The experimental work described in this thesis was carried out by the candidate while based in the Biological Sciences Discipline, School of Life Sciences, College of Agriculture, Engineering and Science, University of KwaZulu-Natal, Pietermaritzburg Campus, under the supervision of Professor Richard P .The content of this work has not been submitted in any form to another university and, unless the work of others is known in the text, the results reported are due to investigations by the candidate.
Plagiarism
Publication
The present study was undertaken to characterize DT in both ferns, particularly the roles of reactive oxygen species (ROS) scavenging enzymes superoxide dismutase (SOD) and peroxidase (POX), sugar accumulation and abscisic acid. Long-term rapid desiccation reduced POX activity in both species, but activity recovered upon rehydration. 16 Table 3: Ion leakage of Crepidomanes inopinatum and Loxogramme abyssinica after one week of silica gel drying (0% RH).
30 Figure 7: The effect of hydrogen peroxide (H2O2) (A) and substrate concentration (o-.dianisidine) (B) on class III peroxidase activity in C.
Literature review
- The pteridophytes
- Occurrence of DT in the pteridophytes
- Morphological and anatomical mechanisms of DT in the pteridophytes
- Physiological and biochemical mechanisms of DT in the pteridophytes
- DT mechanisms based on avoiding ROS formation
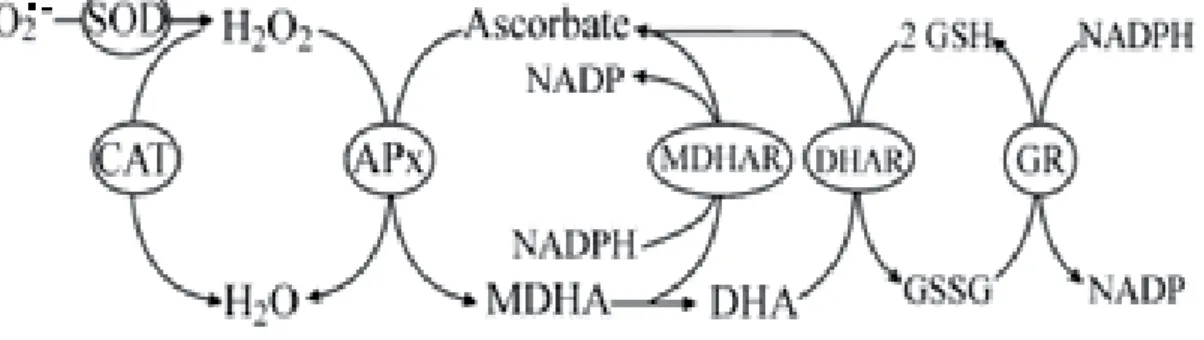
- DT mechanisms based on antioxidant enzymes
- DT mechanisms based on non-enzymatic antioxidants
- DT mechanisms based on sugars
- DT mechanisms based on LEA and other proteins
- Role of ABA in DT
- DT in pteridophytes – summary
- Introduction to present study
Because of such inducible responses to desiccation, Hilker et al. 2016) termed this strategy a “fundamental cost-cutting strategy”. Here, chlorophyll degradation occurs during desiccation, and chlorophylls are resynthesized by rehydration (Bravo et al., 2016; Oliveira and Moraes, 2015) e.g. Their water-conducting cells are arranged to resemble the densely packed tracheid of conifers (Pittermann et al., 2013).
The genomes of most plants generally contain a large number of peroxidase isoforms (Dunand et al., 2003). Third, they stabilize the thylakoid membrane and associated photosystems (Ahmad et al., 2009; Gill and Tuteja, 2010). They may contribute to overall antioxidant protection (Rice-Evans et al., 1996), but also play other important roles.
The accumulation of phenolics in ferns is said to be an important adaptation to the terrestrial environment (Ribeiro et al., 2007). PPO (polyphenol oxidase, a Cu-containing enzyme) together with peroxidase is responsible for the oxidation of phenols by oxygen-dependent oxidation of monophenols or ο-diphenols to o-quinones (Živković et al., 2010). Phenolic acids can also act as a signaling molecule and are mainly stored in the vacuole or in the apoplast (Živković et al., 2010).
In addition to stabilizing dry cells and intracellular glass formation, sucrose can also stabilize many enzymes in the dry state (Toldi et al., 2009). The accumulation of sugars, specifically sucrose, tends to occur at a late stage of desiccation (Toldi et al., 2009). Furthermore, LEA proteins maintain hydrogen bonding within and between a variety of macromolecules (Voytena et al., 2014).
A desiccation-tolerant fern Polypodium polypodioides was found to express 31-kDa putative dehydrin polypeptide during desiccation but not upon rehydration (Layton et al., 2010).
Material and Methods
- Plant material
- Effect of pH on peroxidase activity
- The effect of pH on metabolism of o-dianisidine
- The effect of pH on metabolism of ascorbic acid
- Optimal pH for metabolism of ABTS
- Optimal pH for metabolism of DMP
- Peroxidase characterization
- The effect of substrate concentration on peroxidase activity
- Effect of hydrogen peroxide concentration on peroxidase activity
- Light response curves
- Effect of slow and rapid desiccation on relative water content
- Chlorophyll fluorescence during drying and wetting cycles
- Ion leakage / conductivity
- Effect of slow and fast desiccation and rehydration on enzyme activity
- Superoxide dismutase assay
- Peroxidase activity
- Gel electrophoresis
- Effect of desiccation and rehydration on soluble sugars
- ABA Treatment
- Data analysis
To measure the ascorbic acid (Class I) peroxidases, 50 μL samples were incubated in quartz cuvettes containing citrate-phosphate buffer over a range of pHs and 1 mM EDTA, 5 mM ascorbate, and 1 mM H2O2. To measure the oxidation of ABTS, 100 μl samples were incubated in quartz cuvettes containing citrate-phosphate buffer with a range of different pHs and 10 mM ABTS for 15 minutes at 30˚C. To measure the oxidation of 2,6-dimethoxyphenol (DMP), 25 μl samples were incubated in quartz cuvettes containing 1 mM DMP, Mcllvaine buffer with a range of pH's, made of 0.1 M citric acid and 0.2 M Na2HPO4 and 1mM H2O2.
Enzyme extracts of 25 μL were incubated with a range of concentrations of o-dianisidine with 1 mM H2O2 in sodium acetate buffer pH 5. Actinic light was then turned on and fluorescence parameters were measured in 15 replicates of each species in a range of light intensities. To compare the effect of slow and fast dehydration, Petri dishes with leaves were placed for 48 hours on a saturated solution of calcium acetate ((CH3.COO) 2Ca), giving a relative humidity of 20% and 1 week in a dryer suspended above. silica gel, giving a relative humidity of 0%, respectively at 10°C under dim light.
Ion leakage is a measure of the effect of desiccation stress resulting in ion leakage after rehydration. This was repeated for leaves of both species placed in a desiccator suspended over silica gel for 1 week (168 h) and 56 h of rehydration (as described in section 2.6) and another group rehydrated for 1 week (168 h) . Replicates were homogenized using 5 ml of 50 mM phosphate buffer (pH 7), and then centrifuged at 10,000 g for 5 min.
The effect of ABA on DT was measured using 100 μM ABA (cis, trans; Sigma) which was dissolved in 0.1% DMSO. The effect of ABA treatment was assessed by measuring chlorophyll fluorescence parameters as described in section 2.4.
Results
- The effect of substrate pH on peroxidase activity
- The effect of substrate and hydrogen peroxide concentration on enzyme activity
- Effect of freezing storage
- Light response curves
- Effects of desiccation
- RWC
- Deacclimation effect of storage
- Effect of desiccation on chlorophyll fluorescence
- Effect of desiccation on Ion leakage / conductivity
- Effect of desiccation and rehydration on enzyme activity
- Superoxide dismutase
- Peroxidase activity
- PAGE analysis of peroxidase isoforms
- Effect of desiccation and rehydration on soluble sugars
- Effect of ABA treatment on the tolerance of photosynthesis to desiccation
At hydrogen peroxide concentrations above 5 mM, peroxidase activity begins to gradually decrease in both species. Interestingly, both species rehydrated to the same extent, suggesting that in the dry state the cuticle of L. FV/FM in freshly collected material (open circles) was reduced more than in stored leaves after storage for 1 week on silica gel.
However, during rehydration, while FV/FM in freshly collected leaves recovered to the original values, the stored leaves could not fully recover (open diamonds). Dehydration of both species for one week over silica gel, followed by sudden rehydration, caused very little ion leakage in both species (Table 3). In both species, slow dehydration for 48 hours over calcium acetate has little effect on POX activity (Figure 14A,B).
Activity in both species increased gradually after rehydration and required at least 1 week for full tolerance/recovery (Figure 14E, F). Rapid drying increased the concentration of sugars in both species, but concentrations quickly returned to pre-stress values after rehydration in both species (Figure 16C,D). FV/FM and ETR, but not NPQ, recovered rapidly during rehydration but then gradually declined (Figure 17B, D, F).
The values of FV/FM and ETR were significantly lower in ABA-treated leaves during rehydration. NPQ decreased similarly to FV/FM and ETR during dehydration, but was only recovered to values c.
Discussion
- Effect of freezing storage
- Light response curves
- Deacclimation of freshly collected material
- Both species of ferns are poikilohydric
- Change in enzyme activity
- Superoxide dismutase
- Peroxidase activity
- Gel electrophoresis analysis of peroxidase isoforms
- Change in soluble sugars
- Effect of ABA treatment on the tolerance of photosynthesis to desiccation
The POX activities of both species were maintained at more or less the same levels in dried and intact leaves after drying and slow rehydration (Figure 14A, B). In higher plants, class III peroxidases are often involved not only in ROS scavenging but also in ROS production, which in various ways form an important component of stress tolerance mechanisms (Almagro et al. 2009) . In the pteridophytes studied here, production apparently occurs by enzymes such as NADPH oxidases, which are responsible for ROS production in some higher plants (Marino et al., 2012).
Soluble sugars have been widely reported to protect plants against various abiotic stresses, and are a very important component of DT in the so-called “modified” DT plant (Oliver et al., 1998). As discussed earlier, it seems likely that the “sugars may act to stabilize membranes and proteins in the dry state by maintaining hydrogen bonding within and between macromolecules” (Denev et al., 2014). In the filmy fern Hymenophyllum caudiculatum, ABA treatment can increase DT by apparently stabilizing PSII (Garces et al., 2018).
There are many examples where stress tolerance is induced in plants without the involvement of ABA, and it seems likely that these "ABA independent" pathways operate in the two species of ferns studied here (Yoshida et al., 2014). Reversible in vivo cellular changes occur during desiccation and recovery: Desiccation tolerance of the resurrection filmy fern Hymenophyllum dentatum Cav. Glass formation in plant anydrobiotes: survival in the dry state. Abscisic acid and the induction of desiccation tolerance in the extremely xerophilic liverwort Exormotheca holstii.
Constitutive components and induced gene expression are involved in the dehydration tolerance of Selaginella tamariscina. The role of active oxygen in the response of plants to water shortage and dehydration.
General Conclusion and Recommendations
General conclusion
DT in plants is undoubtedly based on more than one underlying mechanism, conferred by a number of genes, although the precise nature of the ``desiccome'' has yet to be revealed (Kranner et al., 2008). At the cellular level, various biochemical mechanisms have been proposed to contribute to an organism's ability to survive desiccation. Two of the most important of these are efficient systems that reduce stress-induced ROS formation and the synthesis or accumulation of high concentrations of sugars (Kranner and Birtic, 2005).
Our original hypothesis was that constitutive mechanisms would predominate in the filmy fern, while inducible mechanisms would be found in the species with a cuticle. This is supported by the lack of an effect of ABA on DT in this species, presumably because desiccation was too rapid to allow the development of tolerance mechanisms. In general, both species are considered DT as they are able to recover from extreme water loss with minimal damage depending on the rate of dehydration.
Stark (2017) argued that it is important to distinguish between “recovery from stress in the field” and “hardening” and suggested that it is essential to completely harden mosses when they are collected before studying DT. Clearly, the "stress history" of field-collected plants will usually be unknown, but may affect physiological experiments. The effect of deacclimation is very important, as some mosses such as Crossidium can take up to 7 days to deacclimatize, while Sphagnum can take up to 3 weeks (Stark, 2017).
Future recommendation
Reactive oxygen species and antioxidant enzymes involved in plant stress tolerance, in: Shanker, A. Ed.), Abiotic and biotic stress in plants - Recent advances and future perspectives. Desiccation tolerance of Hymenophylaceae film ferns is mediated by constitutive and non-inductive cellular mechanisms. Changes in chlorophyll fluorescence, photosynthetic CO2 assimilation and xanthophyll cycle interconversions during dehydration in desiccation tolerant and intolerant liverworts.
The early history of vascular land plants based on the late Silurian and early Devonian floras of the British Isles. To desiccation and beyond” – desiccation preparation and rehydration in desiccation-tolerant vegetative plants. Comparative ecophysiological measurements on light responses, water relations and desiccation tolerance of the film fern Hymenophyllum willsonii Hook.
Abscisic acid increases the ability of the desiccation-tolerant fern Polypodium virginianum to withstand desiccation. Pleopeltis pleopeltifolia (Polypodiopsida, Polypodiaceae), a desiccation-tolerant poikilochlorophyll fern: anatomical, biochemical and physiological responses during water stress. Mechanism of desiccation tolerance in the resurrection fern Selaginella tamariscina revealed by physiological and proteomic analysis.