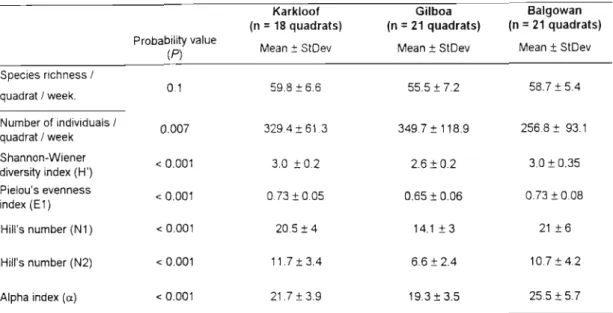
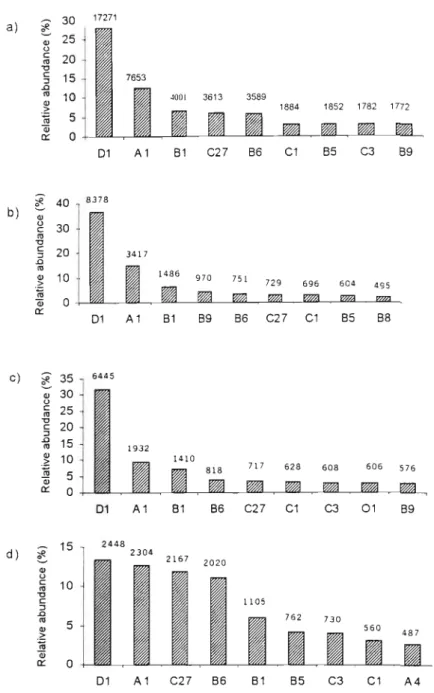
Density compensation usually describes an inverse relationship between population density and species richness (McGrady-Steed and Morin, 2000; Stevens and Willig, 2000). Epigaeic invertebrate point diversity (a. diversity, or species richness per quadrat), and differentiation diversity (~ diversity, or species turnover between quadrats and forests) were calculated (Magurran, 1988). In the large (control) Karkloof Forest, the abundance of the four dominant species was more evenly distributed than in the two forest complexes (Fig 1).
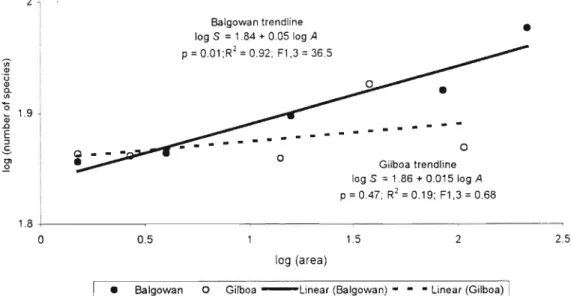
Log - tree species richness (estimates of rarity, each forest standardized to 2311 individuals) regressed against forest area (recorded value). It is therefore a possibility that area did not influence the richness of rare epigaeic invertebrate species in the Gilboa forests while it did in the Balgowan forests. The average species richness per quadrat as a percentage of the total species richness obtained from a forest was 40% for Gilboa and 41% for Balgowan.
These findings suggest that pine matrix influenced epigaean invertebrate species richness in fog belt-mixed Podocarpus forest fragments. Regression results by species area for selected epigaean invertebrate taxa, illustrating the lack of an area effect on species richness for the majority of taxa. Log-log species-area regression for predatory species, illustrating the lack of area effect on species richness.
In contrast, epigaean invertebrate densities of Gilboa showed a non-significant, positive relationship with species richness. Habitat heterogeneity does not explain the variation in species richness among the forests observed in this study.
Discussion
However, the species richness of epigaean invertebrates of the Balgowan Complex may have been area dependent, and the species richness-dependent density compensation in the Balgowan Complex would then suggest that the epigaean invertebrate assemblages were approaching saturation if they were not already saturated with species . The overall positive correlation between population density and species richness in the Gilboa could arise from a number of mechanistic processes acting individually or in concert, and the mechanisms may differ between species (Connor et al. 2000). In the Karkloof, a less disturbed fog belt mixed Podocarpus forest habitat, dominance was shared with two other species.
Within four weeks of the wildfire, there was a marked increase in the abundance and dominance of the seed-feeding ant Prolasius pallidus. The matrix has three potential roles in fragment community formation (Davies et al., 2001). In a study by Armstrong et al. 1996b), the number of insect-pollinated plant species dropped from 212 in natural fynbos to 82 species in pine plantations alone.
Forests within the grassland matrix showed a significant species-area relationship, while no such relationship was detected for forests within the pine matrix. Animal-habitat relationships in the Knysna Forest, South Africa: distinction between forest types by birds and invertebrates. The impact of forest clearing on the diversity of ground beetles (Coleopters: Carabidae) in the southern Appalachians.
Ed.), Development of International Principles and Practices of Wildlife Research and Management: Asian and American Approaches. In: Macdonald, lAW., Kruger, F.J., and Ferrar, AA (eds.), Ecology and Management of Biological Invasions in Southern Africa.
34;shopping-basket" of ecological indicators
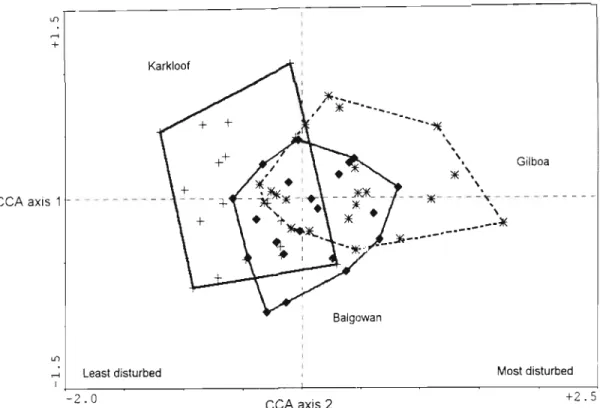
I expected those fragments of mixed Podocarpus forest with mist belt in pine plantation matrix (Gilboa fragments) to be more disturbed than those surrounded by grassland (Balgowan fragments) and Karkloof forest to be less disturbed in terms of measured habitat variables. It was expected that invertebrate diversity would fluctuate according to individual species-response to the calculated disturbance, and thus that a group of species would serve as environmental indicators of forest condition. Thus, the distribution of sites along the CA axes is at a maximum (Ter Braak, 1995).
This multivariate technique selects a linear combination of environmental variables that maximize the dispersion of species scores along the first CCA axis (Jongman et al., 1987; Ter Braak and Prentice, 1988; Palmer, 1992). Therefore, by comparing the eigenvalues of the ordination axes under unconstrained conditions (CA) with those constrained by measured environmental conditions (CCA), we can assess whether the most important environmental variables were included in the analysis as predictors of species abundance (Okland, 1996). ). In this study, quadrats were used as the basic unit of analysis, as the response of invertebrates to environmental disturbance was likely to be at the quadrat rather than forest scale.
However, there are many criticisms of the use of morphospecies, including the fact that the life histories of individual morphospecies and thus their ecological significance are unknown (King et al., 1998; Hamer and Slotow, 2000). The family is represented in all forests and quadrats and comprises four of the seven morphospecies in the "shopping basket" of the above indicators. Talistroides africana (Amphipoda) show a significant, positive response to increasing disturbance, and the slope of the abundance/disturbance regression was very steep relative to other species, indicating a high sensitivity and rate of increase in response to disturbance.
Ants as bioindicators of habitat disturbance: validation of the functional group modal for the humid tropics of Australia. Arthropods as indicators of the effects of fluoride pollution on post-sand mining succession.
Summary and conservation and management recommendations
Density offset (decrease in species density with increasing species richness in a patch) in the Balgowan Complex suggests that epigeic invertebrate assemblages are approaching saturation, if not already species saturated. This lack of density compensation in Gilboa can be explained by the "enemies hypothesis" or the "movement hypothesis". According to the "movement hypotheses", the pine matrix acts to further isolate the indigenous forest fragments in Gilboa.
Recolonization dynamics may therefore play a small role in the regional persistence of a species within a pine matrix, and persistence will be ensured by the preservation of a small number of large habitat fragments containing large, extinction-resistant populations. This study did not sample epigaeal invertebrates in the grassland and pine matrices to identify those species that invaded the fog belt forests, but this is a future research opportunity in this area. Talistroides africana and Tetramorium avium dominated epigaeic invertebrate abundance in both Gilboa and Balgowan, although to a lesser extent in the grassland-surrounded Balgowan complex.
In Karkloof, in the least disturbed forest, dominance was shared with two other species. A habitat disturbance gradient was evident from the relatively undisturbed Karkloof quadrats to the more disturbed Gilboa quadrats situated in the pine matrix, with intermediate disturbance values for the Balgowan fragments. Leaf litter depth was the most important variable in explaining the variation of epigeic invertebrate species.
Epigaean invertebrates appeared to be a sensible choice for ecological indicators at a general level, as species-area relationships and trends in density compensation mirrored those found in birds by Wethered (2001) in the same region. This range of species (abundance data) in the mist belt mixed Podocarpus forests of midland KwaZulu-Natal will provide an accurate indication of forest condition in summer.